



1. Introduction
Tissue regeneration, or the re-establishment of the form and function of a damaged or lost structure, is an example of post-embryonic morphogenesis. The history of regeneration research is long and rich in breakthroughs (Dinsmore, Reference Dinsmore1991), and some of the key molecular and mechanical details have been understood in recent decades (Elchaninov et al., Reference Elchaninov, Sukhikh and Fatkhudinov2021; Ikeuchi et al., Reference Ikeuchi, Ogawa, Iwase and Sugimoto2016; Liu et al., Reference Liu, Zhu and Xiao2023; Morinaka et al., Reference Morinaka, Sakamoto, Iwase and Sugimoto2023; Sugimoto et al., Reference Sugimoto, Temman, Kadokura and Matsunaga2019).
The role of cell proliferation in the re-establishment of lost structures has long been recognized as central to the process of regeneration (Morgan, Reference Morgan1901). At the most fundamental level, there are basic yet unanswered questions regarding the type of dynamics and the parameters controlling it. For example, does cell proliferation during regeneration follow unique dynamics, distinguished from the ones driving other types of morphodynamics such as embryonic development or post-embryonic organogenesis, including metamorphosis in animals or flower formation in plants? Is regeneration a smooth process, or does it go through sharp transitions, perhaps analogous to phase transitions observed in many complex dynamical systems? Unfortunately, more than a hundred years after the first observations, a complete quantitative description of cell proliferation dynamics during organ regeneration is lacking, impeding our efforts to understand how biological shapes and functions are established and maintained.
Here we present a quantitative analysis of cell divisions in regenerating root tips of the plant model system Arabidopsis thaliana. Given the relatively long duration of root regeneration following full tip excision (Sena et al., Reference Sena, Wang, Liu, Hofhuis and Birnbaum2009), we adopted light-sheet microscopy for sustained, high-resolution, time-lapse imaging. In plants, this method had been previously adapted first to Arabidopsis roots (Maizel et al., Reference Maizel, von Wangenheim, Federici, Haseloff and Stelzer2011; Sena et al., Reference Sena, Frentz, Birnbaum and Leibler2011) and then to other tissues (Berthet & Maizel, Reference Berthet and Maizel2016; Clark et al., Reference Clark, Van den Broeck, Guichard, Stager, Tanner, Blilou, Grossmann, Iyer-Pascuzzi, Maizel, Sparks and Sozzani2020).
Quantitative analyses of cell divisions in intact and regenerating Arabidopsis roots have a long history. Modern imaging methods span from simple light microscopy (Beemster & Baskin, Reference Beemster and Baskin1998) to confocal microscopy (Campilho et al., Reference Campilho, Garcia, Toorn, Wijk, Campilho and Scheres2006; Lavrekha et al., Reference Lavrekha, Pasternak, Ivanov, Palme and Mironova2017; Rahni & Birnbaum, Reference Rahni and Birnbaum2019) and light-sheet microscopy (Buckner et al., Reference Buckner, Madison, Chou, Matthiadis, Melvin, Sozzani, Williams and Long2019; de Luis Balaguer et al., Reference de Luis Balaguer, Ramos-Pezzotti, Rahhal, Melvin, Johannes, Horn and Sozzani2016; Sena et al., Reference Sena, Frentz, Birnbaum and Leibler2011; von Wangenheim et al., Reference von Wangenheim, Fangerau, Schmitz, Smith, Leitte, Stelzer and Maizel2016), but no comparison has been attempted between these dynamics and those in regenerating roots.
Algorithms to track cell divisions in light-sheet microscopy 4D datasets have been developed multiple times (Amarteifio et al., Reference Amarteifio, Fallesen, Pruessner and Sena2021; Buckner et al., Reference Buckner, Madison, Chou, Matthiadis, Melvin, Sozzani, Williams and Long2019; Sena et al., Reference Sena, Frentz, Birnbaum and Leibler2011). For this work, we adopted hardware and software previously developed in our lab (Amarteifio et al., Reference Amarteifio, Fallesen, Pruessner and Sena2021; Baesso et al., Reference Baesso, Randall and Sena2018).
By comparing the dynamics of cell proliferation in a growing intact root with that in a regenerating one, in this work we address the following fundamental questions: Is there a quantitative difference between the dynamics of cell division in an uncut root and that in a regenerating one? Is there a clear transition between different ‘phases’ in cell division dynamics during root regeneration?
