
47 results
18 - Chemotaxis and Detection
- from Part II - Single Bacteria
-
- Book:
- The Physics of Bacteria
- Published online:
- 12 December 2024
- Print publication:
- 19 December 2024, pp 190-202
-
- Chapter
- Export citation
3 - Molecular Communication in Biological Systems
-
- Book:
- Molecular Communication
- Published online:
- 12 December 2024
- Print publication:
- 19 December 2024, pp 37-51
-
- Chapter
- Export citation
Dynamical behaviour of a logarithmically sensitive chemotaxis model under time-dependent boundary conditions
- Part of
-
- Journal:
- European Journal of Applied Mathematics , First View
- Published online by Cambridge University Press:
- 04 November 2024, pp. 1-27
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
-
This article studies the dynamical behaviour of classical solutions of a hyperbolic system of balance laws, derived from a chemotaxis model with logarithmic sensitivity, with time-dependent boundary conditions. It is shown that under suitable assumptions on the boundary data, solutions starting in the
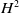 $H^2$-space exist globally in time and the differences between the solutions and their corresponding boundary data converge to zero as time goes to infinity. There is no smallness restriction on the magnitude of the initial perturbations. Moreover, numerical simulations show that the assumptions on the boundary data are necessary for the above-mentioned results to hold true. In addition, numerical results indicate that the solutions converge asymptotically to time-periodic states if the boundary data are time-periodic.
$H^2$-space exist globally in time and the differences between the solutions and their corresponding boundary data converge to zero as time goes to infinity. There is no smallness restriction on the magnitude of the initial perturbations. Moreover, numerical simulations show that the assumptions on the boundary data are necessary for the above-mentioned results to hold true. In addition, numerical results indicate that the solutions converge asymptotically to time-periodic states if the boundary data are time-periodic.
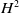
Stabilization in a chemotaxis system modelling T-cell dynamics with simultaneous production and consumption of signals
- Part of
-
- Journal:
- European Journal of Applied Mathematics , First View
- Published online by Cambridge University Press:
- 18 September 2024, pp. 1-14
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
-
In a smoothly bounded domain
 $\Omega \subset \mathbb{R}^n$,
$\Omega \subset \mathbb{R}^n$,  $n\ge 1$, this manuscript considers the homogeneous Neumann boundary problem for the chemotaxis systemwith parameter
$n\ge 1$, this manuscript considers the homogeneous Neumann boundary problem for the chemotaxis systemwith parameter \begin{eqnarray*} \left \{ \begin{array}{l} u_t = \Delta u - \nabla \cdot (u\nabla v), \\[5pt] v_t = \Delta v + u - \alpha uv, \end{array} \right . \end{eqnarray*}
\begin{eqnarray*} \left \{ \begin{array}{l} u_t = \Delta u - \nabla \cdot (u\nabla v), \\[5pt] v_t = \Delta v + u - \alpha uv, \end{array} \right . \end{eqnarray*} $\alpha \gt 0$ and with coincident production and uptake of attractants, as recently emphasized by Dallaston et al. as relevant for the understanding of T-cell dynamics.
$\alpha \gt 0$ and with coincident production and uptake of attractants, as recently emphasized by Dallaston et al. as relevant for the understanding of T-cell dynamics.It is shown that there exists
 $\delta _\star =\delta _\star (n)\gt 0$ such that for any given
$\delta _\star =\delta _\star (n)\gt 0$ such that for any given  $\alpha \ge \frac{1}{\delta _\star }$ and for any suitably regular initial data satisfying
$\alpha \ge \frac{1}{\delta _\star }$ and for any suitably regular initial data satisfying  $v(\cdot, 0)\le \delta _\star$, this problem admits a unique classical solution that stabilizes to the constant equilibrium
$v(\cdot, 0)\le \delta _\star$, this problem admits a unique classical solution that stabilizes to the constant equilibrium  $(\frac{1}{|\Omega |}\int _\Omega u(\cdot, 0), \, \frac{1}{\alpha })$ in the large time limit.
$(\frac{1}{|\Omega |}\int _\Omega u(\cdot, 0), \, \frac{1}{\alpha })$ in the large time limit.








