



Introduction
Coronavirus disease-2019 (Covid-19) was declared a global pandemic by the World Health Organization (WHO) on 11 March 2020, leading to a variety of responses by different governments around the world. The nonpharmaceutical intervention strategy of New Zealand was particularly stringent and effectively eliminated a burgeoning Covid-19 outbreak through a nationwide lockdown, physical distancing, improved hand hygiene and ongoing 14-day quarantine for returning travellers (Robert, Reference Robert2020; Baker et al., Reference Baker, Kvalsvig and Verrall2020a). A four-level alert system (New Zealand Government, 2020) was introduced on 21 March, beginning at level 2 and quickly moving to level 3 on 23 March and level 4 (i.e. the entire nation goes into self-isolation) on 25 March. The level 4 lockdown remained in place until 27 April, when it was reduced to level 3. By 11 May it was lowered to level 2 and finally decreased to level 1 on 8 June (Baker et al., Reference Baker, Wilson and Anglemyer2020b). The primary intention of these measures in New Zealand was to reduce the transmission of Covid-19, but they also brought about a reduction in influenza and other respiratory viral infections in 2020 (Huang et al., Reference Huang, Wood, Jelley, Jennings, Jefferies, Daniells, Nesdale, Dowell, Turner, Campbell-Stokes, Balm, Dobinson, Grant, James, Aminisani, Ralston, Gunn, Bocacao, Danielewicz, Moncrieff, McNeill, Lopez, Waite, Kiedrzynski, Schrader, Gray, Cook, Currin, Engelbrecht, Tapurau, Emmerton, Martin, Baker, Taylor, Trenholme, Wong, Lawrence, McArthur, Stanley, Roberts, Rahnama, Bennett, Mansell, Dilcher, Werno, Grant, van der Linden, Youngblood, Thomas, Webby and Consortium2021). Similar impacts have also been observed elsewhere (Fricke et al., Reference Fricke, Glöckner, Dreier and Lange2021; Yang et al., Reference Yang, Xiao, Gu, Liang, Cao, Mou, Huang, Chen and Liu2021), but little attention to date has been given to parasitic enteric diseases.
Cryptosporidium species are obligatory intracellular intestinal parasites that infect a wide range of vertebrate hosts, causing a considerable burden of gastrointestinal disease (Xiao et al., Reference Xiao, Fayer, Ryan and Upton2004; Garcia and Hayman, Reference Garcia and Hayman2016). In New Zealand, cryptosporidiosis is a notifiable disease with a higher incidence compared to other developed countries (Learmonth et al., Reference Learmonth, Ionas, Ebbett and Kwan2004). Two species, C. hominis and C. parvum are responsible for the majority of human infections globally, and in New Zealand (Garcia-R and Hayman, Reference Garcia-R and Hayman2017; Garcia-R et al., Reference Garcia-R, French, Pita, Velathanthiri, Shrestha and Hayman2017; Garcia-R et al., Reference Garcia-R, Pita, Velathanthiri, French and Hayman2020b). Of these, C. hominis is transmitted primarily through human-to-human contact whereas C. parvum is mainly zoonotic with ruminants as the primary reservoir host [(Learmonth et al., Reference Learmonth, Ionas, Ebbett and Kwan2004; Feltus et al., Reference Feltus, Giddings, Schneck, Monson, Warshauer and McEvoy2006; Leitch and He, Reference Leitch and He2011) though see King et al. (Reference King, Tyler and Hunter2019)]. Clear seasonal peaks in cryptosporidiosis are present in New Zealand. An early autumnal peak for C. hominis associated with increased recreational water use and a spring peak for C. parvum associated with dairy cattle calving season is observed each year (Learmonth et al., Reference Learmonth, Ionas, Ebbett and Kwan2004; Lake et al., Reference Lake, Pearce and Savill2008; Garcia-R et al., Reference Garcia-R, Pita, Velathanthiri, French and Hayman2020b). Transmission of both species is through the faecal−oral route, following contact with infected hosts or water contaminated with the transmissive stages (oocysts) of the organism (Bouzid et al., Reference Bouzid, Hunter, Chalmers and Tyler2013).
