This journal utilises an Online Peer Review Service (OPRS) for submissions. By clicking "Continue" you will be taken to our partner site https://rse.msp.org/submit_new.php?j=rse_proc_a_math. Please be aware that your Cambridge account is not valid for this OPRS and registration is required. We strongly advise you to read all "Author instructions" in the "Journal information" area prior to submitting.
This page lists all time most cited articles for this title. Please use the publication date filters on the left if you would like to restrict this list to recently published content, for example to articles published in the last three years. The number of times each article was cited is displayed to the right of its title and can be clicked to access a list of all titles this article has been cited by.
Using the ‘monotonicity trick’ introduced by Struwe, we derive a generic theorem. It says that for a wide class of functionals, having a mountain-pass (MP) geometry, almost every functional in this class has a bounded Palais-Smale sequence at the MP level. Then we show how the generic theorem can be used to obtain, for a given functional, a special Palais–Smale sequence possessing extra properties that help to ensure its convergence. Subsequently, these abstract results are applied to prove the existence of a positive solution for a problem of the form
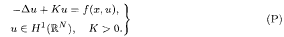
We assume that the functional associated to (P) has an MP geometry. Our results cover the case where the nonlinearity f satisfies (i) f(x, s)s−1 → a ∈)0, ∞) as s →+∞; and (ii) f(x, s)s–1 is non decreasing as a function of s ≥ 0, a.e. x → ℝN.
In a series of papers W. F. Sheppard (1912, 1914) has considered the approximate representation of equidistant, equally weighted, and uncorrelated observations under the following assumptions:–
(i) The data being u1, u2, …, un, the representation is to be given by linear combinations
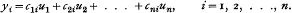
(ii) The linear combinations are to be such as would reproduce any set of values that were already values of a polynomial of degree not higher than the kth.
(iii) The sum of squared coefficients 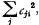 which measures the mean square error of yi, is to be a minimum for each value of i.
which measures the mean square error of yi, is to be a minimum for each value of i.
We study the existence of positive solutions for the nonlinear Schrödinger equation with the fractional Laplacian
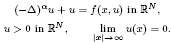
Furthermore, we analyse the regularity, decay and symmetry properties of these solutions.
The frequency ratio of the allelomorphs of a Mendelian factor is only stable if selection favours the heterozygote: such factors, though occurring rarely, will accumulate in the stock, while those of opposite tendency will be eliminated.
The survival of a mutant gene although established in a mature and potent individual is to a very large extent a matter of chance ; only when a large number of individuals have become affected does selection, dependent on its contribution to the fitness of the organism, become of importance. This is so even for dominant mutants; for recessive mutants selection remains very small so long as the mutant form is an inconsiderable fraction of the interbreeding group.
The distribution of the frequency ratio for different factors may be calculated from the condition that this distribution is stable, as is that of velocities in the Theory of Gases : in the absence of selection the distribution of log 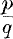 is given in fig. 1. Fig. 2 represents the case of steady decay in variance by the action of random survival (the Hagedoorn effect).
is given in fig. 1. Fig. 2 represents the case of steady decay in variance by the action of random survival (the Hagedoorn effect).
Fig. 3 shows the distribution in the somewhat artificial case of uniform genetic selection : this would be the distribution to be expected in the absence of dominance. Fig. 4 shows the asymmetrical distribution due to uniform genotypic selection with or without homogamy.
Under genotypic selection the dominance ratio for complete dominance comes to be exactly ⅓, in close agreement with the value obtained from human measurements.
The rate of mutation necessary to maintain the variance of the species may be calculated from these distributions. Very infrequent mutation will serve to counterbalance the effect of random survival; for equilibrium with selective action a much higher level is needed, though still mutation may be individually rare, especially in large populations.
It would seem that the supposition of genotypic selection balanced by occasional mutations fitted the facts deduced from the correlations of relatives in mankind.
1. Integrals of the form  where ψ(x) is a function with a limited number of turning-points in the range of integration and k is a constant which may take up large values, frequently occur in investigations in mathematical physics, and their computation by quadratures is often desirable.
where ψ(x) is a function with a limited number of turning-points in the range of integration and k is a constant which may take up large values, frequently occur in investigations in mathematical physics, and their computation by quadratures is often desirable.
The generalisation to the stationary case of the solution given by Weyl (1918) of Einstein's gravitational equations in a statical axially symmetric universe, has been attempted by various writers. It has not, however, so far been possible to reduce the equations to linear form and thus to find the most general solution. A set of special solutions has been obtained by Lewis (1932), valid in a region free of matter, which contain Weyl's solution as a particular case. They depend upon an arbitrary solution of the equation
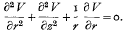
1. When a battery of intelligence tests is administered to a set of persons it is a common practice among psychologists to “explain” the scores obtained in terms of a number of “factors.” Thus if we suppose that there are altogether t tests and that xi, denotes the score of any one person in the ith test, then it is assumed that
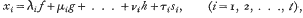
where f, g, …, h represent the person's measures in one or more general or group factors, and Si, is the person's specific ability in the ith test. It is further assumed that for a given and supposed infinite population of persons all the factors, specific and otherwise, are distributed normally and independently and that they are standardised, i.e. that their standard deviations are unity.
Consider the second order Hamiltonian system:

where q ∊ ℝn and V ∊ C1 (ℝ ×ℝn ℝ) is T periodic in t. Suppose Vq (t, 0) = 0, 0 is a local maximum for V(t,.) and V(t, x) | x| → ∞ Under these and some additional technical assumptions we prove that (HS) has a homoclinic orbit q emanating from 0. The orbit q is obtained as the limit as k → ∞ of 2kT periodic solutions (i.e. subharmonics) qk of (HS). The subharmonics qk are obtained in turn via the Mountain Pass Theorem.
We construct local minimisers to certain variational problems. The method is quite general and relies on the theory of Γ-convergence. The approach is demonstrated through the model problem
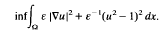
It is shown that in certain nonconvex domains Ω ⊂ ℝn and for ε small, there exist nonconstant local minimisers uε satisfying uε ≈ ± 1 except in a thin transition layer. The location of the layer is determined through the requirement that in the limit uε →u0, the hypersurface separating the states u0 = 1 and u0 = −1 locally minimises surface area. Generalisations are discussed with, for example, vector-valued u and “anisotropic” perturbations replacing |∇u|2.

