


A tribe including many members who, from possessing in a high degree the spirit of patriotism, fidelity, obedience, courage, and sympathy, were always ready to aid one another, and to sacrifice themselves for the common good, would be victorious over most other tribes; and this would be natural selection.
— Charles Darwin (Reference Darwin1874, pp. 178–79)1. Introduction
Few scholars doubt that the scale of cooperation found in contemporary human societies is unusual. We typically live in large, rule-bound, anonymous societies in which the mean genetic relatedness between members is low (Moffett Reference Moffett2013). Ethnographic evidence from hunter-gatherer societies suggests that our Late Pleistocene ancestors already had many interactions with non-kin that were regulated by systems of norms and institutions, even in societies numbering only a few hundred to a few thousand (Hill et al. Reference Hill, Wood, Baggio, Hurtado and Boyd2014). If that is the case, then there are two key evolutionary questions: (1) How did cooperation among non-kin evolve in small-scale societies in prehistoric times, and (2) how did small-scale societies evolve into much larger and more complex societies during the Holocene? We outline here the evidence that supports the hypothesis that cultural group selection (CGS) was and still is an important force in the evolution of human societies. We compare CGS to other evolutionary mechanisms that have been proposed to account for human sociality. See Boehm (Reference Boehm2012), Bowles and Gintis (Reference Bowles and Gintis2011), and Cronk and Leech (Reference Cronk and Leech2013) for reviews of the various mechanisms proposed to date.
Heretofore, the debate about the importance of CGS relative to competing explanations for human cooperation has not been fully engaged. Some debates have focused on narrow issues such as the interpretation of experimental results (Hagen & Hammerstein Reference Hagen and Hammerstein2006) and the assumptions of mathematical models (Boyd et al. Reference Boyd, Richerson and Henrich2011; Lehmann et al. Reference Lehmann, Feldman and Foster2008). Reviews of evolutionarily plausible accounts of human cooperation often do not discuss CGS (e.g., Sigmund Reference Sigmund2007) or treat it as an unsupported conjecture or semantic error (e.g., West et al. Reference West, Griffin and Gardner2007). Even the sympathetic commentator, E. A. Smith (Reference Smith2010), remarks that CGS is under-theorized and in need of much more empirical analysis. We provide a guide to the relevant theory and argue that the empirical evidence from a range of domains provides robust qualitative support for the point that CGS describes real and important processes in human social evolution. In many domains, the best evidence is quantitative. We cannot provide a precise quantitative account of the importance of CGS compared to competing (but generally not mutually exclusive) processes in the evolution of human cooperation. That is an ongoing project with recent pioneering studies in critical areas. We will discuss the following hypotheses:
-
1. Cultural group selection. The CGS hypothesis is an updated version of the one that Darwin (Reference Darwin1874) formulated in The Descent of Man, the essentials of which are quoted in our epigraph (Richerson & Boyd Reference Richerson, Boyd, Bell, Futuyma, Eanes and Levinton2010). Darwin lacked the modern distinction between genes and culture, but in Descent he argued that the organic differences between human groups were small, while the differences in durable cultural features – customs and traditions were the terms he used – were large. He also noted that human groups – he uses the term tribe in the quote in our epigraph – compete. If this is the case, then group selection on cultural variation is an important force. We sketch the CGS hypothesis in more detail in section 2.
-
2. Culture doesn't evolve in the Darwinian sense. One critique of cultural evolutionary hypotheses generally is that transmitted culture, if it exists, lacks the properties to evolve in a Darwinian fashion at all, much less at the group level (Brown & Richerson Reference Brown and Richerson2014; Pinker Reference Pinker2012; Tooby Reference Tooby2014; Tooby & Cosmides Reference Tooby, Cosmides, Barkow, Cosmides and Tooby1992). The objection of some evolutionary biologists to CGS (e.g., West et al. Reference West, El Mouden and Gardner2011) seems to turn on a similar argument. If culture is taken to be a proximate mechanism rather than an ultimate cause, and if it is true that proximate mechanisms cannot influence the ultimate evolutionary processes, then culture cannot play any active role in the evolutionary process. For a general response to this argument, see Laland et al. (Reference Laland, Sterelny, Odling-Smee, Hoppitt and Uller2011).
-
3. Kin selection and reciprocity suffice. Many alternatives to CGS argue that the familiar evolutionary processes acting on genes – reciprocity and inclusive fitness – which generally explain cooperation in nonhumans, are also sufficient to explain the case of human cooperation (e.g., Alexander Reference Alexander1987). Henrich et al. (Reference Henrich, Bowles, Boyd, Hopfensitz, Richerson, Sigmund, Smith, Weissing, Young and Hammerstein2003) provide a useful discussion of the most prominent of these. The issue here is not whether effects such as nepotism, reputation building, and other mechanisms of cooperation supported by reciprocity and inclusive fitness exist – no one doubts this – but whether they are sufficient to explain the large-scale cooperation in human societies. Some hypotheses hold that CGS became important fairly late in human evolution and that a considerable amount of cooperation evolved earlier due to kin selection and reciprocity (Tomasello et al. Reference Tomasello, Melis, Tennie, Wyman and Herrmann2012). Campbell (Reference Campbell1975) suggested that CGS only became important once state-level societies developed, about 5,000 years ago. In simpler societies at least, individuals might bargain and negotiate their way to efficient social equilibria, as some game theorists argue (see Bowles & Gintis Reference Bowles and Gintis2011, Ch. 5 for a critical review).
-
4. Pleistocene social mechanisms mismatched in Holocene environments. A related alternative hypothesis holds that our innate social psychology evolved in the Pleistocene in societies that were so small that the familiar mechanisms mentioned previously could entirely account for their cooperative aspects. Subsequently, in the radically different environments of the Holocene, the cognitive mechanisms that evolved in the Pleistocene often caused people to behave in novel ways in environments that were outside the range of those in which the mechanism evolved. Some evolutionary psychologists argue that large-scale cooperation may be a by-product of a Pleistocene cognitive mechanism expressed in much larger, more anonymous Holocene societies (Delton et al. Reference Delton, Krasnow, Cosmides and Tooby2011; Krasnow et al. Reference Krasnow, Cosmides, Pedersen and Tooby2012; Tooby & Cosmides Reference Tooby and Cosmides1989; but see Zefferman Reference Zefferman2014a).
-
5. A role for genetic group selection. Some alternate theories invoke genetic group selection (on groups larger than those composed of close kin) as at least a partial explanation for human sociality (Rushton Reference Rushton1989; D. S. Wilson Reference Wilson2002; E. O. Wilson Reference Wilson2012). Several authors have imagined hybrid models in which culturally transmitted institutions reduce within-group phenotypic variation to such low levels that modest amounts of between-group genetic variation can come under selection (Boehm Reference Boehm1997; Bowles & Gintis Reference Bowles and Gintis2011, Ch. 7; D.S. Wilson & Kniffin Reference Wilson and Kniffin1999). We propose that CGS can exert selection on genes via culturally transmitted cost and benefit schedules (coevolutionary social selection). Culture-led gene-culture coevolution could produce much the same result as group selection more directly on genes.
-
6. Trigger hypotheses. Many hypotheses regarding human origins invoke some key selection event deep in our evolutionary history that set us on a path that led to modern human culture and societies. Proposals of this sort include: bipedality (Washburn Reference Washburn1959); intelligence (Pinker Reference Pinker2010); cooperative breeding (Burkart et al. Reference Burkart, Hrdy and Van Schaik2009); cooperative hunting (Stanford Reference Stanford1999), language (Smith Reference Smith2010); cooking (Wrangham Reference Wrangham2009); and docility (Simon Reference Simon1990). Such hypotheses are difficult to test independently of the coevolutionary extension of the CGS hypothesis. In living hunter-gatherers, and other modern humans, culture is closely involved in all of these phenomena (Hill et al. Reference Hill, Barton and Hurtado2009). The paleoanthropological record is remarkably hard to interpret with regard to the order in which the various components of modern human sociality evolved and which components coevolved. Bipedal locomotion may have been necessary, but it was not a sufficient trigger because bipedality evolved in the Australopithecine lineage well before the record contains evidence of any advance in cultural behavior over that in nonhuman apes. On the other hand, even the most innatist theories of language evolution imagine a culture-led gene-culture coevolutionary process driving language evolution (Pinker & Bloom Reference Pinker and Bloom1990). Thus, we have little to say about hypotheses of this type.
In sum, if CGS is an active process in human social evolution, then we should see evidence that: (1) culture acts as an inheritance system; (2) group-variable culturally transmitted social norms and institutions are among the domains of culture so transmitted; and (3) success or failure in intergroup competition is frequently caused by institutional variation. As a result of these three features, we should also find evidence of: (4) group-level functionality in cultures, particularly in their social norms and institutions. The CGS hypothesis is thus vulnerable to disproof in several empirical domains. Competing hypotheses argue that cultural systems lack one or more of these features, or possess them to such a limited extent that the explanation for human sociality must depend in the main on other processes.
2. The cultural group selection hypothesis
In this section, we characterize the CGS hypothesis. Three distinct CGS processes exist, united by being driven by group-level heritable cultural variation, but differing in the mechanism of selection. Two of these forms are driven by human choices – choices in part based on prosocial emotions like Darwin's sympathy and patriotism. Because prosocial impulses play such a large role in CGS, it is important to account for their evolution. In the second part of this section, we describe the “tribal social instincts hypothesis,” a modernization of Darwin's proposal, which can, in principle, explain the evolution of prosocial emotions.
Humans exhibit a wide variety of culturally transmitted organizational phenomena larger than, and different from, those reflecting biological relatedness and close pairwise partnerships. These include corporate kin groups, ethnicities, nations, classes, castes, religions, clubs, nations, and empires. CGS is potentially acting on more than one of these groups simultaneously, leading many authors to speak of multi-level selection. The most ancient groups are the ethnolinguistic tribes into which simpler societies are generally divided. As we discuss below, selection on tribes can account for the innate foundation of our social psychology via the process of gene-culture coevolution. The evolution of complex societies in the last five millennia has generated a diverse array of social groups that can vary culturally, compete, and be subject to group selection. We discuss business firms and religions as examples. While modern nations and the subgroups of which they are composed differ in many ways from ancestral tribes, they rest on the innate foundations that evolved from competition between ethnolinguistic units (Richerson & Boyd Reference Richerson and Boyd1999). Given their short history, selection on modern groups will not have had a major impact on genes, but we argue that it has been very important in the ongoing evolution of complex societies.
