


Periodontal disease is a chronic inflammatory disease characterised by the destruction of connective tissue and alveolar bone and, in severe cases, culminates in the loss of teeth( Reference Williams 1 ). There is evidence suggesting that periodontal disease acts as a potential risk factor for several systemic diseases( Reference Teng, Taylor and Scannapieco 2 , Reference Seymour, Ford and Cullinan 3 ).
It is clear that periodontal tissue destruction is caused as a consequence of the host response to the periodontopathic bacteria. Thus, it is expected that the modulation of host-mediated tissue destruction might be a treatment strategy, in addition to traditional periodontal treatment, for periodontal disease.
Lipopolysaccharide (LPS) is a major component of the outer membrane of Gram-negative bacteria. LPS of periodontal pathogens induce inflammatory cascades, including a variety of proinflammatory mediators such as NO and cytokines produced by different cell types, and thereby promote the host inflammatory reactions associated with periodontal disease( Reference Sosroseno, Barid and Herminajeng 4 – Reference Kim, Choi and Kim 7 ). Hence, host modulatory substances regulating NO and specific cytokines may be useful for the prevention and treatment of periodontal disease.
n-3 PUFA, including DHA, are essential fatty acids that have been shown to have potent anti-inflammatory actions in a variety of chronic inflammatory diseases( Reference Calder 8 – Reference Geusens, Wouters and Nijs 10 ). There have been several studies in animal models and human subjects suggesting that dietary supplementation with n-3 PUFA could be used to modulate periodontal disease progression through its anti-inflammatory effects( Reference Kesavalu, Vasudevan and Raghu 11 – Reference Iwasaki, Yoshihara and Moynihan 13 ). DHA is a principal n-3 PUFA present in fish oil. Several reports have indicated that dietary intake of DHA is associated with lower prevalence of periodontitis in adults( Reference Naqvi, Buettner and Phillips 12 , Reference Iwasaki, Yoshihara and Moynihan 13 ). However, to our knowledge, data are rather limited with regard to the underlying mechanisms of action of DHA in periodontal disease progression. In the present study, we investigated the effect of DHA on the production of proinflammatory mediators in murine macrophages stimulated with LPS isolated from Prevotella intermedia, a pathogen implicated in inflammatory periodontal disease, and its mechanisms of action. The present study may serve to explain at a cellular level the beneficial effect of DHA on periodontal disease described in animal and human studies.
Methods
Reagents
DHA was purchased from Sigma-Aldrich. Primary antibodies against inducible NO synthase (iNOS), c-Jun N-terminal kinase (JNK), phospho-JNK, p38, phospho-p38, inhibitory κB (IκB)-α, signal transducer and activator of transcription (STAT)1 and phospho-STAT1 were acquired from Cell Signaling Technology. Antibodies against haeme oxygenase 1 (HO-1) NF-κB p65, NF-κB p50, β-actin and poly (ADP-ribose) polymerase-1 (PARP-1) were the products of Santa Cruz Biotechnology. Tin protoporphyrin IX (SnPP) was obtained from Frontier Scientific, Inc. All other chemicals and reagents used were purchased from Sigma-Aldrich, unless otherwise stated.
Bacteria, culture conditions and lipopolysaccharide isolation
P. intermedia ATCC 25 611 cells were grown in GAM broth (Nissui) supplemented with 1 μg/ml menadione and 5 μg/ml haemin under anaerobic conditions as described in our previous work( Reference Kim, Ha and Choi 5 ). LPS was isolated from lyophilised P. intermedia cells using the standard hot-phenol–water protocol as described previously( Reference Kim, Ha and Choi 5 ). Nucleic acids were eliminated by treating the LPS suspension with deoxyribonuclease and ribonuclease. Contaminating proteins were hydrolysed with proteinase K.
Cell culture and determination of cytotoxicity
RAW264.7 cells (American Type Culture Collection) were maintained in Dulbecco's modified Eagle's medium supplemented with 10 % heat-inactivated fetal bovine serum, penicillin (100 U/ml) and streptomycin (100 μg/ml) under a humidified atmosphere of 5 % CO2–95 % air at 37°C. The cells were plated at a density of 5 × 105 cells/well in twenty-four-well culture plates and incubated for at least 12 h to allow them to adhere to the plates. The cells were then pretreated with different concentrations of DHA for 2 h and incubated with P. intermedia LPS for the indicated periods of time, after which culture supernatants were harvested and assayed for NO, IL-1β and IL-6. To determine the cytotoxicity of DHA, 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay was carried out according to the manufacturer's instructions.