2. Results
2.1. Temporal sequence of mitotic events is intermittent
The cyclin-dependent protein kinase CYCB1;1 is commonly used as a reporter of the G2/M transition in the cell cycle and, indirectly, of mitotic events (Reddy et al., Reference Reddy, Heisler, Ehrhardt and Meyerowitz2004). Transgenic Arabidopsis plants expressing CYCB1;1::GFP (Reddy et al., Reference Reddy, Heisler, Ehrhardt and Meyerowitz2004) were mounted on an open hardware light-sheet microscope setup (Baesso et al., Reference Baesso, Randall and Sena2018) specifically designed for imaging and tracking the meristematic region of a single root tip every 15 minutes (Methods and Supplementary Figure S1).
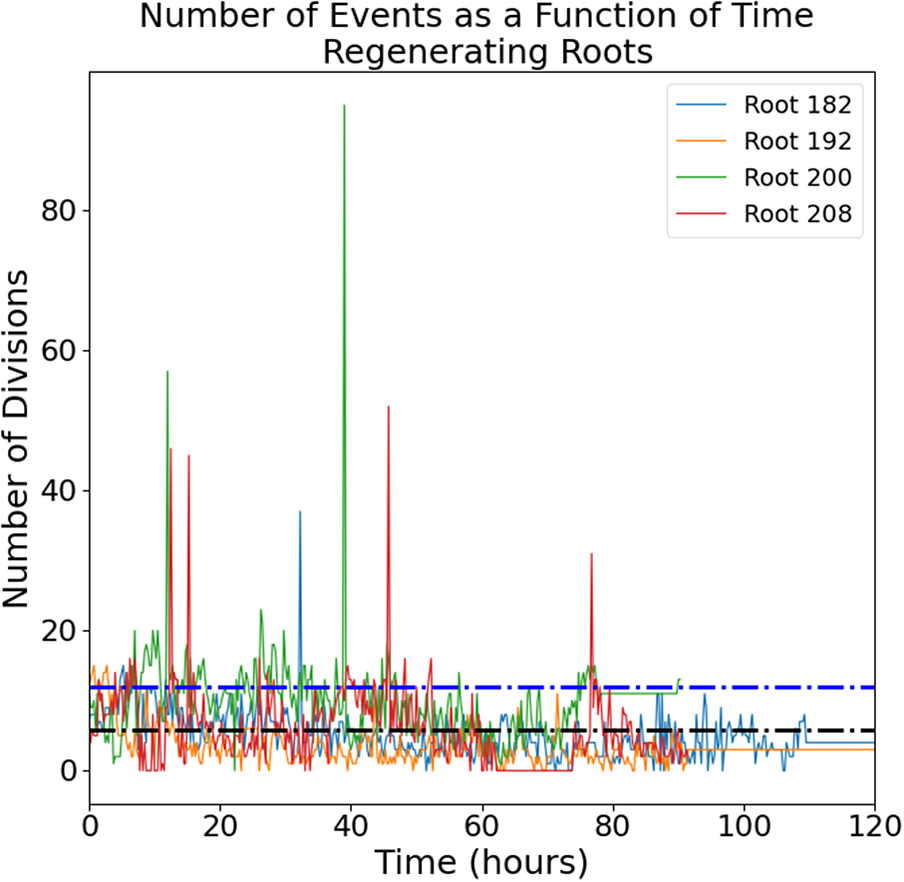
The raw images (a representative time-lapse is shown in Supplementary Figure S2) were processed using our previously published routine (Amarteifio et al., Reference Amarteifio, Fallesen, Pruessner and Sena2021) to track and count the mitotic events in 3D. The number of cell divisions detected in each frame follows an intermittent temporal pattern with a noisy baseline below 20 events per frame punctuated by a few isolated bursts of much higher activity (Figure 1).

Figure 1. Number of mitotic events detected in intact (left panel) and regenerating (right panel) root tips. Four independent roots are shown for each group. Black dotted line, mean; blue dotted line, mean + standard deviation.
2.2. Regenerating and intact roots exhibit different distributions of temporal ‘bursts’ of mitotic events
The intermittent nature of the temporal series in Figure 1 is interesting and can be further quantified. We define a ‘burst’ as a significant peak in the temporal series. More specifically, it is a collection of mitotic events occurring in a single time-point and at least one standard deviation higher than the mean of events observed in the entire temporal series; this value was calculated to be 10.95 events for intact roots and 11.63 events for regenerating roots. The size of the burst is simply the total number of cell divisions captured at that time point. The two distributions of burst sizes for intact and regenerating roots are significantly different (Figure 2; K-S test, p < 0.01) and indicate that regeneration is on average characterized by larger bursts of cell division activity.