2.1. How human culture facilitates CGS
How do cultures generate the group-level variation that CGS requires, and what sort of selective processes act on that variation? In this section, we define the CGS hypothesis by reviewing the mechanisms that make cultural inheritance and cultural evolution quite different from genetic evolution.
The mechanisms that maintain intergroup variation are the following:
-
1. Accurate, rapid social learning. Human cognition includes adaptations for imitation and teaching that allow the cumulative evolution of complex traits, including norms and institutions that regulate social behavior (Tomasello Reference Tomasello1999). See section 3.
-
2. Conformist social learning. If people tend to differentially copy locally common behaviors, the homogenizing effect of migration will be reduced and group differences will be maintained (Boyd & Richerson Reference Boyd and Richerson1985).
-
3. Coordination payoffs. If the payoff to any strategy depends on the local frequency of its use, then even subpopulations in identical environments may reach different stable equilibria over time. Frequency-dependent payoffs and multiple stable equilibria are probably very common in human social institutions (Cooper Reference Cooper1999).
-
4. Punishment of deviant behaviors. Punishment of those who engage in uncommon behaviors can stabilize essentially any norm within a group, regardless of its other adaptive consequences (Boyd & Richerson Reference Boyd and Richerson1992b). Since these norms will often be influenced by historical happenstance, groups will often end up with different sets of morally enforced customs. Punishment is a strong force if the punishing is cheap to the punishers but costly to the punished.
-
5. Strong prestige bias or one-to-many transmission. If people learn disproportionately from those who are most prestigious or successful in their local population, then the “effective size” of the cultural population may be small and cultural drift will diversify populations (Cavalli-Sforza & Feldman Reference Cavalli-Sforza and Feldman1981).
-
6. Symbolic markers of group boundaries. As groups differentiate, the importance of learning the locally proper behaviors and customs becomes increasingly important. If neutral marker traits correlate with behavioral norms that are difficult to observe, then selection may favor attentiveness to these markers as reliable signals of whom to imitate (McElreath et al. Reference McElreath, Boyd and Richerson2003). Dialect is an example (see sect. 4.2). This contributes to group differentiation by facilitating in-group assortment and reducing the chances that migrants are imitated, especially in those regions where neighboring groups interact.
-
7. Institutional complexity that generates inertia and reduces borrowing. Institutions are often a complex array of norms organized to solve cooperation dilemmas and navigate the intricacies of coordination (see sect. 4.3). The functioning of complex institutions is highly dependent on tacit knowledge (Aoki Reference Aoki2001). Even in-group members may have an incomplete explicit understanding of how an institution works, and the overall rules driving the institution may be distributed among the norms of various specialized participants. This makes the institution quite opaque to out-groups, limiting the diffusion of institutions between societies.
The high group-level variation between human groups sets the stage for the evolution of group-beneficial behaviors. There are three distinct processes by which group-beneficial behavior can spread:
-
1. Natural selection. Groups exhibit variation in behaviors that affect the rate at which they grow, produce daughter groups, overcome resource constraints, avoid internal political conflict, succeed in war, and ultimately replace other groups. This provides a relatively slow form of selection for traits that contribute to group success (Soltis et al. Reference Soltis, Boyd and Richerson1995).
-
2. Selective imitation of successful groups. If people prefer to copy successful individuals, and if they have some contact with out-group members, then members of less successful groups will sometimes copy members of more successful groups. This favors the spread of group-beneficial norms and institutions (Boyd & Richerson Reference Boyd and Richerson2002). This force may be especially powerful if the new cultural characteristic is adopted by particularly influential group members who are more likely to be copied by others in their group.
-
3. Selective migration between groups. Individuals in dysfunctional groups may migrate to another group if they observe that the other group more successfully maintains behaviors that benefit all members (Boyd & Richerson Reference Boyd and Richerson2009). The resulting diminution of unsuccessful groups may weaken their competitive ability and/or encourage leaders to adopt successful foreign norms, strengthening the two processes defined previously.
All three selective processes will tend to reduce group-level variation. Thus, the standing group-level variation is a balance between variation-creating and variation-reducing forces. It is an empirical matter whether cultural evolutionary processes can maintain substantial between-group variation or whether, as is the case with genetic variation, appreciable group variation is hard to maintain. See section 4.1, particularly for a quantitative discussion of this issue.
Throughout the remainder of this article, we will generally use economic terms, such as “cost,” “benefit,” and “success,” to characterize the contribution of traits to CGS. Evolutionary biologists generally express such terms in the currency of genetic fitness. Cultural evolution and gene-culture coevolution are considerably more complex. A cultural variant that attracts many imitators has a reproductive success which is not necessarily tied to biological reproduction. For example, prosocial leaders gain prestige, which may attract imitators not related genetically to the leader. In the case of the three mechanisms of CGS and the mechanism of gene-culture coevolution, we can use models to specify the costs, benefits, and success of cultural variants in terms of changes in their frequency. Model-fitting approaches have been used to estimate the strength of complex evolutionary forces in laboratory cultural evolution experiments (McElreath et al. 2008). So far, too few quantitative studies have been performed on CGS and competing processes to allow for much but qualitative judgments. One of our purposes here is to make the case that qualitative investigations already performed amply support the pursuit of more definitive quantitative data. We will discuss the relatively small number of pioneering quantitative studies of cultural evolution that have been conducted to date (see, e.g., our Figure 1 in section 4.1 of this article). This work has demonstrated the feasibility of quantitative work on every phase of cultural evolution. For example, ambitious efforts are underway to use historical and archaeological data to construct synthetic time series that will be suitable for quantitative model fitting investigations (Collard et al. Reference Collard, Edinborough, Shennan and Thomas2010; Turchin Reference Turchin2009; Turchin et al. Reference Turchin, Whitehouse, Francois, Slingerland and Collard2012).
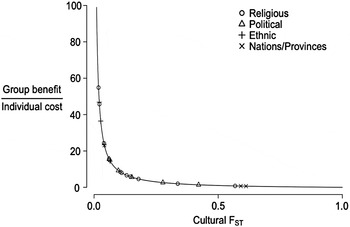
Figure 1. The scope for group selection. The curve is the minimum variation required for a group-beneficial trait to be favored by natural selection as a function of the level of cultural differences between groups (F ST). The points are cultural F ST measurements across several kinds of group identities. For details, see the appendix.
Note that many cultural evolutionary processes generate cultural variation and cause evolution. CGS is only relevant to situations in which groups compete; it is not a complete theory of the evolution of cultural variation.
2.2. The tribal social instincts hypothesis
If the three CGS processes described in the previous section have operated over spans of time consistent with the evolution of genes, then culture-led gene-culture coevolution may have favored genes that encourage group advantageous behavior.
How did human culture become evolvable at the group level? Richerson and Boyd (Reference Richerson, Boyd, Eibl-Eibesfeldt and Salter1998) proposed a “tribal social instincts” hypothesis to account for the emergence of innate social predispositions that support the norm- and institution-bound social life of humans. Once humans could culturally transmit elementary norms, simple moral communities would exist. Moral norms that evolved under the influence of CGS would exert social selection on human genes via mechanisms such as selective child neglect, mate choice, and punishment (Feldman & Laland Reference Feldman and Laland1996; Newson Reference Newson, Clancy, Hinde and Rutherford2013). Perhaps the earliest cultural norms merely solidified the bonds of kinship and reciprocity that were evolving through participation in systems of cooperative breeding (Chapais Reference Chapais2008; Hrdy Reference Hrdy2009). Infants younger than one year prefer puppets that are helpful to others over those that are antisocial (Hamlin et al. Reference Hamlin, Wynn, Bloom and Mahajan2011), suggesting that our social predispositions are partly innate. See section 4.2 for more evidence on this point.
The complexity of technology that a group can maintain is a function of its size (Henrich Reference Henrich2004; Shennan Reference Shennan2001). Docile individuals more inclined to conform to norms would find it easier to enter larger more norm-bound groups and hence be able to acquire more sophisticated technology (Simon Reference Simon1990). A population of more docile individuals could then permit the cultural evolution of slightly more sophisticated and effective norms. This would allow groups to maintain more reliable cooperation and tools, which would, in turn, be favored by CGS acting on this variation.
A recent quantitative estimate of rates of cultural evolution using a large body of historical and archaeological data showed that it evolves considerably faster than genes (Perreault Reference Perreault2012). Cultural evolution will thus tend to be the leading process, and genetic evolution the lagging process, in gene-culture coevolution. Over repeated rounds of such coevolution, stretching back perhaps several hundred thousand years, our Pleistocene ancestors came to have institutionalized social systems with (if ethnographic analogs are reliable) complex sets of linked norms organizing tribal societies of a few hundred to a few thousand people (Hill et al. Reference Hill, Wood, Baggio, Hurtado and Boyd2014). At some point, language became sufficiently rich that complex norms could be explained to children, and participants could make promises, negotiate conflicts, and discuss institutional innovations (see sect. 6.1).
Several hypotheses compete with this coevolutionary account. Ayala (Reference Ayala2010) argues that natural selection acted directly on human cognitive capacities to the point that all of the innate underpinnings necessary for the maintenance of norms and institutions were in place before the cultural transmission of them was important. Other accounts root human sociality in genetically evolved skills for social exchange, with no important role for cultural evolution (Tooby & Cosmides Reference Tooby, Cosmides, Barkow, Cosmides and Tooby1992).