Measurement of nitric oxide concentrations
NO synthesis was determined by an assay of the culture supernatant for nitrite (
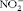 $$NO _{2}^{ - } $$
), a stable oxidative metabolite of NO(
Reference Green, Wagner and Glogowski
14
). Briefly, the cells were pretreated with the indicated doses of DHA for 2 h and incubated with P. intermedia LPS for 24 h, and 100 μl of Griess reagent were allowed to react with equal volumes of culture supernatants and left at room temperature for 10 min. Optical densities were measured at 540 nm with SpectraMax 250 ELISA Reader (Molecular Devices), and the concentrations of nitrite were determined using a standard curve prepared with serial dilutions of NaNO2.
$$NO _{2}^{ - } $$
), a stable oxidative metabolite of NO(
Reference Green, Wagner and Glogowski
14
). Briefly, the cells were pretreated with the indicated doses of DHA for 2 h and incubated with P. intermedia LPS for 24 h, and 100 μl of Griess reagent were allowed to react with equal volumes of culture supernatants and left at room temperature for 10 min. Optical densities were measured at 540 nm with SpectraMax 250 ELISA Reader (Molecular Devices), and the concentrations of nitrite were determined using a standard curve prepared with serial dilutions of NaNO2.
Measurement of IL-1β and IL-6 concentrations
The cells were pretreated with the indicated doses of DHA for 2 h and incubated with P. intermedia LPS for 24 h (for IL-6) or 48 h (for IL-1β), and the concentrations of IL-1β and IL-6 released into the culture medium were determined by ELISA using a commercially available kit (OptEIA; BD Pharmingen) according to the manufacturer's protocols.
Real-time PCR analysis
The cells were plated in 100 mm tissue-culture dishes at a density of 1 × 107 cells/dish and pretreated with the indicated doses of DHA for 2 h, followed by incubation with P. intermedia LPS for the indicated periods of time. The extraction of total RNA was carried out using the RNeasy Mini Kit (Qiagen) according to the manufacturer's protocol, and complementary DNA was synthesised from 1 μg of the total RNA using the iScript cDNA Synthesis Kit (Bio-Rad). A real-time PCR was carried out using the CFX96 Real-Time PCR Detection System (Bio-Rad) with specific primers for mouse iNOS, IL-1β, IL-6 and HO-1. β-Actin primer was used as an endogenous control. PCR was conducted with SsoFast EvaGreen Supermix (Bio-Rad) according to the manufacturer's directions. The following thermal cycler conditions were applied: after denaturation at 98°C for 30 s, PCR was carried out for a total of forty-five cycles, each of which consisted of denaturation at 95°C for 1 s and combined annealing/extension at 60°C for 5 s. The primers used were as follows: iNOS (130 bp) – sense 5′-GCACCACCCTCCTCGTTCAG-3′ and antisense 5′-TCCACAACTCGCTCCAAGATTCC-3′; IL-1β (131 bp) – sense 5′-TTCAGGCAGGCAGTATCA-3′ and antisense 5′-AGGATGGGCTCTTCTTCAA-3′; IL-6 (162 bp) – sense 5′-GCCAGAGTCCTTCAGAGAGATACAG-3′ and antisense 5′-GAATTGGATGGTCTTGGTCCTTAGC-3′; HO-1 (149 bp) – sense 5′-CAATGTGGCCTTCTCTCTGT-3′ and antisense 5′-TTTTGGTGAGGGAACTGTGT-3′; β-actin (149 bp) – sense 5′-TGAGAGGGAAATCGTGCGTGAC-3′ and antisense 5′-GCTCGTTGCCAATAGTGATGACC-3′. Each assay was normalised to β-actin mRNA.