Figure 2. Distribution of burst size (i.e. number of division events in that burst) in intact and regenerating root tips. Experimental data (histograms) and Poisson distributions peaking at 11 (yellow) and 13 (blue) burst sizes.
If the mitotic events were completely uncorrelated from each other, these distributions would be indistinguishable from Poisson distributions. This is not what we observed. The Poisson distribution looks very different from the experimental distribution with the same maximum, both for intact and regenerating roots (Figure 2).
2.3. Regenerating and intact roots exhibit different periodicities of mitotic events
To reveal hidden periodicities in the pattern, we generated a periodogram or a standard spectral analysis of the temporal series of single mitotic events (see Methods). Briefly periodograms show a distribution of fundamental periodicities in a time series. Our analysis indicates strong fundamental periodicities corresponding to approximately 4, 6 and 24 hours for intact roots and 11 and 16 hours for regenerating roots (Figure 3). Since we enforced a 24-hour light cycle (16-hour light: 8-hour dark) on all the plants during germination, periodicities of 24 hours and its subdivisions (e.g. 12, 6, 4, etc.) might be expected and trivial. On the other hand, the peak at approximately 16 hours observed in the periodogram of regenerating roots, and not in that of intact roots, suggests a nontrivial periodicity specific to the regeneration process.

Figure 3. Periodogram of the temporal series shown in Figure 1 for intact and regenerating root tips. PSD, power spectral density. Red arrow, 16-hour period in regenerating roots, suggesting nontrivial periodicity.
Although the cause of these periodicities remains unclear, the spectral analysis suggests fundamental differences in the cell division dynamics in unperturbed and regenerating tissues.
2.4. Difference in the distribution of mitotic events per frame between regenerating and intact roots emerges only 24 hours after excision
To further characterize the dynamics of mitotic events in both intact and regenerating roots, we compared the distributions of mitotic events in each frame, that is, the probabilities of detecting a mitotic event at a single time point (Figure 4). The distributions for intact and regenerating roots are significantly different over the entire duration of our observation (Figure 4A; K-S test, p < 0.001), further supporting the hypothesis that the underlying dynamics of cell divisions are different for intact roots than for regenerating roots.
While both distributions peak at approximately 3.5 divisions per frame and are skewed towards higher values, the regenerating root distribution shows a ‘shoulder’ of approximately 11 divisions per frame, which is not as evident in the intact root sample (Figure 4A). This suggests the existence of two unresolved subpopulations of events in the regenerating roots: one with a maximum of approximately 3.5 divisions per frame, as in the intact roots, and a second one centred at approximately 11 divisions per frame. This second peak is unmatched in the data from intact roots, suggesting a unique feature of self-organizing tissue.
To address whether root regeneration is a single continuous process or, instead, is made of distinct developmental phases, we asked whether the highly active time-points with 11 divisions per frame occurred throughout the entire regeneration process or only at specific moments.
We re-analysed the data into temporal bins, 0–6 hours, 6–24 hours, 24–72 hours and >72 hours after the excision. The distributions of divisions per frame are statistically indistinguishable between intact and regenerating roots during the first 6 hours (Figure 4B; K-S test, p = 0.21) and between 6 and 24 hours (Figure 4C; K-S test, p = 0.83). Crucially, between 24 and 72 hours after excision, the two distributions are marginally significantly different (Figure 4D; K-S test, p = 0.025), with the one for regenerating roots showing a longer tail between 10 and 20 divisions per frame. Finally, the two distributions remained significantly different 72 hours after excision (Figure 4E; K-S test, p < 0.001). Taken together, these data indicate that the main difference in cell division dynamics between regenerating and intact roots appears only 24 hours after tip excision, with the regenerating roots showing an excess of 10–15 events per time-point.

Figure 4. Distributions of mitotic events detected in one frame in intact and regenerating root tips. (A) all events; (B) events detected in the first 6 hours; (C) events detected between 6 and 24 hours; (D) events detected between 24 and 72 hours; (E) events detected after 72 hours. Histograms, experimental data lines and kernel density estimation of the experimental data (smooth fitting).

Figure 5. Distribution of cluster numbers and their size at a single time-point. The size of each point represents its frequency or how often that point appears in the data. (A) Intact roots; (B) regenerating roots.