Evidence currently exists that supports the hypothesis of culture-led gene-culture coevolution for a few simple genetic traits whose function is well known, but unfortunately not yet for genes related to behavior. The best understood examples of culture-led gene-culture coevolution are adult lactase persistence (Durham Reference Durham1991) and the hemoglobin polymorphisms that confer resistance to malaria (Allison Reference Allison1964). Recent molecular evidence suggests that the genes associated with these and other phenotypic changes swept to appreciable frequencies in the Holocene, as agricultural diets exerted selective pressures on digestive physiology (Itan et al. Reference Itan, Powell, Beaumont, Burger and Thomas2009; Tishkoff et al. Reference Tishkoff, Reed, Ranciaro, Voight, Babbitt, Silverman, Powell, Mortensen, Hirbo, Osman, Ibrahim, Omar, Lema, Nyambo, Ghori, Bumpstead, Pritchard, Wray and Deloukas2007) and human populations became dense enough to carry epidemic diseases (Wolfe et al. Reference Wolfe, Dunavan and Diamond2007). The dramatic changes of modern environments are resulting in detectable selection on genes related to health and fertility (Byars et al. Reference Byars, Ewbank, Govindaraju and Stearns2010; Kohler et al. Reference Kohler, Rogers and Christensen1999). Genome-wide scans for genes under selection and comparative studies on chimpanzee–human differences detect strong recent selection at a large number of loci affecting genes that are expressed in the nervous system and in hormone producing glands. These are candidates to test the tribal social instincts hypothesis and its competitors, but the specific functions of the selected alleles are as yet unknown (Laland et al. Reference Laland, Odling-Smee and Myles2010; Ross & Richerson Reference Ross and Richerson2014). For example, if genes for docility were selected early in the hominin lineage before we have evidence for sophisticated culture, they are less likely to have been a product of culture-led gene-culture coevolution than if they evolved in the last 150,000 years as culture increased to modern levels of sophistication.
Several possible cases of genes related to behavior coming under culture-induced selection in the Holocene have been proposed. Cochran and Harpending (Reference Cochran and Harpending2009) suggest that the rise of modern urban occupations have favored genes for higher intelligence in some populations. Chiao and Blizinsky (Reference Chiao and Blizinsky2010) argue that neural processing of social status cues differs between more and less hierarchical societies. The genetic mechanism proposed as a cause of such differences is variation in a serotonin transporter gene.
Genomic tools promise to substantially improve our ability to test the tribal social instincts hypothesis by allowing us to estimate the dates at which major selective events occurred (Richerson et al. Reference Richerson, Boyd and Henrich2010). Ongoing advances in whole genome sequencing will provide much larger and higher resolution samples of genomes. Improvements in statistical and modeling techniques promise to improve the resolution for detecting signatures of selection, and for discovering their selective consequences more easily (Pritchard et al. Reference Pritchard, Pickrell and Coop2010). Over the next decade, we can expect a flood of new data that can be used to test hypotheses related to gene-culture coevolution.
3. Culture acts as an inheritance system
Two quite distinct lines of evidence show that human culture acts as an inheritance system. First, evidence from developmental studies shows how a cognitive system for imitation arises in the first 12 months of life and dovetails with adult behaviors that support children's social learning. Other apes have a much more rudimentary social learning system. Second, historical evidence shows that human culture exhibits a phylogenetic pattern of descent with modification in which ancestral commonalities can be traced back thousands of years. Together, these lines of evidence cast strong doubt on the idea that most human behavioral differences have arisen via an extreme version of the “evoked culture” hypothesis (Tooby & Cosmides Reference Tooby, Cosmides, Barkow, Cosmides and Tooby1992). By “evoked culture,” these authors mean that human minds have a universal psychological architecture, selected in Pleistocene environments, which allows for the expression of phenotypic differences during development in various evolutionarily relevant domains such as social exchange and mate choice. It is also difficult to see how the developmental and historical evidence can be reconciled with an extreme version of the later improvisational intelligence hypothesis of Cosmides and Tooby (Reference Cosmides, Tooby, Sternberg and Kaufman2001). Individual humans might invent complex adaptations on the spot as needed, with little or no need to depend on cultural inherited traditions. If this were so, children would have little need to acquire complex adaptations with a system adapted for high-fidelity imitation. Nor would we observe deep phylogenetic patterns in comparative and historical data; high rates of individual improvisational innovation would rapidly erase any historical signal due to culture transmission by accurate imitation. This is not to defend a blank slate hypothesis. Gene-based cognitive constraints on cultural evolution are surely many. Nor do we mean to imply that humans are not highly intelligent. Nor are we even sure that anyone defends extreme versions of the evoked culture or improvisational intelligence hypotheses. It is easy to imagine that a phenomenon like religion is co-produced by genetically inherited cognitive biases and the cultural transmission of specific religious doctrines (Gervais et al. Reference Gervais, Willard, Norenzayan and Henrich2011), and the great speed of cultural relative to genetic evolution no doubt owes much to our capacity for foresightful innovation (Perreault Reference Perreault2012).
3.1. A cognitive system for accurate, voluminous cultural transmission
Children have powerful cognitive systems evolved to acquire quite complex cultural knowledge from their elders. Adults and knowledgeable children also appear to be adapted to behave in ways that scaffold the learning of naïve children (Csibra & Gergely Reference Csibra and Gergely2011; Dean et al. Reference Dean, Kendal, Schapiro, Thierry and Laland2012; Kline Reference Kline2015). The transmission of complex cultural information and its cumulative evolution requires accurate imitation (Tennie et al. Reference Tennie, Call and Tomasello2009). Nonhuman social learners tested so far lack the skills necessary to acquire a large repertoire of complex skills, and there are so far no reports of nonhumans having acquired complex traits (traits composed of more than two or three separate innovations) by social learning. Human children routinely learn concepts with help from knowledgeable others that qualitatively transcend innate intuitions (Carey Reference Carey2009). In the 1970s and 1980s, a highly innatist hypothesis for language learning was a major part of the cognitive revolution (Pinker Reference Pinker1994). It has failed to account, however, for the wide diversity of syntactic devices comparative linguists have since discovered (e.g., Evans & Levinson Reference Evans and Levinson2009; Newmeyer, Reference Newmeyer2004). Most grammatical constructions appear to be cultural features underpinned only indirectly by rather general cognitive features (e.g., Hurford Reference Hurford2011). Research over the last two decades has produced a fairly detailed picture of how human imitation develops over the first 12 months of life (Carey Reference Carey2009; Tomasello et al. Reference Tomasello, Carpenter, Call, Behne and Moll2005) and how older individuals support the imitative efforts of children (Csibra & Gergely Reference Csibra and Gergely2011). The same basic system seems to operate across all domains of culture (Harris Reference Harris2012).
Human social learning contrasts sharply with that of chimpanzees and other nonhuman animals. Chimpanzee social learning focuses primarily on goal-related behaviors, such as retrieving food. In comparative studies with human children, chimpanzees tended to imitate only the behaviors necessary to reach a goal, while children diligently imitated all of an adult's behaviors whether or not they directly resulted in reaching a tangible goal (Whiten et al. Reference Whiten, McGuigan, Marshall-Pescini and Hopper2009). If adults suggest that it is important, children will replicate adult actions in ritualistic detail (Herrmann et al. Reference Herrmann, Legare, Harris and Whitehouse2013). This finding initially seems counterintuitive – humans appear to unnecessarily “over-imitate,” while chimpanzees seem better able to boil down a stream of actions to only the necessary, goal-related behaviors. However, over-imitation is probably an essential adaptation for building a repertoire of elementary skills and maneuvers that will eventually be incorporated into complex crafts and social institutions (Harris Reference Harris2012). Many cultural adaptations are so complex that the functional significance of many elements of them is opaque to learners and often to accomplished practitioners as well. Carefully following demonstrated recipes preserves functionality, albeit at some cost of replicating non-functional elements.
Thus, studies of child development strongly suggest that human social learning does accurately transmit cultural variants over a wide range of design space in many domains (Carey Reference Carey2009). In section 4.2, we address the issue of the cultural transmission of social norms.
Many experimental studies of human social learning have been conducted on adults, mostly student volunteers (Kempe & Mesoudi Reference Kempe and Mesoudi2014). Individuals use a variety of strategies to acquire social information, tending to favor success-based strategies when such information is available to them; conformist strategies are typically employed when payoff-based information is unavailable or uninformative (Molleman et al. Reference Molleman, van den Berg and Weissing2014).
3.2. Descent with modification
Darwinian evolutionary processes shape adaptations incrementally. Marginal change occurs over each small time step in history, followed by reasonably faithful transmission and further marginal changes. Over many such time steps, the evolving system can, but need not necessarily, change dramatically. Often successful lineages split, often repeatedly, to form many sister groups. The Romance language societies are an example. A necessary but not sufficient test of the CGS hypothesis is whether the social systems of human societies follow a phylogenetic pattern, the way genes do, thus demonstrating descent with modification. If the transformational processes operating in human history were as strong as the improvisational intelligence hypothesis (Cosmides & Tooby Reference Cosmides, Tooby, Sternberg and Kaufman2001; Pinker Reference Pinker2010) holds, then humans would routinely switch such things as their political organization quite rapidly and in large jumps, thus not leaving a phylogenetic pattern. Similarly, the evoked culture hypothesis holds that human behavioral variation is based on a pan-human evolved psychology that is expressed in different ways in different environments (Tooby & Cosmides Reference Tooby, Cosmides, Barkow, Cosmides and Tooby1992). If this hypothesis were true, phylogenetic patterns would also be weak or undetectable; variation would reflect current ecology, not cultural history. A pattern of descent with modification by itself does not test the CGS process against competing evolutionary processes acting at lower levels of organization than groups; it only establishes the action of an evolutionary rather than a non-evolutionary process.
Historical linguists established that languages evolved by descent with modification before Darwin and his contemporaries began to apply the idea to biological evolution (Müller Reference Müller1862). Recently, phylogenetic methods, originally borrowed from biology, have been applied to detecting descent-with-modification patterns in cultural data (Mace & Jordan Reference Mace and Jordan2011). These methods have reached a high degree of sophistication (e.g., Bouckaert et al. Reference Bouckaert, Lemey, Dunn, Greenhill, Alekseyenko, Drummond, Gray, Suchard and Atkinson2012). Language phylogenies appear to be a reasonable proxy for population movements, often matching phylogenetic patterns based on genes reasonably well (Cavalli-Sforza et al. Reference Cavalli-Sforza, Piazza, Menozzi and Mountain1988), albeit not perfectly.