Immunoblot analysis
The cells were plated in 60 mm tissue-culture dishes at 4 × 106 cells/dish and pretreated with the indicated doses of DHA for 2 h, followed by incubation with P. intermedia LPS for the indicated periods of time. Whole-cell lysates and nuclear fractions were prepared and analysed as described in our previous work( Reference Choi, Jin and Lee 15 ). Briefly, cell lysates were prepared using lysis buffer (50 mm-Tris·Cl, 150 mm-NaCl, 0·002 % sodium azide, 0·1 % SDS and 1 % Nonidet P-40), including protease inhibitors (1 mm-phenylmethanesulfonyl fluoride, 5 mg/ml aprotinin, 5 mg/ml pepstatin A and 5 mg/ml leupeptin). The nuclear fractions were prepared from the cells using the Active Motif Nuclear Extract Kit (Active Motif). Proteins (30 μg) were separated by SDS–PAGE, electrotransferred onto nitrocellulose membranes and then probed with the specific primary antibodies. After incubation with horseradish peroxidase-conjugated secondary antibodies, the immunoreactive bands were visualised on an X-ray film using enhanced chemiluminescence (Cell Signaling Technology). The intensity of each protein-specific band was quantified using a densitometer with densitometric software.
Assay of NF-κB DNA-binding activity
RAW264.7 cells were placed in 60 mm tissue-culture dishes, at a density of 4 × 106 cells per dish, and pretreated with the indicated doses of DHA for 2 h, followed by incubation with P. intermedia LPS for the indicated periods of time. After extraction of the nuclear protein as described above, the DNA-binding activity of NF-κB p65 or p50 was assayed using the TransAM NF-κB p65/NF-κB p50 Transcription Factor Assay Kit (Active Motif) as described previously( Reference Choi, Jin and Lee 15 ).
Statistical analysis
Data are expressed as means and standard deviations, and statistical analysis was carried out using Student's t test with P< 0·05 being considered statistically significant.
Results
DHA suppresses Prevotella intermedia lipopolysaccharide-induced production of nitric oxide, IL-1β and IL-6
The stimulation of cells with P. intermedia LPS led to significant increases in the production of NO, IL-1β and IL-6 compared with the control (Fig. 1). However, pretreatment of cells with DHA significantly attenuated the LPS-induced production of these proinflammatory mediators in a dose-dependent fashion (Fig. 1). At the highest concentration of 100 μm, DHA suppressed the release of NO, IL-1β and IL-6 approximately by 86, 61 and 81 %, respectively. At the concentrations tested, DHA did not exhibit remarkable cytotoxicity as assessed by the MTT assay (Fig. 1(d)), suggesting that the inhibitory effects of DHA on the production of these proinflammatory mediators are not ascribable to its non-specific cytotoxic effects.

Fig. 1 Effects of DHA on Prevotella
intermedia lipopolysaccharide (LPS)-induced production of NO2
−, IL-1β and IL-6 and on viability in RAW264.7 cells. The cells were pretreated with the indicated doses of DHA for 2 h and incubated with P. intermedia LPS for 24 h (for NO2
− and IL-6) or 48 h (for IL-1β). The concentrations of (a) NO2
−, (b) IL-1β and (c) IL-6 released into the culture medium were determined by ELISA. Values are means of three independent experiments, with standard deviations represented by vertical bars. (d) Cell viability was determined using the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide assay. Values are means of three independent experiments, with standard deviations represented by vertical bars. ![]() , 24 h culture; ■, 48 h culture. Mean values were significantly different from those obtained when treated with P. intermedia LPS alone: * P< 0·05; ** P< 0·01.
, 24 h culture; ■, 48 h culture. Mean values were significantly different from those obtained when treated with P. intermedia LPS alone: * P< 0·05; ** P< 0·01.
By carrying out an immunoblot analysis, we also found that DHA significantly inhibited the expression of iNOS protein in a concentration-dependent manner in P. intermedia LPS-stimulated cells (Fig. 2). Further study using real-time PCR analysis clearly showed that DHA also potently suppressed the LPS-induced expression of iNOS, IL-1β and IL-6 mRNA, parallel to the effects on the production of these proinflammatory mediators (Fig. 3).

Fig. 2 Effect of DHA on Prevotella intermedia lipopolysaccharide (LPS)-induced inducible NO synthase (iNOS) protein expression in RAW264.7 cells. The cells were pretreated with the indicated doses of DHA for 2 h and incubated with P. intermedia LPS for 24 h. The synthesis of iNOS protein was measured by an immunoblot analysis of cell lysates using iNOS-specific antibody. The expression of iNOS protein was quantified by densitometric scanning. A representative immunoblot from three separate experiments with similar results is shown. Values are means, with standard deviations represented by vertical bars. Mean values were significantly different from that obtained when treated with P. intermedia LPS alone: * P< 0·05; ** P< 0·01.

