We use cookies to distinguish you from other users and to provide you with a better experience on our websites. Close this message to accept cookies or find out how to manage your cookie settings.
This journal utilises an Online Peer Review Service (OPRS) for submissions. By clicking "Continue" you will be taken to our partner site
https://mc.manuscriptcentral.com/qpb.
Please be aware that your Cambridge account is not valid for this OPRS and registration is required. We strongly advise you to read all "Author instructions" in the "Journal information" area prior to submitting.
To save this undefined to your undefined account, please select one or more formats and confirm that you agree to abide by our usage policies. If this is the first time you used this feature, you will be asked to authorise Cambridge Core to connect with your undefined account.
Find out more about saving content to .
To send this article to your Kindle, first ensure [email protected] is added to your Approved Personal Document E-mail List under your Personal Document Settings on the Manage Your Content and Devices page of your Amazon account. Then enter the ‘name’ part of your Kindle email address below. Find out more about sending to your Kindle.
Find out more about saving to your Kindle.
Note you can select to save to either the @free.kindle.com or @kindle.com variations. ‘@free.kindle.com’ emails are free but can only be saved to your device when it is connected to wi-fi. ‘@kindle.com’ emails can be delivered even when you are not connected to wi-fi, but note that service fees apply.
Studies on the mechanics of plant cells usually focus on understanding the effects of turgor pressure and properties of the cell wall (CW). While the functional roles of the underlying cytoskeleton have been studied, the extent to which it contributes to the mechanical properties of cells is not elucidated. Here, we study the contributions of the CW, microtubules (MTs) and actin filaments (AFs), in the mechanical properties of Nicotiana tabacum cells. We use a multiscale biomechanical assay comprised of atomic force microscopy and micro-indentation in solutions that (i) remove MTs and AFs and (ii) alter osmotic pressures in the cells. To compare measurements obtained by the two mechanical tests, we develop two generative statistical models to describe the cell’s behaviour using one or both datasets. Our results illustrate that MTs and AFs contribute significantly to cell stiffness and dissipated energy, while confirming the dominant role of turgor pressure.
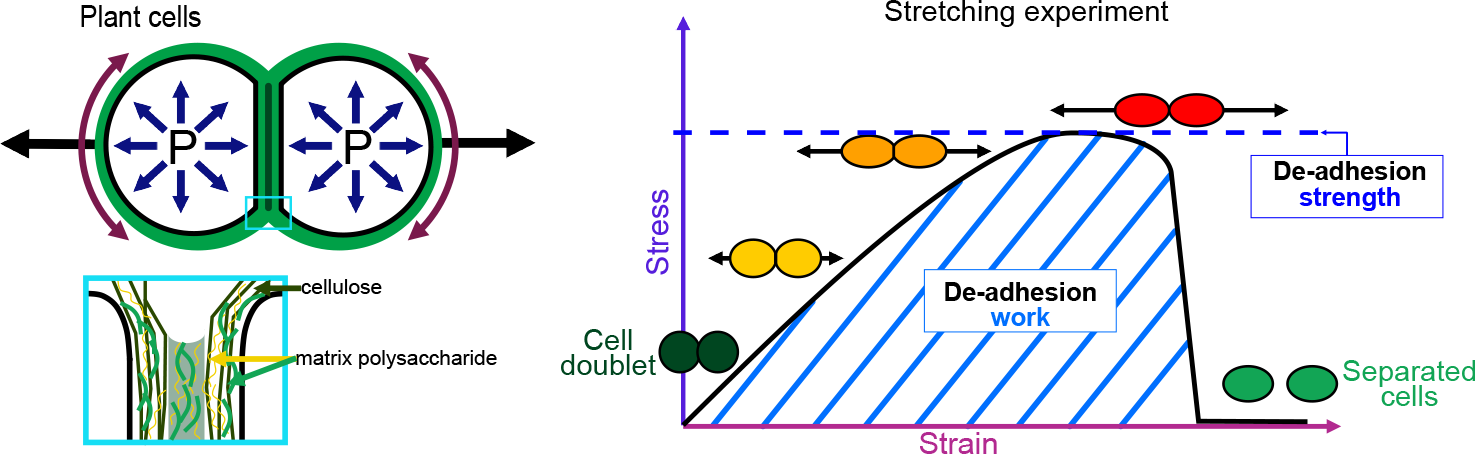
Cell–cell adhesion is a fundamental feature of multicellular organisms. To ensure multicellular integrity, adhesion needs to be tightly controlled and maintained. In plants, cell–cell adhesion remains poorly understood. Here, we argue that to be able to understand how cell–cell adhesion works in plants, we need to understand and quantitatively measure the mechanics behind it. We first introduce cell–cell adhesion in the context of multicellularity, briefly explain the notions of adhesion strength, work and energy and present the current knowledge concerning the mechanisms of cell–cell adhesion in plants. Because still relatively little is known in plants, we then turn to animals, but also algae, bacteria, yeast and fungi, and examine how adhesion works and how it can be quantitatively measured in these systems. From this, we explore how the mechanics of cell adhesion could be quantitatively characterised in plants, opening future perspectives for understanding plant multicellularity.