One of the difficulties with monitoring cryptosporidiosis is the lack of morphological characters to identify different Cryptosporidium species. Frequently, diagnosis is based on microscopy by the presence of oocysts in stool samples of infected patients or enzyme immunoassay and species-level identification is rarely attempted. Reliable species and subtype level identification is possible through molecular methods, though diagnostic laboratories moving to molecular diagnostics typically utilize qPCR and report presence/absence, without speciation. The most widely used marker for this purpose is the 60 kDa glycoprotein locus (gp60), upon which the subtyping of Cryptosporidium species is based (Feng and Xiao, Reference Feng and Xiao2017). This classification scheme has been used to study the epidemiology of human outbreaks of cryptosporidiosis in many previous studies (O'Brien et al., Reference O'Brien, McInnes and Ryan2008; Chalmers et al., Reference Chalmers, Robinson, Elwin and Elson2019). Here, we examine the disease dynamics of Cryptosporidium infections using gp60 for species identification and subtype family diversity patterns before, during and after lockdown measures in New Zealand.
Materials and methods
Anonymous Cryptosporidium-positive stool samples were collected between 2015 and 2021 through routine public health investigation activities from symptomatic humans visiting private and public general practitioner (GP) surgeries and hospitals throughout New Zealand. Samples were sent from accredited diagnostic laboratories to the Protozoa Research Unit at Hopkirk Research Institute (Massey University) for molecular analyses under a Ministry of Health contract. Stools were collected in 10-ml screw-cap tubes and stored at 4°C in the laboratory until DNA extraction.
DNA extraction was carried out 1–2 weeks after receiving samples at the Hopkirk Research Institute. Before DNA extraction, the oocysts of Cryptosporidium were physically disrupted using a beadbeater (Tissue Lyser II, Qiagen) at 30 Hz for 5 min. DNA extractions were performed using Zymo Quick-DNA Fecal/Soil Microbe kits following the manufacturer's instructions. A fragment of the gp60 gene was amplified using a nested PCR (Alves et al., Reference Alves, Xiao, Sulaiman, Lal, Matos and Antunes2003; Waldron et al., Reference Waldron, Ferrari and Power2009) and amplification products were sequenced in both directions using Big Dye Terminator version 3.1 reagents on an ABI 3730XL automated DNA sequencer (Applied Biosystems, Foster City, California, USA).
Cryptosporidium gp60 sequences were classified into species and subtype families according to guidelines in Feng and Xiao (Reference Feng and Xiao2017). While our sampling prior to 2020 included a wide range of regions throughout New Zealand, the 2020 samples were dominated by the Auckland region (118/130). For this reason, we filtered out results from elsewhere and limited our analyses to samples collected from the Auckland region. To check the representativeness of our dataset, we compared it with data from the national notifiable disease surveillance website (https://surv.esr.cri.nz/surveillance/annual_diseasetables.php).
Results
Our dataset included 1502 sequences received from 9 January 2015 to 3 March 2021 (Supplementary Table 1). Of these, C. parvum was the most common (n = 868), followed by C. hominis (598), Cryptosporidium cuniculus (28) and Cryptosporidium erinacei (8). Seasonal dynamics of C. hominis and C. parvum before 2020 and the impact of the Covid-19 lockdown are illustrated in Fig. 1. Cryptosporidium hominis was commonly found in the first (Q1) and second quarters (Q2) of the year between January and June while C. parvum was more common in the third (Q3) and fourth quarters (Q4) from July to December. The exception to this is in 2020, when no C. hominis was detected after 14 February 2020 (Fig. 1).

Fig. 1. Cryptosporidium hominis and C. parvum quarterly cases from Auckland, New Zealand 2015–2021. Dotted line in Q1 2020 indicates the start of Covid-19 national lockdown.
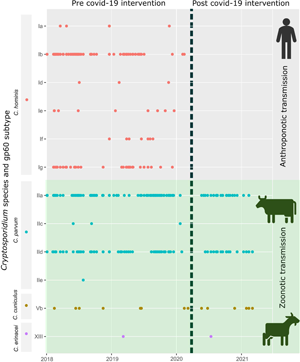
Further examination of gp60 subtype families shows the diversity and temporal dynamics of C. hominis and C. parvum (Fig. 2). Within C. hominis, subtype family Ib was detected consistently between 2015 and the beginning of 2020. Other gp60 subtype families in C. hominis appeared as sporadic outbreaks e.g. Id in 2016, Ig in 2017, 2018 and 2019 and/or single cases e.g. Ia in 2015–2019. Within C. parvum, subtype families IIa and IId were consistently detected whilst IIc and IIe appeared as sporadic single cases. Of the other zoonotic Cryptosporidium species, C. cuniculus and C. erinacei were only detected occasionally.