Fig. 3 Effects of DHA on Prevotella intermedia lipopolysaccharide (LPS)-induced (a) inducible NO synthase (iNOS), (b) IL-1β and (c) IL-6 mRNA expression in RAW264.7 cells. The cells were pretreated with the indicated doses of DHA for 2 h, followed by incubation with P. intermedia LPS for 6 h (for IL-1β) or 24 h (for iNOS and IL-6). Real-time PCR was carried out using EvaGreen Supermix (Bio-Rad), with β-actin being used as an endogenous control. Values are means of three independent experiments, with standard deviations represented by vertical bars. Mean values were significantly different from those obtained when treated with P. intermedia LPS alone: * P< 0·05; ** P< 0·01.
DHA-induced haeme oxygenase-1 plays a role in the inhibition of Prevotella intermedia lipopolysaccharide-induced nitric oxide production
We examined whether the suppressive effects of DHA on P. intermedia LPS-induced production of NO, IL-1β and IL-6 are mediated through the expression of HO-1. Immunoblot analysis indicated that DHA dose dependently up-regulated the expression of HO-1 protein in cells treated with P. intermedia LPS (Fig. 4(a)). Real-time PCR analysis indicated that DHA also induced a dose-dependent increase in the levels of HO-1 mRNA (Fig. 4(b)).

Fig. 4 Role of haeme oxygenase-1 (HO-1) in the inhibitory effects of DHA on Prevotella intermedia lipopolysaccharide (LPS)-induced NO production in RAW264.7 cells. (a, b) The cells were pretreated with the indicated doses of DHA for 2 h, followed by incubation with P. intermedia LPS for 6 h (for HO-1 protein) or 3 h (for HO-1 mRNA). (a) The synthesis of HO-1 protein was measured by an immunoblot analysis of cell lysates using HO-1-specific antibody. The expression of HO-1 protein was quantified by densitometric scanning. A representative immunoblot from three separate experiments with similar results is shown. * Mean value was significantly different from that obtained when treated with P. intermedia LPS alone (P< 0·05). (b) Real-time PCR was carried out using EvaGreen Supermix (Bio-Rad), with β-actin being used as an endogenous control. Values are means of three independent experiments, with standard deviations represented by vertical bars. Mean values were significantly different from those obtained when treated with P. intermedia LPS alone: * P< 0·05; ** P< 0·01. (c) The cells were pretreated with DHA (100 μm) for 2 h and incubated with P. intermedia LPS (10 μg/ml) for 24 h in the presence of different doses of tin protoporphyrin IX (SnPP). The production of NO was assayed by measuring the accumulation of nitrite in culture supernatants. Values are means of three independent experiments, with standard deviations represented by vertical bars. ** Mean value was significantly different from that obtained when treated with P. intermedia LPS alone (P< 0·01). †† Mean values were significantly different from those obtained when treated with P. intermedia LPS plus DHA (P< 0·01).
To investigate the possibility that the expression of HO-1 induced by DHA is responsible for the inhibition of P. intermedia LPS-induced production of NO, IL-1β and IL-6, we treated the cells with DHA (100 μm) and P. intermedia LPS (10 μg/ml) in the presence of different doses of SnPP, a selective inhibitor of HO-1 activity, and measured the concentrations of these proinflammatory mediators. SnPP treatment significantly mitigated the inhibitory effects of DHA on LPS-induced NO production concentration dependently (Fig. 4(c)), whereas it had no effect on the DHA-mediated inhibition of IL-1β and IL-6 production (data not shown).
DHA prevents Prevotella intermedia lipopolysaccharide from activating the c-Jun N-terminal kinase pathway
We have previously shown that the JNK, p38, NF-κB and Janus kinase (JAK) 2/STAT1 pathways are involved in the production of NO and IL-6 induced by P. intermedia LPS in RAW264.7 cells( Reference Choi, Jin and Lee 15 ). These pathways are also involved in P. intermedia LPS-induced production of IL-1β (data not shown). We first investigated the action of DHA on P. intermedia LPS-induced activation of JNK and p38. DHA treatment significantly attenuated the phosphorylation of JNK induced by LPS, whereas it had little effect on the phosphorylation of p38 (Fig. 5).