The phylogenies of several large language families have been investigated, including Bantu (Holden Reference Holden2002), Austronesian (Gray et al. Reference Gray, Drummond and Greenhill2009), Tupi (Walker et al. Reference Walker, Wichmann, Mailund and Atkisson2012), and Indo-European (Bouckaert et al. Reference Bouckaert, Lemey, Dunn, Greenhill, Alekseyenko, Drummond, Gray, Suchard and Atkinson2012). The Walker et al. study is notable for quantitatively estimating rates evolution in several Tupi cultural domains (usually less than five transitions in 10 millennia). From the correspondence of linguistic time-depth estimates and archaeological dates, each of these language families appears to constitute a cultural adaptive radiation based upon agricultural subsistence and associated social-organizational features. These innovations allowed their speakers to expand at the expense of hunter-gatherers in the Early and Middle Holocene. In the case of the eastern Austronesians, expansion was partly to remote, previously uninhabited oceanic islands. Early farmers in Europe were clearly the result of a demic expansion (Haak et al. Reference Haak, Balanovsky, Sanchez, Koshel, Zaporozhchenko, Adler, Der Sarkissian, Brandt, Schwarz, Nicklisch, Dresely, Fritsch, Balanovska, Villems, Meller, Alt and Cooper2010). Currie and Mace (Reference Currie and Mace2009) found that the area over which a language is spoken is more closely related to its political complexity than to any other variable tested, consistent with the hypothesis that CGS on social organization features correlated with language played an important role in the creation of patterns of linguistic diversity. Historically attested expansions of empires, typically accomplished by a mixture of military conquest, assimilation of conquered peoples, and migration into the more prosperous dominant society, appear to exhibit all three CGS processes (Turchin Reference Turchin2006).
Language trees can be used as a foundation for studying other kinds of cultural evolution. Such studies show that features of social organization often include strong phylogenetic signals that are correlated with the linguistic signal. Again, this is a necessary but not sufficient condition for CGS to have been an important process. Guglielmino et al. (Reference Guglielmino, Viganotti, Hewlett and Cavalli-Sforza1995) studied 47 cultural traits, many of them related to social organization, in 277 African societies. They used correlation with language groups to estimate “vertical” (within-culture) transmission, similarity to neighboring groups to estimate “horizontal” diffusion from linguistically unrelated groups, and correlation with vegetation type to infer the effects of adaptation to local environments. They detected a strong effect of vertical inheritance of cultural traits related to family and kinship and the economy, and a medium vertical effect on traits related to social stratification and house type. The effect of horizontal diffusion was high on the division of labor by sex and house type. The direct effect of environments was strongest on economy-related traits and medium to low on other traits. In other words, the effect of either vertical or horizontal transmission was strong on all traits examined. Currie et al. (Reference Currie, Greenhill, Gray, Hasegawa and Mace2010) used the rise and fall of political complexity in Austronesian societies to directly test the descent-with-modification hypothesis. They found that in the best-fitting model, political complexity increased and decreased in incremental steps. The second-best-fitting model allowed for decreases (but not increases) in complexity to skip intermediate steps.
As more historical and archaeological data are converted to time series suitable for quantitative analysis, tests of descent with modification will become more common and more precise (Turchin 2008). It should also be possible to estimate the parameters of mechanistic evolutionary models to such data to more directly test the CGS hypothesis and its competitors. See Bouckaert et al. (Reference Bouckaert, Lemey, Dunn, Greenhill, Alekseyenko, Drummond, Gray, Suchard and Atkinson2012) for a simple example. Of particular interest will be investigations that trace the evolution of Roman law and similar institutions, as they were transmitted both vertically and horizontally. Certainly, no evidence carefully examined to date suggests that non-evolutionary patterns dominate the archaeological and historical record, as would be suggested by strong versions of the evoked culture and improvisational intelligence hypotheses.
4. Culture exhibits group-level variation
If CGS processes are to operate on cultural variation, then we must document appreciable variation at the level of groups. Group selection on genes is generally implausible because migration between groups prevents substantial genetic variation from arising in groups larger than those composed of close kin. In section 2.1 we reviewed the mechanisms by which cultural systems can preserve much more variation between groups of unrelated or distantly related individuals than can genetic inheritance. Here we review the evidence that between-group cultural variation is large enough to support group selection and that the facility with which children learn the norms of their own group shows how the cultural inheritance system maintains CGS relevant variation.
4.1. Quantitative evidence of cultural differences between groups
The more groups differ, the more effective CGS is likely to be. The Price equation, a general depiction of selection operating on multiple levels, can be re-arranged to express the required conditions for a group-beneficial trait to be favored by selection in the face of individual-level selection against the trait (Bell et al. Reference Bell, Richerson and McElreath2009; Bowles Reference Bowles2006):
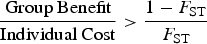 $${\hbox{Group Benefit} \over \hbox{Individual Cost}} \gt {\displaystyle{1 - F_{{\rm ST}} \over F_{{\rm ST}}}}$$
$${\hbox{Group Benefit} \over \hbox{Individual Cost}} \gt {\displaystyle{1 - F_{{\rm ST}} \over F_{{\rm ST}}}}$$
where F ST is the fraction of the total variance that is between groups. If groups do not differ at all for some trait, then F ST = 0, and if the groups have nothing in common, then F ST = 1. Developed to measure inbreeding and genetic differences between populations, F ST can also be used to measure cultural differences. Equation (1) says that traits will be favored if they have zero individual costs and positive group benefits. Traits with some detrimental individual-level effect on reproduction and/or survival can also be favored if group benefits are sufficiently large to satisfy the inequality. Figure 1 describes the level of between-group variation required and plots the relationship between the minimum required benefit-to-cost ratio and the level of cultural differences represented in Equation (1). Note that the scope for group selection is sensitive to changes in low values of F ST, because the minimum condition declines geometrically with a linear increase in F ST. This relationship suggests that evolutionary forces that create relatively small differences between similar groups can provide some scope for group selection in cases where group benefits are high relative to costs.
Aside from a few cases (e.g., Edgerton Reference Edgerton1971), computing cultural F ST from ethnographic data is not possible because most ethnographers studying simple societies report only normative behavior and neglect individual variation. Ethnographers and historians tacitly assume that individual variation for normative behaviors is typically small within groups, is large between groups, and grows larger with time in most lineages. If this is correct, the scope for CGS is large. To test the assumption, we used publically available datasets to measure variation in normative beliefs and behaviors within and between contemporary populations. These data capture within- and between-group differences at various scales, from the national level to small political groups within larger communities.
We calculated the cultural F ST of a wide range of neighboring groups for a wide range of beliefs and behaviors (Fig. 1) using data from responses to international and state-level surveys, such as the World Values Survey, the AfroBarometer, the Public Policy Institute of California statewide surveys, the Scottish Health Survey, and the Anwar Sadat Chair Arab Public Opinion Surveys, among many others. Together, these datasets provide a comprehensive report of opinions and behaviors in group-level organizations that are important in human societies: political, religious, ethnic, geographic, and socioeconomic identities. Some measurements are based on responses to one or two questions in a survey, analogous to one or two loci for a genetic F ST. Others are based on many questions and thus reflect broader level differences between groups. All measurements select questions that relate to a particular theme relevant to group performance, such as attitudes toward immigration, a state-controlled economy, or the adoption of strict religious codes. Details can be found in the table in the appendix of this article.
Consistent with the hypothesis that individual variation for many normative beliefs and behaviors is small within groups and large between groups, we find large between-group differences among all groups of all scales (Fig. 1). Among small political or ethnic groups, nations, and religious majorities, we consistently report F ST values greater than 0.01, with some traits showing F ST scores above 0.6. These values are important, as small increases in cultural F ST beyond 0.01 increase the possibility for CGS dramatically. Our data reach well into the plateau of the L-shaped region of the group selection condition.
The large cultural differences measured in this analysis suggest a wide scope for CGS. Our average measurements of cultural F ST place the group-benefit-to-cost requirement well within range of some important cultural behaviors. As described in section 5 further on in this article, some cultural variants provide a distinct advantage over other groups, outweighing the individual costs of implementation. Bell et al. (Reference Bell, Richerson and McElreath2009) compared cultural F ST from the World Values Survey with genetic F ST from Cavalli-Sforza et al.'s (Reference Cavalli-Sforza, Menozzi and Piazza1994) classic study. For neighboring groups that might actually compete, cultural F ST is approximately an order of magnitude larger that genetic F ST. These data suggest that the scope for CGS is substantially higher than for genetic group selection.
4.2. The transmission of social norms
Social norms are among the cultural traits that children readily acquire from adults and other children. Young children are strikingly inflexible with social norms and rules, resulting in young moral absolutists (Levy et al. Reference Levy, Taylor and Gelman1995). Human children are also motivated to engage with others to pursue a shared goal without any reward, apart from the enjoyment of engaging with another person socially (Tomasello & Carpenter Reference Tomasello and Carpenter2007). For example, children as young as 14 months will readily provide instrumental help to an adult, and children of 18 months will help and cooperate with unfamiliar adults. Chimpanzees' abilities in this regard are more limited (Warneken & Tomasello Reference Warneken and Tomasello2009). Attempts to raise chimpanzee infants as if they were children are unsuccessful because young chimpanzees rather quickly mature into independent individuals that are not very responsive to being taught social norms, much less adopting them spontaneously (e.g., Hayes Reference Hayes1951).
Not all potential providers of social learning are equal. Even very young children prefer teachers and social partners who belong to their own social group. For example, preschool children preferred to learn both linguistic and non-linguistic information from individuals who spoke with the same accent as themselves (Kinzler et al. Reference Kinzler, Corriveau and Harris2011), and 12-month-olds preferentially copy the food selection choices of adult native speakers of their own language (Shutts et al. Reference Shutts, Kinzler, McKee and Spelke2009). The effects of out-group discrimination and in-group preference based upon cultural categories like race are evident as early as 3 years of age, when children begin to implicitly associate same-race faces with positive facial expressions and different-race faces with negative facial expressions (Hugenberg & Bodenhausen Reference Hugenberg and Bodenhausen2004). Children show adult levels of racial in-group bias by age 6 (Dunham et al. Reference Dunham, Baron and Banaji2008), although language difference seems to be a more intuitively salient marker than differences in skin color (Kinzler et al. Reference Kinzler, Shutts, DeJesus and Spelke2009).
By the time people are adults, much experimental work by social identity theorists shows that, in many social situations, individuals' behavior is substantially influenced by their affect-laden social group identifications (Haslam Reference Haslam2001). “Who are we?” is an important component of the question “who am I?” The accurate imitation of cultural variation, especially group identities and social norms, is the basis for heritable cultural variation between groups upon which CGS can act. Social learning biases, such as those described previously, act as a general force adapting culture to local environments. They tend to preserve locally successful social behaviors and reduce the diffusion of cultural variation from one social group to another (see sects. 2.1 and 6.1). Identification with a group sometimes results in derogation of other groups. Attitudes toward out-groups are certainly not invariably invidious, however, contrary to some theories of ethnocentrism (Brewer Reference Brewer2007).