Global boundedness and large time behaviour in a higher-dimensional quasilinear chemotaxis system with consumption of chemoattractant
- Part of
-
- Journal:
- Proceedings of the Royal Society of Edinburgh. Section A: Mathematics , First View
- Published online by Cambridge University Press:
- 30 April 2024, pp. 1-26
-
- Article
-
- You have access
- HTML
- Export citation
-
This paper deals with the following quasilinear chemotaxis system with consumption of chemoattractant
\[ \left\{\begin{array}{@{}ll} u_t=\Delta u^{m}-\nabla\cdot(u\nabla v),\quad & x\in \Omega,\quad t>0,\\ v_t=\Delta v-uv,\quad & x\in \Omega,\quad t>0\\ \end{array}\right. \]in a bounded domain $\Omega \subset \mathbb {R}^N(N=3,\,4,\,5)$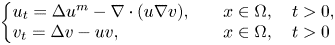
 with smooth boundary $\partial \Omega$
with smooth boundary $\partial \Omega$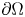 . It is shown that if $m>\max \{1,\,\frac {3N-2}{2N+2}\}$
. It is shown that if $m>\max \{1,\,\frac {3N-2}{2N+2}\}$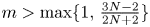 , for any reasonably smooth nonnegative initial data, the corresponding no-flux type initial-boundary value problem possesses a globally bounded weak solution. Furthermore, we prove that the solution converges to the spatially homogeneous equilibrium $(\bar {u}_0,\,0)$
, for any reasonably smooth nonnegative initial data, the corresponding no-flux type initial-boundary value problem possesses a globally bounded weak solution. Furthermore, we prove that the solution converges to the spatially homogeneous equilibrium $(\bar {u}_0,\,0)$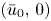 in an appropriate sense as $t\rightarrow \infty$
in an appropriate sense as $t\rightarrow \infty$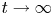 , where $\bar {u}_0=\frac {1}{|\Omega |}\int _\Omega u_0$
, where $\bar {u}_0=\frac {1}{|\Omega |}\int _\Omega u_0$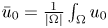 . This result not only partly extends the previous global boundedness result in Fan and Jin (J. Math. Phys. 58 (2017), 011503) and Wang and Xiang (Z. Angew. Math. Phys. 66 (2015), 3159–3179) to $m>\frac {3N-2}{2N}$
. This result not only partly extends the previous global boundedness result in Fan and Jin (J. Math. Phys. 58 (2017), 011503) and Wang and Xiang (Z. Angew. Math. Phys. 66 (2015), 3159–3179) to $m>\frac {3N-2}{2N}$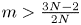 in the case $N\geq 3$
in the case $N\geq 3$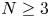 , but also partly improves the global existence result in Zheng and Wang (Discrete Contin. Dyn. Syst. Ser. B 22 (2017), 669–686) to $m>\frac {3N}{2N+2}$
, but also partly improves the global existence result in Zheng and Wang (Discrete Contin. Dyn. Syst. Ser. B 22 (2017), 669–686) to $m>\frac {3N}{2N+2}$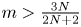 when $N\geq 2$
when $N\geq 2$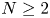 .
.
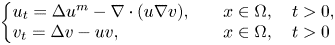
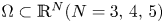
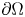
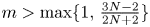
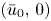
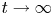
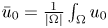
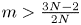
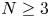
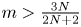
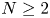
The fully parabolic multi-species chemotaxis system in
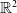 $\mathbb{R}^{2}$
$\mathbb{R}^{2}$
- Part of
-
- Journal:
- European Journal of Applied Mathematics / Volume 35 / Issue 5 / October 2024
- Published online by Cambridge University Press:
- 19 January 2024, pp. 675-706
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
-
This article is devoted to the analysis of the parabolic–parabolic chemotaxis system with multi-components over
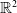 $\mathbb{R}^2$. The optimal small initial condition on the global existence of solutions for multi-species chemotaxis model in the fully parabolic situation had not been attained as far as the author knows. In this paper, we prove that under the sub-critical mass condition, any solutions to conflict-free system exist globally. Moreover, the global existence of solutions to system with strong self-repelling effect has been discussed even for large initial data. The proof is based on the modified free energy functional and the Moser–Trudinger inequality for system.