2.5. Mitotic events occur in small spatial clusters that are more abundant in regenerating roots
The lack of a persistent reference point across time frames makes the spatial localization of the mitotic event relative to biologically significant landmarks in the root intractable. Instead, the spatial information allows the calculation of the relative distance between events. One important question from the developmental point of view is whether these occur uniformly within the tissue or rather in clusters.
To define a spatial cluster of events, we first determined the centre of mass of each event using our tracking algorithm (Amarteifio et al., Reference Amarteifio, Fallesen, Pruessner and Sena2021). Around each centre of mass, we modelled a 6 X 4 X 4 μm cuboid cell with a maximum diagonal, or ‘diameter’, equal to 8.24 μm. We used the DBScan algorithm (Ester et al., Reference Ester, Kriegel, Sander and Xu1996) to identify all events within three cell diameters (ε = 3 x 8.24 μm = 24.72 μm) from each other as part of a single cluster. Finally, we plot the distribution of cluster sizes, or how many clusters of which size we detected at a single time-point, for the populations of intact and regenerating roots (Figure 5). In both distributions, most of the time-points contain 1–3 spatial clusters made of 2–4 events each, but at any given time, regenerating roots are more likely to contain a higher number of clusters (up to 4–5) of the same 2–4 cell size (Figure 5). This can be seen by noting the slightly larger size of the dots at 4–5 clusters in the regenerating roots compared to the same in intact roots (Figure 5). This subtle distinction suggests a sharp limit in the correlation length among cell division events (i.e. small clusters of cell divisions) but also a propensity of regenerating roots to exhibit a higher number of foci of mitotic activity.
2.6. Density of mitotic event clusters is constant and analogous between regenerating and intact roots
To quantify the density of cell division clusters in both regenerating and uncut roots, we measured the mean pairwise distance between their centres of mass (Figure 6). Overall the two distributions were significantly different (Figure 6A; K-S test, p < 0.001), with a barely significant difference in the first few hours of regeneration (Figure 6B; K-S test, p = 0.001), and then disappeared (Figure 6C; K-S test, p = 0.001) only to become statistically very clear 24 hours after excision (Figure 6D and 6E; K-S test, p < 0.001).

Figure 6. Distributions of pairwise distances between cluster centres of mass. Histograms, experimental data lines and kernel density estimation of the experimental data (smooth fitting).
3. Discussion
We presented a quantitative characterization of the temporal and spatial distribution of cell divisions in intact and regenerating Arabidopsis root tips. Several biologically relevant observations can be extracted from the data.
First, the intermittent nature of the temporal sequence of mitotic events (Figure 1) indicates that mitotic events are not randomly distributed in time. In other words, the underlying dynamics of cell divisions in the tissue cannot be explained simply by perfectly uncoupled cells undergoing a noisy cell cycle. A significant body of work describes the complex genetic networks regulating cell–cell interactions during cell division and differentiation in Arabidopsis roots, so the fact that cell divisions are not simply independent random events is perhaps not surprising. An intermittent pattern can be described as a sequence of ‘bursts’ or periods of activity above an arbitrary threshold. The distributions of burst size (Figure 2) look very different than a Poisson distribution, confirming that these are not random, uncorrelated events. This might be expected given the short- and long-range cell–cell signalling, but it is an important quantitative visualzation. Our data also show that regenerating roots tend to produce slightly larger bursts, involving a larger number of cell divisions, compared to intact roots (Figure 2). This indicates that regeneration entails not only more cell divisions but also that these are compacted in discrete periods (bursts) of higher activity.
Second, cell division activity in both intact and regenerating roots shows a superposition of several periodicities, but regenerating roots are characterized by an underlying period of 16 hours, which is not detected in intact roots (Figure 3). Although the regenerating and intact groups are composed of random individuals taken from the same isogenic seed population and have been germinated and grown under identical conditions, we note that the seedlings are germinated under a regime of 16 hours in light and 8 hours in darkness. Immediately after root tip excision, the plants were grown and imaged under constant light. Is it possible that a memory of the 16-hour light cycle persists at the cellular level and that it is reflected in the cell division dynamics? If so, our data indicate that this should happen only during tissue regeneration, as no 16-hour periodicity was observed in intact roots. Future experiments carried out with different light/dark regimes might attempt to test this hypothesis.