Norm transmission has not been extensively studied in adults or undergraduate volunteers. However, early experiments showed a strong conformity effect (Jacobs & Campbell Reference Jacobs and Campbell1961), and rather firm socialization pressure with normative content is observed in some experiments (Baum et al. Reference Baum, Richerson, Efferson and Paciotti2004; Reference Baum, Paciotti, Richerson, Lubell and McElreath2012). Insko et al. (Reference Insko, Gilmore, Drenan, Lipsitz, Moehle and Thibaut1983) demonstrated the evolution of institutions in a multi-generation experiment designed to mimic the origin of states. Efferson et al. (Reference Efferson, Lalive and Fehr2008) showed experimentally that symbolic markers and in-group favoritism coevolve. Derex et al. (Reference Derex, Godelle and Raymond2014) showed that cultural transmission increases substantially in between-group competition compared to within-group competition treatments.
4.3. Institutions are group-level cultural traits
Scott (Reference Scott2001, p. 48) provides a formal definition of institutions: “Institutions are social structures that have attained a high degree of resilience. [They] are composed of cultural-cognitive, normative, and regulative elements that, together with associated activities and resources, provide stability and meaning to social life.” Institutions are important because they create and maintain variation between groups while simultaneously damping down variation among individuals within groups. Institutions only function when large numbers of people conform to them. Institutional economists often characterize institutions as equilibrium solutions to social games that include rewards and punishments that act to induce individual conformity to the institution's norms (Aoki Reference Aoki2001; Greif Reference Greif2006). As a system of commonly accepted norms, they are a property of a group and often vary qualitatively from group to group. Institutions are probably an ancient feature of human social life, ancient enough to have generated the coevolution of genes related to social behavior. The social life of living hunter-gatherers is certainly heavily institutionalized (Hill et al. Reference Hill, Barton and Hurtado2009).
Game theory tells us that in repeated games without fixed end points, the number of equilibria multiplies inordinately. This is easiest to see in the case of institutions involving moralistic punishment. If punishing is sufficiently cheap for punishers and sufficiently costly to the punished, punishment can stabilize any behavior (Boyd & Richerson Reference Boyd and Richerson1992b). The empirical literature on village-scale commons management is particularly well developed and shows that the institutional diversity predicted by theory is amply manifest on the ground (Ostrom Reference Ostrom2005). Similarly, ethnographers have documented ample institutional diversity between small-scale societies that are our closest living approximations to Late Pleistocene societies (e.g., Jorgensen Reference Jorgensen1980). Modern institutions vary substantially cross-culturally (e.g., Nivette Reference Nivette2011).
Henrich et al. (Reference Henrich, Boyd, Bowles, Camerer, Fehr and Gintis2004; Reference Henrich, McElreath, Barr, Ensminger, Barrett, Bolyanatz, Cardenas, Gurven, Gwako, Henrich, Lesorogol, Marlowe, Tracer and Ziker2006) have systematically investigated cross-cultural variation using experimental games, including dictator, ultimatum, third-party punishment, and public goods games. These studies suggest that institutional variation creates substantial differences in behavior. For example, Paciotti et al. (Reference Paciotti, Hadley, Holmes and Borgerhoff Mulder2005) conducted ultimatum game experiments in a Tanzanian village with two ethnic groups. The Sukuma, who are an expanding population of agro-pastoral colonizers, were found to have one of the highest average offers in the ultimatum game ever measured. The Pimbwe, in contrast, made quite low offers. Interestingly, the Sukuma have a unique, elaborate institution of village governance, which the Pimbwe lack, and the Sukuma organize the provision of public goods much more effectively than the Pimbwe.
Herrmann et al. (Reference Herrmann, Thöni and Gächter2008) compared the behavior of participants in public goods game experiments in 16 diverse societies. Participants were given the opportunity to punish other members of their group. Groups from societies that achieved high rates of cooperation over the 10 rounds of the game had high rates of punishment of non-contributors. However, in some societies, participants sanctioned high contributors as well as low contributors. In such societies, rates of cooperation in the games were low. The authors also compared the 16 societies by using responses to the World Values Survey to construct indices of civic cooperation and rule of law. They found that societies where experiment participants used punishment to promote cooperation also scored higher on these indices than societies in which some experimental participants punished cooperators.
In section 2.1, we outlined how various mechanisms can generate and maintain between-group variation in the face of migration. One important institutional mechanism with substantial empirical support is the marking of group members through differences in dialect, dress, religion, and similar symbols. These markings are often readily observed and hard-to-fake symbols (e.g., Iannaccone Reference Iannaccone1994) that are emotionally salient to their bearers. Symbolic markers thus serve to identify members of distinct social groups and subgroups and to define who is and who is not entitled to benefits from the activities of the organization.
Institutions from one group are often quite difficult for other groups to copy. The tendency of cultural items to diffuse between groups is favored by “observability” and “trialability” (Rogers & Shoemaker Reference Rogers and Shoemaker1971). The institutions of another society are often hard to observe because outsiders can seldom see the entire system of rules working together. Institutions are hard to try out, because for an institution to be effective, some critical mass of group members has to adopt its rules. Considerable collective decision-making and learning effort must be expended to integrate a novel institution with the other institutions of the adopting society. Greif's (Reference Greif2006) study of the evolution of early modern Mediterranean trading systems notes that institutional evolution occurs when ongoing changes destabilize an equilibrium previously regulated by an institution. Evolution then takes the form of a collective, culturally local search for a replacement institution. For example, the Genoese trading system was long regulated by personal reputations. As the Genoese traders grew wealthy, reputation monitoring became too time-consuming to mesh with the increasing management responsibilities of traders. Eventually they settled on a familiar formal legal system modeled on Roman law as preserved by the Church in the form of canon law.
If culturally transmitted institutions had a major coevolutionary impact on the innate components of our social psychology (see sect. 2.2), then for some part of our evolutionary history humans must have lived in institutionally structured groups of a few hundred to several thousand individuals that were symbolically marked and shared a common identity. Some authors argue that institutions only arose in the Holocene (Runciman Reference Runciman2005), although archaeological evidence and ethnographic insights from living foragers suggests that tribal-scale societies were common in the Pleistocene. Even the Shoshone, often used as an example of the simplest sort of family-band society (Steward Reference Steward1955), had supra-familial institutions involving upwards of 15 separate families to cooperatively hunt in winter camps and a system for incorporating stranger families into these camps (Murphy & Murphy Reference Murphy, Murphy and d'Azevedo1986); because of the high mobility of families, winter camps regularly incorporated co-ethnic strangers. The Ju/'hoansi (!Kung), another primarily family-band society, frequently organize at the camp- and regional-level when drought forces them to aggregate at permanent water holes (Lee Reference Lee1972). In 32 present-day foraging societies, Hill et al. (Reference Hill, Walker, Božičević, Eder, Headland, Hewlett, Hurtado, Marlowe, Wiessner and Wood2011) found evidence of frequent inter-band relations, low group genetic relatedness of co-residents within a band, and substantial interaction networks consisting of unrelated individuals, all structured by institutions that differ from society to society. Even “acephalous” societies (those lacking formal leadership roles) use egalitarian institutions to create societies with quite large-scale activities (Mathew & Boyd Reference Mathew and Boyd2011). See section 5.1.
How similar modern foragers are to Pleistocene foragers is difficult to say with certainty. However, a reasonable assumption is that Late Pleistocene foragers exhibited a range of institutional complexity comparable to modern-day foragers. Archaeological data suggest that during the Middle Paleolithic to Upper Paleolithic transition, about 45 k.y.a, coherent cultural groups with stylistically different technology were moving about the Western Eurasian landscape (Tostevin Reference Tostevin2013). African evidence suggests that human use of symbolic markers goes back to about 164 k.y.a (Marean Reference Marean2010). CGS could well have been an active process as far back as symbolic marking is evident.
The strategies used to formulate tribal institutions are widely applied to modern organizations such as churches, villages, neighborhoods, clubs, business organizations, and bureaucracies. For example, class and regional variations are usually marked by linguistic differences (Labov Reference Labov2001). These institutional differences among groups in turn facilitate CGS via any of the three processes identified in section 2.1. The increase in institutional size and complexity in many societies in the Holocene is striking, but the evidence does suggest that the organization of anonymous societies using institutions has roots in the Pleistocene.
A theoretically significant set of institutions are those that regulate family life and small-scale partnerships. Mathew et al. (Reference Mathew, Boyd, Van Veelen, Richerson and Christiansen2013) note that humans routinely engage in a broader set of small-scale cooperative activities than do our primate relatives, and they argue that institutionalized third-party rewards and punishments often regulate such activities. Marriage and kinship are examples. Almost all cultures have one or a small number of standard, legitimate marriage contracts. Mating according to such a contract confers social benefits of the cooperative breeding system on the couple, and mating outside it is often significantly sanctioned. Kinship institutions specify the responsibilities of family members toward each other. Dispute resolution is an example of the institutionalization of small-scale reciprocity. Many societies have such institutions of varying degrees of formality (Nader Reference Nader1969). If human cooperation could rest entirely on innate adaptations for kin cooperation and reciprocity, then why should these intimate relationships require institutional support? More likely, institutional support is required if people are to enjoy the benefits of highly cooperative interactions even in small social units. Marriage is a credible signal of commitment, for example, because of institutionalized third-party enforcement. Large groups will benefit from maintaining institutions that encourage their members to cooperate at the level of small groups or families to the extent cooperation at such scales is important economically and to the quality of child rearing. For the case of marriage, see Henrich et al. (Reference Henrich, Boyd and Richerson2012b); for dispute resolution, see Gambetta (Reference Gambetta1993).
5. Social groups in competition
Here we consider examples of group competition in modern humans through the three processes outlined in section 2.1: selective borrowing between groups, migration of individuals between groups, and selective extinction and proliferation of groups. This evidence suggests that CGS occurs wherever significant cultural variation exists between neighboring and competing groups.
5.1. Warfare
The two conditions needed for warfare to cause CGS are (1) that variation in cultural traits between groups influences variation in success in warfare, and (2) that variation in success in warfare influences the spread of cultural traits. Losers of military competitions are sometimes killed outright and are sometimes voluntarily or involuntarily incorporated into the victorious society. Success and failure are often due to variation in the competing societies' technology, norms, and institutions. Losers or potential losers will also often attempt to emulate the technology and institutions of stronger societies.