$\mathbb{R}^2$. The optimal small initial condition on the global existence of solutions for multi-species chemotaxis model in the fully parabolic situation had not been attained as far as the author knows. In this paper, we prove that under the sub-critical mass condition, any solutions to conflict-free system exist globally. Moreover, the global existence of solutions to system with strong self-repelling effect has been discussed even for large initial data. The proof is based on the modified free energy functional and the Moser–Trudinger inequality for system.
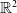
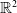
On the global existence of solutions to chemotaxis system for two populations in dimension two
- Part of
-
- Journal:
- Proceedings of the Royal Society of Edinburgh. Section A: Mathematics / Volume 153 / Issue 6 / December 2023
- Published online by Cambridge University Press:
- 09 January 2023, pp. 2106-2128
- Print publication:
- December 2023
-
- Article
- Export citation
-
We consider the global existence for the following fully parabolic chemotaxis system with two populations
\[\left\{ \begin{array}{@{}ll} \partial_tu_i=\kappa_i\Delta u_i-\chi_i\nabla\cdot(u_i\nabla v),\quad i\in\{1,2\}, & x\in\Omega,\ t>0, \\ v_t=\Delta v-v+u_1+u_2, & x\in\Omega,\ t>0,\\ u_i(x,t=0)=u_{i0}(x),\quad v(x,t=0)=v_0(x), & x\in\Omega, \end{array} \right. \]where $\Omega =\mathbb {R}^2$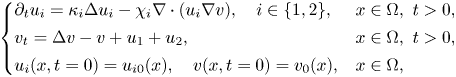
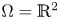 or $\Omega =B_R(0)\subset \mathbb {R}^2$
or $\Omega =B_R(0)\subset \mathbb {R}^2$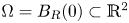 supplemented with homogeneous Neumann boundary conditions, $\kappa _i,\chi _i>0,$
supplemented with homogeneous Neumann boundary conditions, $\kappa _i,\chi _i>0,$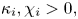 $i=1,2$
$i=1,2$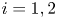 . The global existence remains open for the fully parabolic case as far as the author knows, while the existence of global solution was known for the parabolic-elliptic reduction with the second equation replaced by $0=\Delta v-v+u_1+u_2$
. The global existence remains open for the fully parabolic case as far as the author knows, while the existence of global solution was known for the parabolic-elliptic reduction with the second equation replaced by $0=\Delta v-v+u_1+u_2$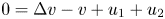 or $0=\Delta v+u_1+u_2$
or $0=\Delta v+u_1+u_2$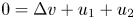 . In this paper, we prove that there exists a global solution if the initial masses satisfy the certain sub-criticality condition. The proof is based on a version of the Moser–Trudinger type inequality for system in two dimensions.
. In this paper, we prove that there exists a global solution if the initial masses satisfy the certain sub-criticality condition. The proof is based on a version of the Moser–Trudinger type inequality for system in two dimensions.
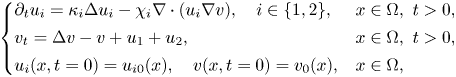
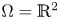
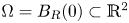
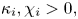
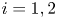
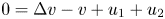
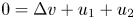
Finite-time blow-up in a repulsive chemotaxis-consumption system
- Part of
-
- Journal:
- Proceedings of the Royal Society of Edinburgh. Section A: Mathematics / Volume 153 / Issue 4 / August 2023
- Published online by Cambridge University Press:
- 06 June 2022, pp. 1150-1166
- Print publication:
- August 2023
-
- Article
- Export citation
-
In a ball $\Omega \subset \mathbb {R}^{n}$
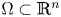 with $n\ge 2$
with $n\ge 2$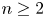 , the chemotaxis system
\[ \left\{ \begin{array}{@{}l} u_t = \nabla \cdot \big( D(u)\nabla u\big) + \nabla\cdot \big(\dfrac{u}{v} \nabla v\big), \\ 0=\Delta v - uv \end{array} \right. \]is considered along with no-flux boundary conditions for $u$
, the chemotaxis system
\[ \left\{ \begin{array}{@{}l} u_t = \nabla \cdot \big( D(u)\nabla u\big) + \nabla\cdot \big(\dfrac{u}{v} \nabla v\big), \\ 0=\Delta v - uv \end{array} \right. \]is considered along with no-flux boundary conditions for $u$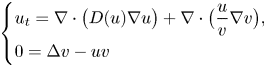
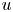 and with prescribed constant positive Dirichlet boundary data for $v$
and with prescribed constant positive Dirichlet boundary data for $v$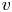 . It is shown that if $D\in C^{3}([0,\infty ))$
. It is shown that if $D\in C^{3}([0,\infty ))$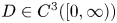 is such that $0< D(\xi ) \le {K_D} (\xi +1)^{-\alpha }$
is such that $0< D(\xi ) \le {K_D} (\xi +1)^{-\alpha }$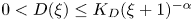 for all $\xi >0$
for all $\xi >0$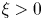 with some ${K_D}>0$
with some ${K_D}>0$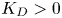 and $\alpha >0$
and $\alpha >0$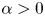 , then for all initial data from a considerably large set of radial functions on $\Omega$
, then for all initial data from a considerably large set of radial functions on $\Omega$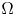 , the corresponding initial-boundary value problem admits a solution blowing up in finite time.