Fig. 5 Effects of DHA on Prevotella intermedia lipopolysaccharide (LPS)-induced phosphorylation of (a) c-Jun N-terminal kinase (JNK) and (b) p38 in RAW264.7 cells. The cells were pretreated with the indicated doses of DHA for 2 h, followed by incubation with P. intermedia LPS (10 μg/ml) for 30 min (for JNK) or 15 min (for p38). Cells lysates were subjected to an immunoblot analysis using specific antibodies. The phosphorylation of JNK and p38 was quantified by densitometric scanning. A representative immunoblot from three separate experiments with similar results is shown. Mean values were significantly different from those obtained when treated with P. intermedia LPS alone: * P< 0·05; ** P< 0·01.
DHA suppresses the transcriptional activity of NF-κB by regulating the nuclear translocation and DNA-binding activity of NF-κB p50 subunit
We investigated the possibility that DHA exerts its effects on P. intermedia LPS-induced production of NO, IL-1β and IL-6 by regulating the NF-κB signalling pathway. We carried out an immunoblot analysis of cell lysates to determine whether DHA is able to inhibit the degradation of IκB-α induced by P. intermedia LPS. While the incubation of cells with P. intermedia LPS induced the degradation of IκB-α, DHA failed to prevent LPS from degrading IκB-α (Fig. 6(a)). We then carried out an immunoblot analysis of nuclear fractions to examine whether DHA prevents the nuclear translocation of NF-κB subunits. The nuclear translocation of NF-κB p65 and p50 subunits was markedly increased upon exposure of cells to P. intermedia LPS, while only the nuclear translocation of NF-κB p50 subunit was significantly reduced by DHA (Fig. 6(b)). Finally, we determined whether DHA could affect the DNA-binding activities of NF-κB subunits by using the ELISA-based NF-κB transcription factor assay kit. P. intermedia LPS treatment strongly promoted the binding of NF-κB p65 and p50 to DNA (Fig. 6(c)). Whereas DHA treatment mitigated LPS-induced NF-κB p50 binding activity dose dependently, it had a weaker effect on NF-κB p65 binding activity (Fig. 6(c)).

Fig. 6 Effects of DHA on Prevotella
intermedia lipopolysaccharide (LPS)-induced activation of NF-κB in RAW264.7 cells. (a–c) The cells were pretreated with the indicated doses of DHA for 2 h and incubated with P. intermedia LPS (10 μg/ml). (a) After 30 min of incubation, the degradation of inhibitory κB-α (IκB-α) was determined by an immunoblot analysis of cell lysates using antibody against IκB-α. The expression of IκB-α was quantified by densitometric scanning. A representative immunoblot from three separate experiments with similar results is shown. (b, c) After 30 min (for NF-κB p65) or 8 h (for NF-κB p50) of incubation, the nuclear fraction was isolated from the cells. (b) The nuclear translocation of NF-κB subunits was assessed by an immunoblot analysis using antibodies against NF-κB p65 and p50. The expression of p65 ■ and p50 ![]() was quantified by densitometric scanning. A representative immunoblot from three separate experiments with similar results is shown. (c) The DNA-binding activity of NF-κB in nuclear extracts was assessed using the ELISA-based NF-κB transcription factor assay kit. Values are means of two independent experiments, with standard deviations represented by vertical bars. Mean values were significantly different from those obtained when treated with P. intermedia LPS alone: * P< 0·05; ** P< 0·01. OD, optical density.
was quantified by densitometric scanning. A representative immunoblot from three separate experiments with similar results is shown. (c) The DNA-binding activity of NF-κB in nuclear extracts was assessed using the ELISA-based NF-κB transcription factor assay kit. Values are means of two independent experiments, with standard deviations represented by vertical bars. Mean values were significantly different from those obtained when treated with P. intermedia LPS alone: * P< 0·05; ** P< 0·01. OD, optical density.
DHA attenuates Prevotella intermedia lipopolysaccharide-induced phosphorylation of signal transducer and activator of transcription 1
We examined the possibility that DHA exerts its effects on P. intermedia LPS-induced production of NO, IL-1β and IL-6 through the inhibition of the STAT1 signalling pathway. Whole-cell lysates were prepared and immunoblot analysis was carried out. DHA significantly inhibited P. intermedia LPS-induced STAT1 phosphorylation (Fig. 7).