Third, we found differences between intact and regenerating roots when considering the entire temporal distribution of single mitotic events or the frequency of single time frames containing a given number of cell divisions, despite the described intermittency. More specifically, while cell divisions in intact roots belong to a single mode centred at approximately 3–5 events at any given time point, during regeneration, a second mode of division emerges, centred at approximately 11 events at any given time-point (Figure 4). This becomes particularly evident 24 hours after root excision, suggesting that after this time-point, the regenerating tissue undergoes a transition towards a more complex regime of cell division dynamics. It can be difficult to obtain sufficient temporal and spatial statistics to identify the collective correlations associated with true phase transitions and criticality, so here we limit our reference to a developmental transition.
Fourth, the mitotic events appeared to be clustered in space, suggesting the existence of a short-range inducing signal to trigger cell division in neighbouring cells, coupled with a long-range inhibitory signal to separate clusters. Although it is beyond the scope of this work, we suggest that effective diffusion constants of the inducing and inhibiting signals could be estimated computationally with a model based on reaction–diffusion (Turing, Reference Turing1952). In addition, mechanical cell–cell interactions may also contribute to spatial and temporal patterns of mitotic activity, in analogy to what is suggested by computational models in animal systems (Carpenter et al., Reference Carpenter, Pérez-Verdugo and Banerjee2024; Shraiman, Reference Shraiman2005).
Finally, regenerating roots contain a slightly higher number of clusters per frame (Figure 5), which are also more densely distributed (i.e. with smaller inter-cluster distance) when compared to intact roots (Figure 6). This suggests that a similar tissue volume is going through cell proliferation in regenerating and intact roots but that subregions of high mitotic activity (clusters) appear more often in the regenerating tissue.
Overall, the presented data paint an original quantitative picture where the cell division dynamics in regenerating roots evolve faster than those in intact roots, possibly revealing a developmental transition approximately 24 hours after physical perturbation. Although this is only a first step towards a full quantitative characterization of tissue regeneration, we believe that the focus on cell divisions is important to capture the complex dynamics driving tissue self-organization. In future works, a persistent fluorescent reporter and increased spatial and temporal resolutions might be used to track individual cells in time during regeneration, as has been demonstrated in intact roots (Sena et al., Reference Sena, Frentz, Birnbaum and Leibler2011), to provide quantitative information beyond cell proliferation dynamics.
4. Methods
4.1. Plant material
Mitotic events were visualized using an existing Arabidopsis transgenic line expressing the cyclin-GFP fusion CYCB1;1::GFP (Reddy et al., Reference Reddy, Heisler, Ehrhardt and Meyerowitz2004). Arabidopsis seeds were sterilized, stratified and stored at 4°C before sowing on sterile room-temperature rectangular plates prepared in sterile conditions with solid media consisting of 0.175% w/v Murashige and Skoog Basal Medium (MS; Sigma–Aldrich, UK), 0.5% w/v sucrose (Sigma–Aldrich), 0.05% w/v MES hydrate (Sigma–Aldrich) and 0.8% w/v agar, adjusted to pH 5.7 (KOH), which was sterilized by autoclaving. The plates were placed in vertical racks in a plant growth chamber with 120 μmol/m2/s light intensity on a 16:8 hour light:dark cycle and constant 23°C.
4.2. Microdissection
Regenerating roots were manually excised using a 100 Sterican 27G needle tip (B Braun) under a Nikon SMZ1000 dissecting microscope, 180x magnification, following published procedures (Kral et al., Reference Kral, Ougolnikova and Sena2016; Sena et al., Reference Sena, Wang, Liu, Hofhuis and Birnbaum2009). Root excisions were performed around 100 μm, just above the quiescent centre in the apical meristem of primary roots.
4.3. Mounting
Five days post-germination, plants were moved and mounted in an imaging cuvette as previously described (Baesso et al., Reference Baesso, Randall and Sena2018). Briefly, roots were taken and placed on solid media plates with 5% w/v agar (all other reagents were the same as the germination plates).