Plant shoot gravitropism is a complex phenomenon resulting from gravity sensing, curvature sensing (proprioception), the ability to uphold self-weight and growth. Although recent data analysis and modelling have revealed the detailed morphology of shoot bending, the relative contribution of bending force (derived from the gravi-proprioceptive response) and stretching force (derived from shoot axial growth) behind gravitropism remains poorly understood. To address this gap, we combined morphological data with a theoretical model to analyze shoot bending in wild-type and lazy1-like 1 mutant Arabidopsis thaliana. Using data from actual bending events, we searched for model parameters that minimized discrepancies between the data and mathematical model. The resulting model suggests that both the bending force and the stretching force differ significantly between the wild type and mutant. We discuss the implications of the mechanical forces associated with differential cell growth and present a plausible mechanical explanation of shoot gravitropism.
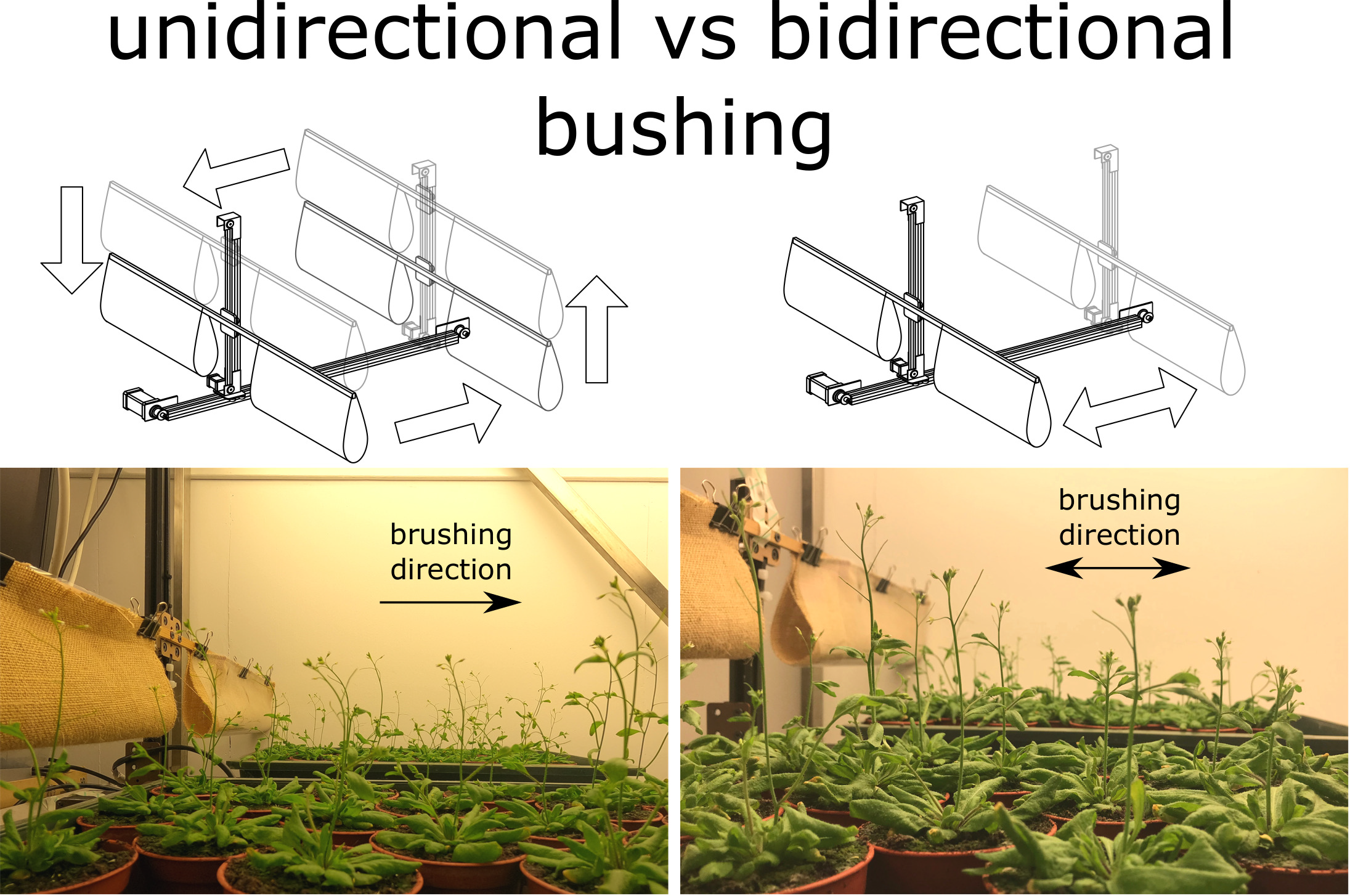
Plants acclimate to various types of mechanical stresses through thigmomorphogenesis and alterations in their mechanical properties. Although resemblance between wind- and touch-induced responses provides the foundation for studies where wind influence was mimicked by mechanical perturbations, factorial experiments revealed that it is not always straightforward to extrapolate results induced by one type of perturbation to the other. To investigate whether wind-induced changes in morphological and biomechanical traits can be reproduced, we subjected Arabidopsis thaliana to two vectorial brushing treatments. Both treatments significantly affected the length, mechanical properties and anatomical tissue composition of the primary inflorescence stem. While some of the morphological changes were found to be in line with those induced by wind, changes in the mechanical properties exhibited opposite trends irrespective of the brushing direction. Overall, a careful design of the brushing treatment gives the possibility to obtain a closer match to wind-induced changes, including a positive tropic response.