Fig. 2. Timeline of Cryptosporidium gp60 subtype cases identified from Auckland, New Zealand 2015–2021. Number of cases per species are indicated on the y axis. Dotted line in 2020 indicates the start of Covid-19 national lockdown.
Our data are representative of the total number of cases reported nationally from the notifiable disease database from 2015 to 2019 (between 33% and 46%, Supplementary Table 1). However, in 2020 the number of cases reported to our lab dropped to a lower proportion (18%). This was probably caused by extra stress on diagnostic laboratories that did not send as many samples to our lab for molecular analyses during and following the Covid-19 outbreak. Data from outside Auckland are not included in our analyses due to low numbers in 2020, but in the few samples from outside Auckland which we do have, no post lockdown cases of C. hominis have been reported.
Discussion
In response to Covid-19, New Zealand underwent a nationwide lockdown beginning in late March 2020, with households spending over 7 weeks at home except for essential personal movements like supermarket and hospital visits. Since this time, our surveillance has revealed that C. hominis has not been detected in reported cases of cryptosporidiosis from the Auckland region. This absence has continued until the time of publication during the first quarter of 2021, which in previous years has been the beginning of peak season for C. hominis infections. In contrast, C. parvum and other less common zoonotic Cryptosporidium species have continued typical seasonal patterns. Our data suggest that the national measures for responding to the Covid-19 also interrupted C. hominis transmission. For instance, the closure of all child care centres and schools may have limited C. hominis transmission since these have been linked to previous outbreaks (Goñi et al., Reference Goñi, Almagro-Nievas, Cieloszyk, Lóbez, Navarro-Marí and Gutiérrez-Fernández2015). Ongoing managed isolation of visitors or citizens returning to New Zealand since then has also likely reduced new imported cases of cryptosporidiosis.
Prior to 2020, infections by C. hominis were reported seasonally (Garcia-R and Hayman, Reference Garcia-R and Hayman2017; Garcia-R et al., Reference Garcia-R, Pita, Velathanthiri, French and Hayman2020b). Furthermore, temporal dynamics of gp60 subtype families generally revealed sporadic infection patterns. Subtype families Ia, Id, Ie, If and Ig appeared at irregular intervals throughout the study period, which may indicate an importation by visitors or returning travellers from a global pool of C. hominis that fail to establish within New Zealand or an opportunistic behaviour of the parasite (Garcia-R and Hayman, Reference Garcia-R and Hayman2017; Garcia-R et al., Reference Garcia-R, Cox and Hayman2020a). The majority of C. hominis cases was subtype family Ib, which is also the most common subtype family globally (Feng et al., Reference Feng, Li, Duan and Xiao2009; Putignani and Menichella, Reference Putignani and Menichella2010; Segura et al., Reference Segura, Prim, Montemayor, Valls and Muñoz2015). These findings imply that despite the hardiness of Cryptosporidium oocysts in the environment (Carey et al., Reference Carey, Lee and Trevors2004), historical C. hominis infections in New Zealand arise through importation and that the levels of sanitation and hand hygiene, as well as a low-density population, may be sufficient to prevent the establishment of endemic C. hominis infections. Similarly, improved hand hygiene on farms, especially during calving season may help control zoonotic C. parvum transmission in New Zealand (Thomas-Lopez et al., Reference Thomas-Lopez, Müller, Vestergaard, Christoffersen, Andersen, Jokelainen, Agerholm and Stensvold2020).