Humans can make war because we solve collective action problems at large scales. Combatants experience the hazards of war, but the benefits of victory are widely shared, even by those who avoid the risks. Human societies that successfully suppress and sanction within-group violence, can still suffer high mortality from intergroup conflict. For instance, in the Turkana, a tribe of about one million people living in northern Kenya, 1% of mortality among Turkana men is from violence among Turkana, whereas 50% is from inter-ethnic raids (Mathew & Boyd Reference Mathew and Boyd2011).
Formal models indicate that genetic group selection can only support warfare in groups of, at most, a few dozen individuals (Choi & Bowles Reference Choi and Bowles2007). However, lethal intergroup violence occurs between large human groups within which genetic relatedness is low and individuals do not know each other personally. For example, one WWII engagement alone, the Battle of Stalingrad, involved more than 1.7 million soldiers and produced over two million military and civilian casualties (Glantz & House Reference Glantz and House1998). CGS can favor the evolution of warfare between cultural groups comprising hundreds to millions of mostly genetically unrelated individuals.
Even in politically un-centralized societies, CGS on simple institutions can support large-scale conflicts. The Turkana frequently raid neighboring ethnic groups for cattle (Mathew & Boyd Reference Mathew and Boyd2011). The scale of such raids can be very large despite the absence of hierarchical institutions. The average raid includes 315 men, with some raids including up to a thousand. Most participants are not close kin, and participants come from a wide swathe of Turkana society, including multiple sub-territories. This means that reciprocity based on warriors' reputations cannot by itself maintain cooperation in the raiding party. Instead, the Turkana have cultural norms that discourage cowardice, norms that are enforced by peers who sanction violators in their local group. In vignette studies, Turkana raiders had very positive attitudes toward warriors who raided animals from another ethnic group and very negative attitudes toward those who raided another Turkana territory, even though the respondents' own territory would benefit from the spoils in both situations. This pattern strongly supports CGS operating among the Turkana and neighboring groups having similar patterns of ethnolinguistic-scale cattle raiding. The common ethnic identity of Turkana combined with effective norms and institutions are the proximate cultural mechanisms that make large-scale raiding possible.
Archaeological and ethnographic data suggest that warfare occurred in a sufficiently wide range of societies for long enough (Gat Reference Gat2008) to be a gene-culture coevolutionary force. Warfare occurs in societies pre-dating agriculture and sedentary living (Keeley Reference Keeley1997). Nevertheless, the direct evidence for warfare in the Pleistocene is very limited, and doubts about war's importance before the Holocene persist (Guthrie Reference Guthrie2005).
As a consequence of warfare, the cultural traits of the successful groups often spread and the cultural traits of defeated groups and polities often decline. Similarly, groups with institutions that more effectively foster cooperation are most likely to be the victor (Turchin Reference Turchin2003; Reference Turchin2006). Victorious groups often expand their borders and absorb conquered peoples. The Mongols (Turchin Reference Turchin2006) and Nuer (Kelly Reference Kelly1985) are examples of militarily successful societies that absorbed defeated groups. Absorption may be passive, in which defeated people adopt the identity of the dominant group through marriage and/or migration, or it can be coercive. Slaves captured by the Comanche, and their subsequent offspring, became Comanche themselves after one or two generations (Hämäläinen Reference Hämäläinen2008). Cultural systems associated with Islam and Christianity have also spread in part through military conquests, facilitated by coercive as well as voluntary conversion of peoples dominated by the victors. Soltis et al. (Reference Soltis, Boyd and Richerson1995) estimated group extinction rates from the differential group extinction and conversion of losers in a sample of societies in Papua New Guinea. They found that the estimated extinction rates would allow a group beneficial cultural trait to spread to most of a meta-population in 500–1,000 years. This is consistent with the gradual increase in the complexity and size of societies over the last 10 millennia.
Variation in economic systems and political institutions between societies also affects military success. Agriculturalists find it easier to stockpile resources to feed standing armies than herders (Turchin Reference Turchin2006). Tilly (Reference Tilly1992) argues that sovereign territorial states out-competed both empires and feudal societies because they were better able to raise capital for military campaigns.
Over the past 200 years, democracies have been replacing autocracies. Part of the reason may be because democracies have been more likely to win wars (Reiter & Stam Reference Reiter and Stam2002). Democratic leaders seem to be better at picking the wars they fight, and they are more politically accountable when wars are lost. Soldiers from democratic nations also seem to fight more effectively than soldiers from non-democratic nations, even after controlling for wealth and other measures of power, perhaps because they more consistently report that they find their government's war policies legitimate (Reiter & Stam Reference Reiter and Stam2002).
To summarize, warfare clearly demonstrates the requirements for CGS. Cultural traits that affect military success are highly variable, and success in war tends to spread such traits by all three forms of CGS.
5.2. Competition between economic organizations
Competition between human organizations is certainly not restricted to violent conflict. Economic competition is an important and typically peaceful form of CGS. Economic CGS has a long history (Curtin Reference Curtin1984) and is very important in modern societies. It is partly responsible for ongoing rapid evolution in the domains of organizational rules, structure, business practices, and technology.
Economists have noted that competition among firms creates a Darwinian dynamic (Nelson & Winter Reference Nelson and Winter1982; Schumpeter et al. Reference Schumpeter, Opie and Elliott1934; Veblen Reference Veblen1898). Well-run firms profit and survive, while poorly run firms with losses expire. This selection of firms in the market was part of what Schumpeter et al. (Reference Schumpeter, Opie and Elliott1934) called “creative destruction.” Some of the success and failure of individual firms is caused by the structure, management style, institutions, and other aspects of the culture of the firm itself. Business organizations have been a focus of social identity research and research on the impact of social identity on organization performance (Ashforth et al. Reference Ashforth, Harrison and Corley2008). New employees acquire the firm's culture through formal and informal socialization and incorporate the firm's culture as part of their social identity. When those cultural differences influence a firm's success, CGS will occur. Nelson and Winter's (Reference Nelson and Winter1982) highly cited classic argues that much of the important know-how influencing firm success is tacit, embedded in the skills of individuals and informal routines of interaction. This information is hard to acquire by borrowing from successful firms and even selective hiring. Hence, cultural differences between firms tend to persist, and differential expansion and contraction of firms is very active despite attempts to acquire the practices of more successful firms.
Economists routinely study all three mechanisms of CGS in competitive market systems. The differential proliferation (market entry and split) and extinction (market exit and bankruptcy) of firms influences the future population of firms and frequencies of business practices. Empirical studies of selection on economic organizations owe much to the pioneering work of Hannan and Freeman (Reference Hannan and Freeman1993), which has attracted thousands of citations. An example is the evolution of the synthetic dye industry in the 19th century (Murmann Reference Murmann2003). The earliest synthetic dye firms were British, but German firms lobbied provincial governments to support research chemists in their universities. The students of the chemists often found jobs in German dye companies, leading to the development of the first industrial research and development (R&D) laboratories. As a result, German companies were able to out-innovate their British counterparts, and by 1900 they dominated the global market. Inspired by successful large-scale R&D during WWII, British and American governments, universities, and firms started to copy German R&D institutions on a large scale (Murmann Reference Murmann2003).
The natural selection of firms occurs when corporate cultures do not match the needs of the external environment (e.g., McGovern Reference McGovern2007). Valuable information also flows between firms through selective imitation and best-practice adoption (e.g., Bolton Reference Bolton1993; Haunschild Reference Haunschild1993) as well as patent licensing and corporate espionage. Information also migrates with employees through processes such as talent raiding (Gardner Reference Gardner2003) and spin-off companies. Klepper and Sleeper (Reference Klepper and Sleeper2005) showed that, in the laser industry, spin-off companies do indeed inherit the capacities of their parent organizations.
Substantial quantitative evidence for CGS within economic systems comes from research on competition between technology firms. For example, Derfus et al. (Reference Derfus, Maggitti, Grimm and Smith2008) observed that the market for hard disk drives created a headlong competitive race to stay near the forefront of technology; failure to keep up led to bankruptcy. Such competition led to various market-level outcomes, such as leapfrogging, in which organizations competing from a lagging position were more responsive to competition than better-positioned competitors (Barnett & Pontikes Reference Barnett and Pontikes2008). A history of competitive success tends to create a tendency for increased mistakes. Higher failure and exit rates have been observed in companies with less competitive experience, such as monopolists (Barnett & McKendrick Reference Barnett and McKendrick2004). Murmann (Reference Murmann2003) found that the originally dominant British dye firms suffered through their lack of exposure to competition. A higher rate of market entry and exit in the German dye industry led to a greater exploration of different business practices, resulting in greater variation and stronger selection. The success of Silicon Valley derives from Valley-wide “ecosystemic” institutions that accelerate all three forms of CGS (Hwang & Horowitt Reference Hwang and Horowitt2012). For example, new firms form easily in order to exploit promising ideas, and investors, entrepreneurs, and founding employees expect that most will fail. An efficient system of gossip ensures a general awareness of individuals' reputation, reducing the risk that the high mobility generated by rapid firm turnover will be exploited by unscrupulous individuals.
Modern nations intentionally regulate and promote CGS in the economic domain (Motta Reference Motta2004). Regulators force businesses to compete through anti-monopoly legislation. Patent systems attempt to induce the socially optimal amount of R&D. During the patent period, firms can recover large R&D costs, but eventually everyone can use the off-patent innovation. Collusion and anti-poaching agreements between companies are outlawed in order to guarantee competition. International free-trade agreements attempt to limit national protection of domestic companies at the expense of foreign competitors. Therefore, the mechanisms of CGS are not just present in modern economies, but are the deliberate focus of institutional design at the national and international level. In general terms, these policies, at least in principle, benefit ordinary citizens by tailoring economic group selection to meet common social goals. These national regulations may themselves constitute another level of organizational selection above the firm, in which governments, industries, and other domestic organizations are selected over time via differential success, learning, and migration between countries. The interplay between CGS at the level of the firm and the level of the nation is the subject of much national and international political discussion.
5.3. Religious competition
The action of CGS on variation in religious institutions has been observed in a number of historical and contemporary populations, often resulting in the expansion of one cultural group relative to others. Religious ethical systems are often integrated into the basic norms and institutions of a society. It is common for different religions to have different birth or death rates, and they often compete for converts.