Biomechanical properties of the cell wall (CW) are important for many developmental and adaptive responses in plants. Expansins were shown to mediate pH-dependent CW enlargement via a process called CW loosening. Here, we provide a brief overview of expansin occurrence in plant and non-plant species, their structure and mode of action including the role of hormone-regulated CW acidification in the control of expansin activity. We depict the historical as well as recent CW models, discuss the role of expansins in the CW biomechanics and address the developmental importance of expansin-regulated CW loosening in cell elongation and new primordia formation. We summarise the data published so far on the role of expansins in the abiotic stress response as well as the rather scarce evidence and hypotheses on the possible mechanisms underlying expansin-mediated abiotic stress resistance. Finally, we wrap it up by highlighting possible future directions in expansin research.
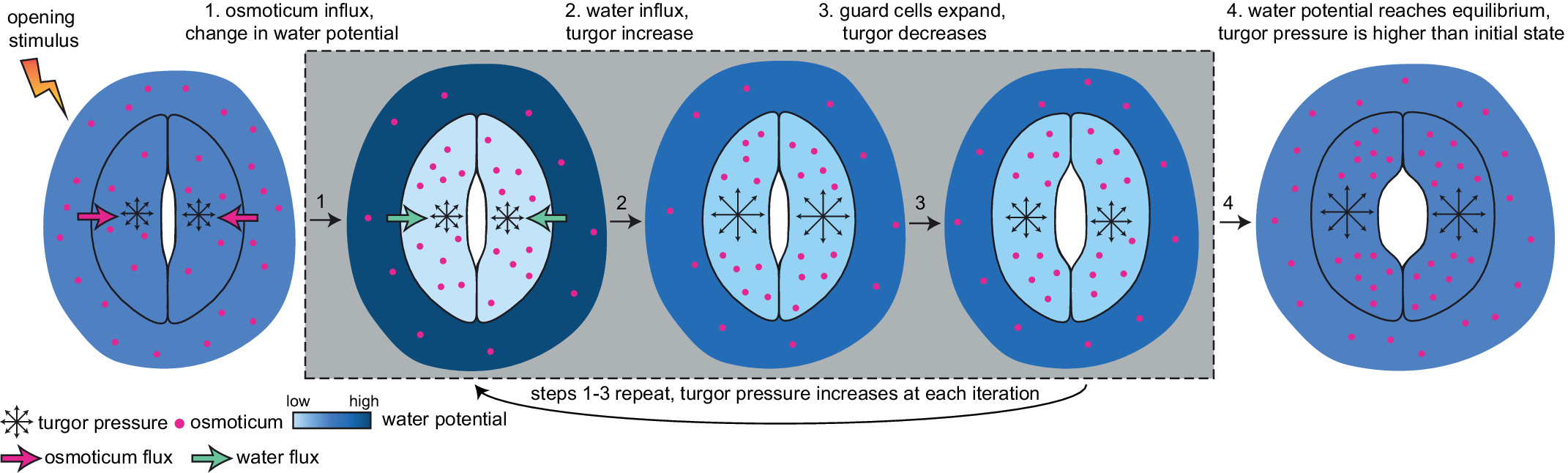
The ability of plants to absorb CO2 for photosynthesis and transport water from root to shoot depends on the reversible swelling of guard cells that open stomatal pores in the epidermis. Despite decades of experimental and theoretical work, the biomechanical drivers of stomatal opening and closure are still not clearly defined. We combined mechanical principles with a growing body of knowledge concerning water flux across the plant cell membrane and the biomechanical properties of plant cell walls to quantitatively test the long-standing hypothesis that increasing turgor pressure resulting from water uptake drives guard cell expansion during stomatal opening. To test the alternative hypothesis that water influx is the main motive force underlying guard cell expansion, we developed a system dynamics model accounting for water influx. This approach connects stomatal kinetics to whole plant physiology by including values for water flux arising from water status in the plant .