Excised and control roots were then both mounted into the corner of an imaging cuvette by flowing liquid media (0.04% w/v MS, 0.5% w/v sucrose, 0.05% w/v MES) and using capillary action to pull the root down the length of the cuvette with the hypocotyl and cotyledons above the top of the cuvette. The root was held in place with a sterile, heat-shrink plastic-coated pin, which was in turn held in place by 2-mm glass beads for one-third of the volume of the cuvette, followed by 1-mm glass beads until 10 mm from the top of the cuvette. Liquid media was perfused into the chamber at ~1 mL/min through a custom cuvette top with a recessed corner for the cotyledons. A second cuvette with a glass coverslip top and two ~0.5 cm2 gas-exchange windows covered with gas-permeable sterile tape was placed over the top of the perfusion chamber, allowing for gas exchange and broad-spectrum incident light on the cotyledons. Media temperature was monitored using an infrared thermometer mounted on tubing before the perfusion chamber, and a multistage heating element was used to keep the media temperature at 23°C. All tubing, media, glass beads, pins, perfusion tops and imaging cuvettes were autoclaved before use. Plastic imaging chamber tops were submerged in a bleach solution for 30 s before multiple rinses with autoclaved sterile water.
4.4. Microscopy
Imaging of the apical meristem of Arabidopsis roots (Supplementary Figure S1) was performed using a previously described home-built light-sheet microscope (Baesso et al., Reference Baesso, Randall and Sena2018). The root was imaged through 60 planes every 15 minutes for up to 7 days. At each plane, 6 images were taken, and the plane of maximum focus was kept. Focusing was enhanced by automatically detecting the edge of the root at the beginning of every image set, moving the focus 30 μm into the root from that plane, automatically detecting the plane of maximum focus, and moving back 30 μm from that point, such that the starting point for the autofocusing step would be at maximum focus in the centre of the root. The root tip is automatically tracked using custom MATLAB code (Baesso et al., Reference Baesso, Randall and Sena2018), which in turn will move the cuvette stage in x, y, and z to keep the root in focus and centred in the field of view throughout the experiment. Cell division events were segmented and tracked across time frames using a previously described Python code (Amarteifio et al., Reference Amarteifio, Fallesen, Pruessner and Sena2021).
4.5. Statistical analysis
When comparing two samples of measurements, the non-parametric two-sample Kolmogorov–Smirnov (KS) test was used. Unless stated otherwise, all comparisons were performed assuming independence (unpaired test). All statistical tests were performed in Python using the scipy statistics package; step-by-step methods for statistical analysis and plots used in the figures are described in the Jupyter Notebooks stored at https://github.com/GiovanniSena/Fallesen_2024.
Data availability statement
The images generated in this study are available from the corresponding author upon request.
Author contributions
T.F. performed the experiments, analysed the data (including statistics) and wrote the article; S.A., G.P. and H.J.J. all contributed to data analysis; G.S. conceived and designed the project, supervised the experiments and contributed to the writing of the manuscript. All authors revised and approved the final manuscript.
Funding statement
This work was supported by the Biotechnology and Biological Sciences Research Council research grant BB/M002624/1.
Competing interest
The authors declare none.
Supplementary materials
The supplementary material for this article can be found at http://doi.org/10.1017/qpb.2024.7.







 Open access
Open access
Comments
London, 28/11/2023
Dear Prof. Hamant,
I am writing to submit our manuscript entitled “Intermittent Cell Division Dynamics in Regenerating Arabidopsis Roots Reveals Complex Long-range Interactions” for consideration in the Quantitative Imaging collection of Quantitative Plant Biology.
In this work, we present a comprehensive quantitative analysis of cell division dynamics in intact and regenerating root meristems of Arabidopsis thaliana. Leveraging a live imaging setup based on light-sheet fluorescence microscopy, previously developed in our laboratory, we achieved the necessary temporal resolution to monitor and analyse cell division events throughout the regeneration process. Our study provides novel insights into the temporal and spatial patterns of cell division, revealing a statistically significant difference in the frequency of mitotic events and the spatial distribution of mitotic event clusters between intact and regenerating roots. This is a step forward toward a quantitative characterisation of self-organisation in plant developmental processes.
We believe that our manuscript aligns well with the scope of Quantitative Plant Biology, particularly within the Quantitative Imaging collection, as it contributes valuable quantitative data, based on high-resolution 3D time-lapse imaging, to the understanding of plant tissue regeneration. The adoption of cutting-edge imaging techniques and the rigorous quantitative analysis presented in our study make it relevant to the journal’s readership interested in advancing the field of plant developmental biology through quantitative approaches.
We have contacted Prof. George Bessel and confirmed that he is available to act as Editor.
We hope that our findings will be of interest to the Quantitative Plant Biology community, and we look forward to the opportunity for our work to be reviewed for publication. We appreciate your time and consideration of our submission.
Sincerely,
Giovanni Sena