Scholars have suggested many examples of the role of religion in competition between groups. The advent of “Axial Age” universalistic religions around 500 B.C.E. created overarching ethical systems. They introduced the innovation of doctrinal mode rituals that could unite formerly disparate cultures and thus favored expansion of empires to units large enough to resist Central Asian invaders, who had perfected horse archery (Turchin Reference Turchin2009; Whitehouse Reference Whitehouse2002). Christians in the Roman Empire were more likely to practice mutual aid and economic cooperation than non-Christians (Stark Reference Stark1997). Christian cooperators positively assorted through association in spiritual communities. Non-cooperators were punished with ostracism. This combination led to greater success in dealing with resource scarcity and disease and thus to higher survival, reproduction, and conversion rates that eventually led to Christianity being the state religion of the Empire and its successor polities. Early Islam promoted strict sacred norms of fairness and unity among co-religionists, leading to its remarkably rapid expansion. Reduced small-scale tribal warfare, increased external defense, and the creation of institutions that greatly facilitated long distance trade facilitated material success, intergroup military success, and large conversion rates (Ibrahim Reference Ibrahim1990).
Recent history suggests that competition between groups based on variation of religious belief and ritual practice continues today. An analysis of demographic data from more than 800 communes in the United States over a 200-year period revealed a strong correlation between costly ritual displays and the longevity of the commune (Sosis & Bressler Reference Sosis and Bressler2003). Finke and Stark's (Reference Finke and Stark1992) study of U.S. religious history documented an overall increase in churchgoing since independence, as well as large shifts among denominations. Hout et al. (Reference Hout, Greeley and Wilde2001) conducted a pioneering quantitative micro-evolutionary study of the relative expansion and contraction of Protestant churches in the United States during the 20th century. They found that variation in birth-rates rather than conversion rates mainly drove the changes. Pronatal norms and strong cultural isolation are leading to rapid demographic expansion of the more traditional or “Old Order” Anabaptists (Kraybill & Bowman Reference Kraybill and Bowman2001) relative to almost all other North American subcultures. The Anabaptist Hutterites are North America's most successful communal movement. They have very high population growth rates resulting from strongly pronatal norms and low apostasy, arguably caused by their strongly isolationist practices (Sato et al. Reference Sato, Nonaka, Miura and Peter1994). The number of Evangelical Protestant churches increased relative to that of Roman Catholic churches in Latin America during the 20th century, mainly due to their stronger emphasis on missionary work (Stoll Reference Stoll1990).
Not surprisingly, given the breadth of evidence, religious variation is frequently cited as one of the most important examples of CGS (e.g., Wilson Reference Wilson2002).
6. Evidence for group-level functionality
If CGS has operated on human cultures directly, and indirectly on human genes via gene-culture coevolution, then we would expect to find clear examples of design for group function. Arguments for group selection in humans have often stressed the evidence from design (e.g., Wilson Reference Wilson2002), whereas we have so far stressed process-level evidence. However, the evidence from design is also important and compelling. We discuss language, social control, and religion as examples.
6.1. Language
Language reflects the operation of CGS in three different ways. First, language is necessary for sophisticated norms and institutions to function. Second, language only functions if there is a considerable degree of trust among speakers, who comprise groups far larger than families. Third, language differences are among the most important markers of group boundaries, and limited linguistic communication makes the diffusion of institutions between societies more difficult.
Humans use language to formulate and transmit social norms and to operate institutional systems of reward and punishment that stabilize these norms. Maynard Smith and Szathmáry (Reference Maynard Smith and Szathmáry1995) nominated language as the key innovation leading to human societies arriving at a new level of biological organization. Smith (Reference Smith2010) reviews the ample evidence that language plays an important role in solving collective action problems. Norms and institutions are articulated publicly so that individuals can understand them. Individuals who appear not to understand them are typically subjected to verbal reprimands. Institutional change is advocated and discussed in speeches and codified by verbal consensus. In complex societies, written records evolved to make possible quite intricate institutions. Even the most selfish political operators usually take care to frame their public arguments to give the impression that they are furthering the general welfare.
Much variation in norms and institutions is deeply embedded in the vocabulary and syntax of languages (Wierzbicka Reference Wierzbicka1992). Such linguistically encoded information is often tacit knowledge that is shared by speakers but diffuses with difficulty to other languages. Language differences thus help preserve the between-group variation that CGS acts on. Expanding language families often mark the expansion of a complex of technical and social innovations; the Indo-European, Bantu, Tupi, and Austronesian families are examples, as we have noted in section 3.2.
The evolution of the capacity for languages that can serve cooperative functions is a considerable puzzle, because the existence of human language presupposes cooperation (Hurford Reference Hurford2007). Communication systems in animals are largely restricted to costly signals or situations where deceptive signals can be detected and punished (Lachmann et al. Reference Lachmann, Szamado and Bergstrom2001). In such systems, very little information is transmitted and syntactic complexity is quite limited (Hurford Reference Hurford2007). Unless listeners are able to trust speakers more often than not, a complex symbolic language would be useless. If most speech acts were self-serving and/or deceptive, listeners would not bother to listen. Thus, at least a modicum of trust and cooperation seems necessary before a low-cost high-fidelity communication system could begin to evolve. Scholars often assume that language is the premier mechanism for cultural transmission. If so, how could culture itself, much less CGS, ever have gotten off the ground?
How could CGS have existed before language? A plausible answer is that culture was already tolerably complex before the capacity for language began to evolve. Stone tool manufacture may go back three million years, and edge-wear evidence suggests that early stone tools were used to make a toolkit of perishable materials such as wood and hides. Merlin Donald (Reference Donald1991) hypothesizes that Homo erectus and related Lower and Middle Pleistocene hominins were talented mimics, and that mimesis served as the evolutionary basis on which language could subsequently evolve. Donald reviews evidence that 19th-century deaf-mutes could learn complex manual skills by imitation alone. They also acquired basic social norms and integrated themselves into complex institutions.Dances and similar motor rituals are among the things that pre-linguistic but highly mimetic humans could likely acquire. Such rituals today have powerful group-bonding effects and are therefore commonly incorporated into religious and secular patriotic celebrations (see sect. 6.3). Thus, a scenario in which the capacity for complex trustworthy language evolved by CGS and culture-led gene-culture coevolution (see sect. 2.2), is plausible. All the recent evolutionary theories of the capacity for language, beginning with Pinker and Bloom's (Reference Pinker and Bloom1990) pioneering article, are culture-led gene-culture coevolutionary hypotheses. Theories differ mainly in the extent to which genes have responded to the coevolutionary pressure with language-specific cognitive structures.
Language diversity suggests that language is an important component of CGS. The stunning diversity of languages shows that differences between languages readily evolve (Nettle Reference Nettle1999). Why? Following the arguments for the efficiency of having innate, information-rich cognitive modules (Tooby & Cosmides Reference Tooby, Cosmides, Barkow, Cosmides and Tooby1992), having tight constraints on languages ought to make them easier and quicker to learn both for children and for people wanting to be able to speak multiple languages. It is easy to imagine how tight, innate constraints might affect language (Sampson Reference Sampson2005), but no evidence of such constraints has been observed. Languages are highly variable in every element that has been studied closely (Newmeyer Reference Newmeyer2004). It might be that language differences, like other symbolic differences, evolve to limit communication between groups (see sect. 2.1). Sociolinguists find that small but socially significant dialect differences emerge rapidly along social fault lines, implicating the active evolution of symbolic marking rather than, or in addition to, drift-like differentiation of languages (Labov Reference Labov2001). Leaders of dialect evolution are typically individuals with high local prestige. Social identity theorists have shown leader charisma to be essentially a recognition by followers that a leader is prototypical of the in-group (Steffens et al. Reference Steffens, Haslam and Reicher2014). The ensuing emotional identification of followers with their leaders probably leads to followers adopting the speech variants of such leaders. The evolution of symbolic boundaries to communication will help protect group-level cultural variation from the homogenizing effects of migration, and thus favor the operation of CGS.
Linguistic variation is extremely important in maintaining social structure and influencing social judgments, as we saw in the case of children's early learning biases (sect. 4.2). In small-scale societies, language differences generally mark the boundaries of groups that conflict (Nettle Reference Nettle1999). It is as if CGS favors not being able to understand those whom you likely cannot trust. Even at the scale of businesses and similar groups whose members speak the same language, group-specific jargon often distinguishes insiders from outsiders. Thus, an innate capacity for the rapid cultural evolution of linguistic and other symbolic differences plausibly coevolved with group selection on cultural variation (see sect. 2.2). Language variation helps maintain between-group variation in other cultural domains, contributing to CGS, but it is also likely to be a product of it.
6.2. Crime, deviance, and punishment
Violent or peaceful competition between organizations depends upon the preservation of between-organization differences, and to compete successfully, an organization must solve dilemmas of cooperation among its members (Wilson Reference Wilson2002). CGS will tend to favor societies that produce public goods like defense, transportation infrastructure, and a fair legal system. One of the most important public goods produced in human societies are institutions of order and justice that protect individuals from predators and public goods from free-riders. All successful small-scale common property systems have institutions for monitoring and punishing defectors (Ostrom Reference Ostrom1990). In contemporary societies, measures of culturally inherited trust are correlated with economic performance (Algan & Cahuc Reference Algan and Cahuc2010). Theoretical models (e.g., Boyd et al. Reference Boyd, Gintis, Bowles and Richerson2003) and experiments (Fehr & Gachter Reference Fehr and Gächter2002) suggest that moralistic punishment can be a very effective mechanism for promoting cooperation. Thus, we expect that the quality of institutions regulating crime and deviance will play a significant role in the performance of human organizations. In experimental settings, people readily migrate from model societies that do not have sanctioning institutions to join ones that do (Gürerk et al. Reference Gürerk, Irlenbusch and Rockenbach2006). In the coevolutionary long run, social institutions of monitoring, reward, and punishment were probably the main factors exerting negative selection on genes that promote antisocial behavior and positive selection on genes that lead to prosocial dispositions (see sect. 2.2).