C. parvum sequences were dominated by gp60 subtype families IIa and IId, which both occurred consistently throughout the study period, including during 2020. These C. parvum subtype families are primarily associated with ruminants and transmitted through them to humans (Mammeri et al., Reference Mammeri, Chevillot, Chenafi, Thomas, Julien, Vallée, Polack, Follet and Adjou2019; Nader et al., Reference Nader, Mathers, Ward, Pachebat, Swain, Robinson, Chalmers, Hunter, van Oosterhout and Tyler2019). Both have been detected in cattle and sheep in New Zealand (Garcia-R et al., Reference Garcia-R, French, Pita, Velathanthiri, Shrestha and Hayman2017) and evidence from overseas suggests that subtype family IId may be more prevalent in sheep and goat populations (Quílez et al., Reference Quílez, Torres, Chalmers, Hadfield, Del Cacho and Sánchez-Acedo2008), but further research is needed to identify if host-specific C. parvum subtype families occur in New Zealand's domestic animals. Overall, C. parvum transmission was not interrupted by Covid-19 control measures except perhaps by a small reduction in peak numbers during 2020 Q3 and Q4. Subtype family IIc is rare in New Zealand, with only six cases in our dataset. Previous research shows that unlike other C. parvum subtype families, IIc is an anthroponotic (human-adapted) and a potential emerging genotype (King et al., Reference King, Tyler and Hunter2019; Nader et al., Reference Nader, Mathers, Ward, Pachebat, Swain, Robinson, Chalmers, Hunter, van Oosterhout and Tyler2019). While no cases of subtype family IIc were detected after Covid-19 control measures, its rarity before Covid-19 prevents any strong conclusions from our data. Further investigation of gp60 diversity, i.e. trinucleotide repeat regions, may have revealed more details of the transmission patterns of cryptosporidiosis in New Zealand, but this would have been difficult to assess since we do not have descriptive epidemiology or exposure data for our cases. In C. parvum, previous research has demonstrated strong associations between animal contact and cryptosporidiosis, though human-to-human transmission is also possible (Chalmers et al., Reference Chalmers, Robinson, Elwin and Elson2019). However, like C. hominis, we suggest that human to human C. parvum outbreaks in New Zealand are contained through basic sanitation measures.
Our study has limitations in the methodology of data collection stemming from significant changes to human behaviours during 2020 which may have influenced our results. The number of samples we have for 2020/2021 is lower than in previous years and adds uncertainty in the data. As well as potentially being due to a reduction in transmission and cases, since the symptoms of cryptosporidiosis do not closely match Covid-19, people with cryptosporidiosis may have been less likely to visit doctors immediately prior to, during and following Covid-19 restrictions compared with previous years. Furthermore, because symptoms usually resolve themselves without treatment, cryptosporidiosis is an under-reported disease throughout the developed world (Dreesman et al., Reference Dreesman, Villarroel-Conzales, Cleves, Reins and Pulz2007; Scallan et al., Reference Scallan, Hoekstra, Angulo, Tauxe, Widdowson, Roy, Jones and Griffin2011). Despite being a notifiable disease in New Zealand it is likely that only a small proportion of cases are diagnosed, thus raising the potential for bias in reporting different Cryptosporidium species. The severity of cryptosporidiosis is impacted by many factors including host (concurrent infection, malnutrition, immunosuppression, age), environmental (dose of exposure) as well as species and genotype of Cryptosporidium (Weir, Reference Weir2001). Of the species in our study, C. hominis is associated with more severe infections and higher shedding rates (Bushen et al., Reference Bushen, Kohli, Pinkerton, Dupnik, Newman, Sears, Fayer, Lima and Guerrant2007), potentially making reporting and detection of C. hominis more likely than C. parvum. Once cryptosporidiosis cases are confirmed, the diagnostic laboratories send stool samples to our lab for genetic analyses, but this may not be nationally representative as evidenced by the lack of samples from outside the Auckland region during 2020. The reasons for this are unclear but may reflect a focus on essential services and over-stressed health systems not having time to send samples since the start of Covid-19.
In conclusion, our study revealed that nonpharmaceutical interventions greatly reduced C. hominis in Auckland, New Zealand. Since the March 2020 national lockdown, travel restrictions and mandatory visitor isolation have kept reported C. hominis numbers at zero. Ongoing monitoring of Cryptosporidium species and gp60 subtype families as international travel restrictions ease will provide further information on cryptosporidiosis in New Zealand. Finally, to further investigate the preliminary findings presented here we recommend future studies focus on descriptive epidemiology and exposure data for cases of human cryptosporidiosis in New Zealand.
Supplementary material
The supplementary material for this article can be found at https://doi.org/10.1017/S0031182021000974.
Data
Sequence data availability is outlined in Garcia-R et al. (2020b).
Author contribution
DH conceived and designed the study. PO, AP and NV conducted data gathering. JG and MK performed analyses. MK wrote the original draft article and all coauthors contributed to editing.
Financial support
The authors thank the New Zealand Ministry of Health for funding. DH and MK acknowledge funding from the Royal Society Te Apārangi Rutherford Discovery Fellowship (MAU1701).
Conflicts of interest
The authors declare there are no conflicts of interest.
Ethical standards
Not applicable.





 Open access
Open access