, the corresponding initial-boundary value problem admits a solution blowing up in finite time.
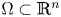
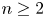
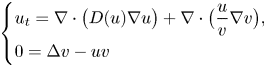
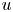
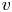
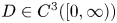
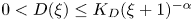
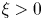
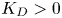
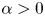
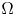
Stationary solutions to the Keller–Segel equation on curved planes
- Part of
-
- Journal:
- Proceedings of the Royal Society of Edinburgh. Section A: Mathematics / Volume 153 / Issue 1 / February 2023
- Published online by Cambridge University Press:
- 31 January 2022, pp. 327-343
- Print publication:
- February 2023
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
-
We study stationary solutions to the Keller–Segel equation on curved planes. We prove the necessity of the mass being $8 \pi$
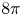 and a sharp decay bound. Notably, our results do not require the solutions to have a finite second moment, and thus are novel already in the flat case. Furthermore, we provide a correspondence between stationary solutions to the static Keller–Segel equation on curved planes and positively curved Riemannian metrics on the sphere. We use this duality to show the nonexistence of solutions in certain situations. In particular, we show the existence of metrics, arbitrarily close to the flat one on the plane, that do not support stationary solutions to the static Keller–Segel equation (with any mass). Finally, as a complementary result, we prove a curved version of the logarithmic Hardy–Littlewood–Sobolev inequality and use it to show that the Keller–Segel free energy is bounded from below exactly when the mass is $8 \pi$
and a sharp decay bound. Notably, our results do not require the solutions to have a finite second moment, and thus are novel already in the flat case. Furthermore, we provide a correspondence between stationary solutions to the static Keller–Segel equation on curved planes and positively curved Riemannian metrics on the sphere. We use this duality to show the nonexistence of solutions in certain situations. In particular, we show the existence of metrics, arbitrarily close to the flat one on the plane, that do not support stationary solutions to the static Keller–Segel equation (with any mass). Finally, as a complementary result, we prove a curved version of the logarithmic Hardy–Littlewood–Sobolev inequality and use it to show that the Keller–Segel free energy is bounded from below exactly when the mass is $8 \pi$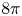 , even in the curved case.
, even in the curved case.