Fig. 7 Effects of DHA on Prevotella intermedia lipopolysaccharide (LPS)-induced activation of signal transducer and activator of transcription 1 (STAT1) in RAW264.7 cells. The cells were pretreated with the indicated doses of DHA for 2 h and incubated with P. intermedia LPS (10 μg/ml). After 4 h of incubation, the expression of phospho-STAT1 was measured by an immunoblot analysis of cell lysates. Total STAT1 was used as an internal control. The phosphorylation of STAT1 was quantified by densitometric scanning. A representative immunoblot from three separate experiments with similar results is shown. ** Mean values were significantly different from that obtained when treated with P. intermedia LPS alone (P< 0·01).
Discussion
The excessive production of proinflammatory mediators by immune cells in response to periodontal pathogens is thought to have an important role in the development of periodontal disease. In the present study, we sought to determine whether DHA could regulate the production of proinflammatory mediators, such as NO, IL-1β and IL-6, by murine macrophages stimulated with LPS isolated from P. intermedia and attempted to determine the possible mechanisms of action.
P. intermedia is a key periodontal pathogen that is prevalent in the periodontal pockets of patients with chronic periodontitis( Reference Slots, Bragd and Wikström 16 , Reference Tanner, Haffer and Bratthall 17 ). This bacterium is also strongly associated with necrotising ulcerative gingivitis( Reference Chung, Nisengard and Slots 18 ) and pregnancy gingivitis( Reference Kornman and Loesche 19 ). We used LPS isolated from P. intermedia as a bacterial inflammatory stimulus in the present study. The findings of our previous reports clearly indicate that P. intermedia LPS is one of the most potent inducers of inflammatory mediators, including NO, TNF-α and IL-6, in murine and human macrophages( Reference Kim, Ha and Choi 5 , Reference Kim, Choi and Kim 7 , Reference Choi, Jin and Lee 15 ). In addition, the LPS of this bacterium has been shown to induce the release of NO and IL-6 from fetal mouse osteoblasts in organoid culture and dose dependently inhibit bone formation by reducing alkaline phosphatase activity and Ca incorporation( Reference Pelt, Zimmermann and Ulbrich 20 ).
The LPS of bacterial species such as Porphyromonas gingivalis might be more relevant to the pathogenesis of periodontal disease. However, there are strain-dependent variations in the ability of P. gingivalis LPS to induce the production of inflammatory mediators such as NO. For example, LPS isolated from P. gingivalis ATCC 33 277 failed to induce the production of NO by RAW264.7 cells, similar to that isolated from P. gingivalis W50( Reference Sosroseno, Herminajeng and Bird 21 , Reference Shapira, Champagne and Van Dyke 22 ). In contrast, the findings reported by Shapira et al. ( Reference Shapira, Champagne and Van Dyke 22 ) and our group (S-J Kim, IS Choi and E-Y Choi, unpublished results) indicate that NO could be produced by murine macrophages when stimulated with LPS preparations of two strains of P. gingivalis, A7436 and 381, although the induction of NO by these LPS is far less potent than that by LPS isolated from enteric bacteria. Therefore, the present study attempted to evaluate the ability of DHA to suppress the production of proinflammatory mediators in cells stimulated with P. intermedia LPS.
Macrophages, which are present in higher numbers in active periodontal lesions than in inactive sites, play a central role in the host inflammatory response to pathogenic bacteria( Reference Zappa, Reinking-Zappa and Graf 23 , Reference Kornman, Page and Tonetti 24 ). Macrophages are main targets for the action of LPS, and P. intermedia LPS is likely to interact with infiltrating macrophages to produce and release proinflammatory mediators. Hence, we used the well-characterised RAW264.7 murine macrophage cell line to investigate the capacity of DHA to suppress the LPS-induced production of inflammatory mediators. However, it is possible that the responsiveness to DHA and LPS might be species specific. Work is in progress to test the effects of DHA on inflammation induced by P. intermedia LPS in human cells.
In the present study, the P. intermedia LPS preparation was treated with deoxyribonuclease, ribonuclease and proteinase K to minimise the effects of contaminating nucleic acids and proteins on macrophage responses. The protein content of the purified LPS was less than 0·1 %, and Coomassie Blue staining of overloaded SDS–polyacrylamide gels did not reveal any visible protein bands in the purified LPS, confirming the purity of the preparation (data not shown).