Punishers do not even need to be cooperative themselves to furnish some of the benefits of cooperation (Eldakar & Wilson Reference Eldakar and Wilson2008), but many experiments have detected altruistic punishment (Fehr & Gachter Reference Fehr and Gächter2002). An important theme of our argument is that the question of altruism as a proximate mechanism is not central to the issue of CGS. CGS can favor reputational and other motivational systems that alter individual incentives, thereby reducing or eliminating the need for altruism (Fehr & Henrich Reference Fehr, Henrich and Hammerstein2003; Panchanathan & Boyd Reference Panchanathan and Boyd2004). Altruistic behaviors have been the focus of models and experiments because they are the hardest ones to account for, not because they are empirically the most important. Careful experimental attempts to expose covert self-interested motives have found support for an “empathy altruism” hypothesis (Batson Reference Batson2011). In experimental treatments designed to induce research participants to empathize with someone who is going to suffer a harm, many people will volunteer to help the victim at a cost to themselves. Batson's program of experiments was designed to show that various egoistic hypotheses could not entirely explain these results. Models suggest that the evolution of altruistic punishment is likely to be frequency dependent (Boyd & Richerson Reference Boyd and Richerson1992b). If punishment is sufficiently costly to the punished and cheap to the punisher, a relatively few punishers suffice to encourage reluctant cooperators and would-be defectors to cooperate. Experimental studies using economic games support the findings of models. They find much individual variation both in propensities to cooperate and to punish non-cooperators (Henrich et al. Reference Henrich, Ensminger, McElreath, Barr, Barrett, Bolyanatz, Cardenas, Gurven, Gwako, Henrich, Lesorogol, Marlowe, Tracer and Ziker2010a; Kurzban & Houser Reference Kurzban and Houser2005). Only a minority of participants act altruistically. We are inclined to think that evolution has built an element of genuine empathy and true altruism into our social psychology because monitoring and punishment are costly to punishers and to punishees, but many people obey norms mainly because deviance is punished.
Historical and ethnographic data suggest that variation in the effectiveness of controlling self-seeking behavior is related to all three forms of group selection. For example, Turchin and Nefedov (Reference Turchin and Nefedov2009) studied dynastic cycles in agrarian societies. Elite groups do provide services in such societies, but they have a tendency to grow and become increasingly corrupt over time. Corrupt elites tend to be replaced by competitors who provide better governance. Successful empires, such as those of ancient Rome and Han China, had more effective institutions for producing public goods and incorporating new populations into their expanding polities than their competitors. The increasing acceptance of the rule of law in Early Modern Europe is fundamental to that region's explosive demographic and economic growth (North et al. Reference North, Wallis and Weingast2009). In the modern world, large numbers of migrants move from countries with low norms of civic cooperation and poor rule of law to countries with more civic cooperation and better rule of law (Boyd & Richerson Reference Boyd and Richerson2009). Thus, we have ample evidence that internal regulatory systems control deviance and solve dilemmas of cooperation to varying degrees of effectiveness. Much of the last five millennia of human history can be read as a slow, halting increase in the effectiveness of such institutions (e.g., Fukuyama Reference Fukuyama2011).
6.3. Religion and ritual
Religions furnish group-boundary–defining symbols, moral justifications for institutions of social control, and rituals that celebrate and facilitate an organization's solidarity. Religions contain effective mechanisms for reducing internal variation through sacred principles and norm-enforcing rules of reward, punishment, and exclusion, which bind participants into moral communities (Atran Reference Atran2010; Graham & Haidt Reference Graham and Haidt2010; Rappaport Reference Rappaport1999). Numerous studies have found evidence for group-functional properties of religions (Atran Reference Atran2002; Atran & Norenzayan Reference Atran and Norenzayan2005; Boyer Reference Boyer2001).
The success of a number of key cultural innovations that generate well-documented associations between in-group prosociality and religious/ritual participation appear to be clear examples of group functionality (Henrich et al. Reference Henrich, Ensminger, McElreath, Barr, Barrett, Bolyanatz, Cardenas, Gurven, Gwako, Henrich, Lesorogol, Marlowe, Tracer and Ziker2010a; Norenzayan & Shariff Reference Norenzayan and Shariff2008). Belief in moralizing high gods is associated with cooperative altruism (Johnson Reference Johnson2005; Shariff et al. Reference Shariff, Norenzayan, Henrich, Schaller, Norenzayan, Heine, Yamagishi and Kameda2009) and willingness to punish non-cooperators (McKay et al. Reference McKay, Efferson, Whitehouse and Fehr2011). Ethnographic, historical, and experimental evidence suggest that synchronized dance and rhythmic ritual facilitate coordination and bonding of groups, and relieve low-level in-group conflict (Kirschner & Tomasello Reference Kirschner and Tomasello2010; McNeill Reference McNeill1995; Rappaport Reference Rappaport1999). Theoretical arguments (Iannaccone Reference Iannaccone1992), supported by quantitative observations (Sosis & Bressler Reference Sosis and Bressler2003; Sosis & Ruffle Reference Sosis and Ruffle2003), suggest that costly signals of group membership, such as hard-to-perform rituals and other sacrifices, help groups weed out free-riders and foster in-group altruism. A common characteristic of religions is sacred rules (Durkheim Reference Durkheim1912; Rappaport Reference Rappaport1999). They supply a stable and enforceable moral code, held as inviolable by religious adherents (Atran Reference Atran2010). These often manifest as prosocial rules, including norms of fairness and norms of group defense, and also as symbolic systems that demarcate in-group membership. At the same time, of course, religions (and other ideologies), when used to create in-group solidarity, sometimes create fear and hatred of out-groups (West Reference West1941). CGS is not utopian!
Non-religious institutions also carry out rituals as part of their group culture, and they have similar effects. McNeill (Reference McNeill1995) gives evidence that the ritual of close-order drill reshaped warfare by facilitating in-group cooperation and camaraderie, leading to the higher success of groups that perform these seemingly functionally meaningless rituals.
7. Conclusion
The evidence we have outlined in this article makes the case that culture varies between human groups ranging in size from organizations within societies to ethnolinguistic tribes and nations, and even multi-ethnic empires. These groups are often symbolically marked, have more or less formal criteria for membership, and possess distinctive social institutions. This variation is transmitted to new generations by the human capacity to acquire norms and social institutions via imitation and teaching. Institutions include mechanisms of selective punishment and reward that reduce small-scale variation in behavior. Human organizations compete, often in highly organized ways, and the success and failure of groups in such competitions is often determined by institutional and other cultural differences. As Darwin (Reference Darwin1874) argued in The Descent of Man, many human groups meet the basic criteria necessary for selection to operate: heritable variation and competition, the outcome of which is based on that heritable variation. Ethnographic evidence of the prerequisites for and operation of CGS in the simplest societies suggests that CGS has operated in our lineage for a few tens of millennia, if not longer. If so, the cooperative imperatives produced by rudimentary culturally transmitted institutions may well have shaped our innate social psychology. This is reflected by the observation that young children learn norms and act on them, but chimpanzee societies have, at best, rudimentary norms. Individual-level human decisions regarding the invention of new cultural variants, the spread of variants by diffusion, and the choices of group affiliation are likely to be strongly colored by moral intuitions like sympathy and patriotism. The current direct evidence for culture-led gene-culture coevolution is only strong for genes that do not directly affect behavior, but evidence regarding behavioral traits is likely to improve greatly in the next decade.
We submit that the evidence sketched here is more than sufficient to justify taking the CGS hypothesis seriously as a basic explanation for our species' highly unusual ability to create large societies with widespread cooperation between non-relatives. We do not see how any of the alternatives to CGS can easily account for the institutionalized cooperation that characterizes human societies. We certainly do not claim that a large body of studies has met the gold standard of accurately estimating the strength of any of the three CGS processes we have discussed. Nor have we estimated the strength of culture-based social selection on human genes. These projects are in their infancy. We have pointed to excellent pioneering studies showing that a proper quantitative investigation of cultural evolution and gene-culture coevolution is possible (e.g., Bouckaert et al. Reference Bouckaert, Lemey, Dunn, Greenhill, Alekseyenko, Drummond, Gray, Suchard and Atkinson2012; Hout et al. Reference Hout, Greeley and Wilde2001; Itan et al. Reference Itan, Powell, Beaumont, Burger and Thomas2009). We need an empirical game theory that estimates costs and benefits generated by people playing various strategies in institutional games. Do individuals bear costs or neglect to take benefits that accrue to them as individuals in order to benefit their social groups? How large are such benefits and costs? Are organizations with many individuals who are willing to bear costs or forego benefits really more likely to win contests with other groups, or attract imitators and immigrants? How does rewarding individuals with prestige or symbolically with medals and prizes affect their genetic fitness or cultural success? The bad news is that the games people play are very complex, and many of those most relevant to human evolution took place many generations ago. The good news is that evolutionary biologists have struggled fairly successfully with similar complexities in organic evolution (Endler Reference Endler1986).
Appendix
Table 1. Description of data with corresponding FST values plotted in Figure 1.






Target article
Cultural group selection plays an essential role in explaining human cooperation: A sketch of the evidence
Related commentaries (27)
A framework for modeling human evolution
Clarifying the time frame and units of selection in the cultural group selection hypothesis
Cultural differentiation does not entail group-level structure: The case for geographically explicit analysis
Cultural evolution need not imply group selection
Cultural group selection in the light of the selection of extended behavioral patterns
Cultural group selection is plausible, but the predictions of its hypotheses should be tested with real-world data
Does cultural group selection explain the evolution of pet-keeping?
Frozen cultural plasticity
How evolved psychological mechanisms empower cultural group selection
Human cooperation shows the distinctive signatures of adaptations to small-scale social life
Human evolutionary history and contemporary evolutionary theory provide insight when assessing cultural group selection
Intergroup competition may not be needed for shaping group cooperation and cultural group selection
Is cultural group selection enough?
Mother–infant cultural group selection
Multi-level selection, social signaling, and the evolution of human suffering gestures: The example of pain behaviors
Self-interested agents create, maintain, and modify group-functional culture
Social selection is a powerful explanation for prosociality
Societal threat as a moderator of cultural group selection
Testing the cultural group selection hypothesis in Northern Ghana and Oaxaca
The burden of proof for a cultural group selection account
The cooperative breeding perspective helps in pinning down when uniquely human evolutionary processes are necessary
The disunity of cultural group selection
The empirical evidence that does not support cultural group selection models for the evolution of human cooperation
The role of cultural group selection in explaining human cooperation is a hard case to prove
The selective social learner as an agent of cultural group selection
The sketch is blank: No evidence for an explanatory role for cultural group selection
When is the spread of a cultural trait due to cultural group selection? The case of religious syncretism
Author response
Cultural group selection follows Darwin's classic syllogism for the operation of selection