Chemotactic and temperature-dependent responses of the Strongyloidoidea superfamily of nematodes
-
- Journal:
- Parasitology / Volume 149 / Issue 1 / January 2022
- Published online by Cambridge University Press:
- 05 October 2021, pp. 116-123
-
- Article
- Export citation
Natural variation in host-finding behaviour of gastropod parasitic nematodes (Phasmarhabditis spp.) exposed to host-associated cues
-
- Journal:
- Journal of Helminthology / Volume 95 / 2021
- Published online by Cambridge University Press:
- 24 February 2021, e10
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
On a repulsion Keller–Segel system with a logarithmic sensitivity
- Part of
-
- Journal:
- European Journal of Applied Mathematics / Volume 33 / Issue 1 / February 2022
- Published online by Cambridge University Press:
- 14 January 2021, pp. 153-181
-
- Article
- Export citation
Immediate smoothing and global solutions for initial data in L1 × W1,2 in a Keller–Segel system with logistic terms in 2D
- Part of
-
- Journal:
- Proceedings of the Royal Society of Edinburgh. Section A: Mathematics / Volume 151 / Issue 4 / August 2021
- Published online by Cambridge University Press:
- 02 September 2020, pp. 1204-1224
- Print publication:
- August 2021
-
- Article
- Export citation
-
This paper deals with the logistic Keller–Segel model
in bounded two-dimensional domains (with homogeneous Neumann boundary conditions and for parameters χ, κ ∈ ℝ and μ > 0), and shows that any nonnegative initial data (u0, v0) ∈ L1 × W1,2 lead to global solutions that are smooth in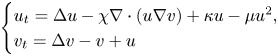 \[ \begin{cases} u_t = \Delta u - \chi \nabla\cdot(u\nabla v) + \kappa u - \mu u^2, \\ v_t = \Delta v - v + u \end{cases} \]
\[ \begin{cases} u_t = \Delta u - \chi \nabla\cdot(u\nabla v) + \kappa u - \mu u^2, \\ v_t = \Delta v - v + u \end{cases} \]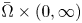 $\bar {\Omega }\times (0,\infty )$.
$\bar {\Omega }\times (0,\infty )$.
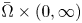
Does indirectness of signal production reduce the explosion-supporting potential in chemotaxis–haptotaxis systems? Global classical solvability in a class of models for cancer invasion (and more)
- Part of
-
- Journal:
- European Journal of Applied Mathematics / Volume 32 / Issue 4 / August 2021
- Published online by Cambridge University Press:
- 17 July 2020, pp. 618-651
-
- Article
- Export citation
7 - SSAs for Reaction–Diffusion–Advection Processes
-
- Book:
- Stochastic Modelling of Reaction–Diffusion Processes
- Published online:
- 04 November 2019
- Print publication:
- 30 January 2020, pp 192-225
-
- Chapter
- Export citation
The role of carbon dioxide in nematode behaviour and physiology
-
- Journal:
- Parasitology / Volume 147 / Issue 8 / July 2020
- Published online by Cambridge University Press:
- 11 October 2019, pp. 841-854
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
Gastropod parasitic nematodes (Phasmarhabditis sp.) are attracted to hyaluronic acid in snail mucus by cGMP signalling
-
- Journal:
- Journal of Helminthology / Volume 94 / 2020
- Published online by Cambridge University Press:
- 15 November 2018, e9
-
- Article
- Export citation
Sharp decay estimates in a bioconvection model with quadratic degradation in bounded domains
- Part of
-
- Journal:
- Proceedings of the Royal Society of Edinburgh. Section A: Mathematics / Volume 148 / Issue 5 / October 2018
- Published online by Cambridge University Press:
- 25 April 2018, pp. 939-955
- Print publication:
- October 2018
-
- Article
- Export citation
-
The paper studies large time behaviour of solutions to the Keller–Segel system with quadratic degradation in a liquid environment, as given by
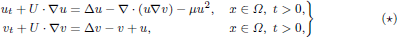
under Neumann boundary conditions in a bounded domain Ω ⊂ ℝn, where n ≥ 1 is arbitrary. It is shown that whenever U : Ω × (0,∞) → ℝn is a bounded and sufficiently regular solenoidal vector field any non-trivial global bounded solution of (⋆) approaches the trivial equilibrium at a rate that, with respect to the norm in either of the spaces L1(Ω) and L∞(Ω), can be controlled from above and below by appropriate multiples of 1/(t + 1). This underlines that, even up to this quantitative level of accuracy, the large time behaviour in (⋆) is essentially independent not only of the particular fluid flow, but also of any effect originating from chemotactic cross-diffusion. The latter is in contrast to the corresponding Cauchy problem, for which known results show that in the n = 2 case the presence of chemotaxis can significantly enhance biomixing by reducing the respective spatial L1 norms of solutions.
Thermodynamically consistent Navier–Stokes–Cahn–Hilliard models with mass transfer and chemotaxis
-
- Journal:
- European Journal of Applied Mathematics / Volume 29 / Issue 4 / August 2018
- Published online by Cambridge University Press:
- 09 October 2017, pp. 595-644
-
- Article
- Export citation