In the present study, we used LPS prepared from P. intermedia instead of non-oral bacterial LPS. The chemical and immunomodulating properties of P. intermedia LPS are quite different from those of LPS preparations of Enterobacteriaceae, such as Escherichia coli and Salmonella species( Reference Hamada, Takada and Ogawa 25 ). In addition, it has been reported that P. intermedia LPS activates the macrophages of classical LPS-non-responsive C3H/HeJ mice( Reference Kirikae, Nitta and Kirikae 26 ). Furthermore, in contrast to LPS isolated from Enterobacteriaceae, that isolated from P. intermedia was not susceptible to polymyxin B treatment( Reference Kirikae, Nitta and Kirikae 26 ).
A variety of inflammatory mediators, such as NO, IL-1β and IL-6, are thought to play an important role in the pathogenesis of inflammatory periodontal disease. Clinically, these proinflammatory mediators have been found at higher concentrations in sites affected by periodontal disease than in healthy sites and to be closely related to the clinical measures of disease severity( Reference Matejka, Partyka and Ulm 27 – Reference Geivelis, Turner and Pederson 31 ). In addition, studies have shown that NO, IL-1β and IL-6 have strong potential to induce bone resorption in periodontal disease( Reference Pelt, Zimmermann and Ulbrich 20 , Reference Tatakis 32 , Reference Liu, Kirschenbaum and Yao 33 ). The blockade of the production of these proinflammatory mediators, therefore, could be a highly efficient tool for blocking the development and progression of periodontal disease.
n-3 PUFA refer to a group of essential fatty acids including DHA, EPA and α-linolenic acid. We found that DHA is more effective than other n-3 PUFA at blocking P. intermedia LPS-induced production of NO, IL-1β and IL-6 in RAW264.7 cells (data not shown). For this reason, we decided to study the effects and underlying mechanisms of action of DHA on the production of proinflammatory mediators.
The production of proinflammatory mediators induced by P. intermedia LPS in RAW264.7 cells reached a maximum at a concentration of 10 μg/ml (data not shown). Hence, we used 10 μg/ml P. intermedia LPS as an inflammatory stimulus in the present study. The results of the present study indicate that DHA significantly attenuated the production of NO, IL-1β and IL-6 in RAW264.7 cells activated by P. intermedia LPS. DHA also potently suppressed the LPS-induced expression of iNOS, IL-1β and IL-6 mRNA, suggesting that it exhibits inhibitory effects on the production of these proinflammatory mediators at gene transcription and translation levels.
HO-1, the inducible isoform of haeme oxygenase that catalyses the degradation of haeme to bilirubin/biliverdin, free Fe and CO, has a number of cytoprotective functions including anti-inflammatory and antioxidant effects( Reference Ryter, Alam and Choi 34 – Reference Morse and Choi 36 ). It has been shown that HO-1-deficient mice exhibit increased inflammatory state and that the overexpression of HO-1 exerts anti-inflammatory effects( Reference Otterbein, Soares and Yamashita 35 , Reference Poss and Tonegawa 37 ). In the present study, we tested the possibility that the expression of HO-1 induced by DHA is responsible for the inhibition of P. intermedia LPS-induced production of NO, IL-1β and IL-6. The results indicated that DHA has an ability to induce the expression of HO-1 in cells treated with P. intermedia LPS and DHA-induced HO-1 plays a role in the inhibition of LPS-induced production of NO. However, HO-1 was not involved in the inhibition of P. intermedia LPS-induced IL-1β and IL-6 release by DHA.
Accumulated evidence has indicated that the anti-inflammatory actions of HO-1 are due to the enzymatic by-products of haeme degradation. These include CO, which has been shown to decrease the production of proinflammatory mediators in LPS-stimulated macrophages, and bilirubin, which is a potent antioxidant( Reference Otterbein, Bach and Alam 38 – Reference Sarady-Andrews, Liu and Gallo 40 ). Therefore, we suggest that the induction of HO-1 by DHA may lead to the inhibition of P. intermedia LPS-induced NO production through the enhanced accumulation of CO and bilirubin arising from haeme degradation.
Mitogen-activated protein kinase signal transduction pathways are known to play critical roles in the LPS-induced activation of macrophages and resultant production of proinflammatory mediators. DHA prevented P. intermedia LPS from activating the JNK pathway, suggesting that this pathways is involved in the inhibition of P. intermedia LPS-induced production of NO, IL-1β and IL-6 by DHA.
NF-κB is one of the most important transcription factors known to play a key role in the LPS-induced expression of various inflammatory cytokines and enzymes in macrophages( Reference Baeuerle and Henkel 41 – Reference Karin and Ben-Neriah 43 ). In unstimulated cells, NF-κB remains inactive in the cytoplasm, where it forms complexes with the IκB proteins. Upon stimulation with a variety of stimuli, including LPS, IκB becomes phosphorylated, ubiquitinated and degraded by proteasomes. The resulting free NF-κB is then translocated into the nucleus, where it binds to DNA to induce the transcription of various target genes( Reference Covert, Leung and Gaston 44 , Reference Li and Verma 45 ). In the present study, DHA failed to prevent P. intermedia LPS from degrading IκB-α. Instead, DHA mitigated LPS-induced NF-κB signalling via the inhibition of the nuclear translocation and DNA-binding activity of NF-κB p50 subunit. DHA treatment did not inhibit the nuclear translocation and DNA-binding activity of NF-κB p65. NF-κB mainly comprises two subunits: p50 and p65. Both NF-κB p65 and 50 have been shown to translocate into the nucleus in response to proinflammatory signals, where they bind to specific regulated DNA sequences to initiate the transcription of NF-κB-responsible genes. While NF-κB p65 subunit has a characteristic transcriptional activation domain and directly stimulates target gene transcription, NF-κB p50 subunit does not have such a domain and, therefore, usually forms a heterodimer with other NF-κB subunits to participate in transcription( Reference Li and Verma 45 – Reference Ghosh, May and Kopp 47 ).
The STAT cascade has been shown to be involved in the regulation of inflammatory responses( Reference Pfitzner, Kliem and Baus 48 ). The transcription factors of this family are activated through JAK and subsequently trigger dimerisation and accumulation in the cell nucleus where they induce the transcription of regulated target genes( Reference Schindler and Darnell 49 , Reference Schindler, Levy and Decker 50 ). STAT1, downstream signalling molecule of JAK2, is an essential signalling component for LPS-mediated gene expression in macrophages( Reference Gao, Filla and Fultz 51 ). In the present study, we examined whether DHA regulates P. intermedia LPS-induced production of NO, IL-1β and IL-6 through the inhibition of the STAT1 signalling pathway. DHA significantly blocked P. intermedia LPS-induced STAT1 phosphorylation, supporting the hypothesis that DHA exerts its effects on the LPS-induced production of NO, IL-1β and IL-6 by regulating STAT1 signalling.
Periodontal disease may affect the serum concentrations of DHA. However, the interrelationship between plasma DHA concentrations and periodontal disease has been reported only by a few studies and remains controversial. Requirand et al. ( Reference Requirand, Gibert and Tramini 52 ) showed that the concentrations of DHA are lower in patients with periodontitis than in the control group, whereas Ramirez-Tortosa et al. ( Reference Ramirez-Tortosa, Quiles and Battino 53 ) found that there is no difference in the concentrations of DHA between periodontitis patients and control group. In addition, it has been reported that the serum concentrations of DHA may be affected by the severity of periodontal disease( Reference Figueredo, Martinez and Koury 54 ). Significantly higher concentrations of DHA have been observed in generalised chronic periodontitis patients than in gingivitis patients, and the pocket depth has been reported to show a significant positive correlation with DHA concentrations.
In conclusion, DHA significantly attenuated P. intermedia LPS-induced production of NO, IL-1β and IL-6 in murine macrophages via anti-inflammatory HO-1 induction and inhibition of JNK, NF-κB and STAT1 activation. DHA can be used to reduce periodontal tissue destruction by down-regulating the destructive aspects of the host inflammatory response to periodontal pathogenic bacteria and up-regulating protective responses. Further in vivo studies are needed to better evaluate the potential of DHA in humans as a therapeutic agent to treat periodontal disease.
Acknowledgements
The present study was supported by a grant from the Korean Health Technology R&D Project, Ministry of Health and Welfare, Republic of Korea (A121041). The Ministry of Health and Welfare had no role in the design and analysis of the study or in the writing of this article.
S.-J. K. and I. S. C. designed the study and wrote the manuscript. E.-Y. C. and J.-Y. J. carried out the experimental work and contributed to the preparation of the manuscript. J.-I. C. contributed to the design and preparation of the manuscript.
None of the authors has any conflicts of interest to declare.







