Introduction
To paraphrase the Wikipedia definition, behavior is the range of actions made by organisms; it is the “response of the organism to various stimuli or inputs, whether internal or external, conscious or subconscious, overt or covert, and voluntary or involuntary.” This is certainly a topic that should interest movement disorder specialists. However, there is another subspeciality field of neurology called behavioral neurology, which deals mainly with cognitive function such as memory and language. Movement disorders are situated in between neuromuscular and behavioral neurology. The field of movement disorders began with a focus on disorders stemming from the basal ganglia, not muscle or cortex, but as the field has matured it has expanded on both ends, including added interest now in behavior as we try to understand more aspects of our complex patients.
Basic Principles
Movement Generation
Movement occurs as a result of muscle contraction, and muscle is controlled by the alpha motoneurons in the spinal cord. Alpha motoneurons are influenced by both segmental and suprasegmental input, with the most important suprasegmental signals coming via the corticospinal and reticulospinal tracts. It is likely that the corticospinal tract conveys the most important information, with the reticulospinal system generally playing a supportive role. The corticospinal tract originates most importantly from the primary motor cortex (M1), but has contributions from some premotor areas as well. Input to the corticospinal tract from sensory cortex may largely modulate sensory input. M1 itself receives input from cortical and subcortical structures. Subcortically, the two major systems are the basal ganglia and the cerebellum. Both derive much of their input from the cortex and send processed information back to the cortex via the thalamus. Although these two systems had been thought to be largely separate, newer evidence has shown clear connections between them [Reference Bostan and Strick1]. In general, the basal ganglia system supports features of movement concerning which movements to make and the magnitude of contraction, while the cerebellar system supports features of movement dealing with the detailed timing and coordination of the different body parts (see also Chapters 2 and 3) [Reference Fahn, Jankovic and Hallett2].
M1 also receives much cortico-cortical input coming from the entire cortical mantle (Figure 1.1). In general terms, the posterior part of the brain is the sensory portion receiving visual, somatosensory, and auditory information. This information is integrated in multisensory regions in the parietal lobe and is the source of external triggering of movement. The parietal to premotor pathways have been the object of intense study in recent decades, and highly specific connections have been identified to which specific functions can be attached [Reference Errante, Ziccarelli, Mingolla and Fogassi3]. For example, the reach and grasp components of a reach-to-grasp movement have separate parietal premotor pathways. Again, in general, the front part of the brain receives and integrates information about the body and is considered the source of internal triggering [Reference Grafton and Volz4]. Internal input includes homeostatic drive from regions such as the hypothalamus and includes factors such as hunger and thirst. Other internal input is limbic and includes factors such as vigilance, fear, anxiety, and sex. Another critical internal input is reward, seeking of pleasure, and this appears largely mediated by mesolimbic dopaminergic function. If a past behavior produced reward, the brain wants to do it again. Facilitation of behavior by repeated reward is called operant conditioning.

Figure 1.1 Scheme for the generation of behavior. Behavior is planned under internal and external influences and then generated. Normally, people think they control behavior (“self-agency”). Self-agency requires a sense of “willing” (feedforward signal) and of “registration” (feedback signal). If “willing” precedes the behavior, the quale of agency can be generated.
Both the external and internal inputs are integrated in mesial frontal areas such as the cingulate and pre-supplementary motor area, as well as the premotor areas directly [Reference Hunt5]. Strong inputs come from the mesial frontal areas to the premotor and motor cortex. What the motor cortex will produce, as behavior, at any one time, is the consequence of the integration of all these factors. In essence, behavior is constantly influenced by all environmental (external) stimuli and multiple forces from frontal (internal) drives. The more information available about all these factors, the more behavior can be probabilistically predicted. It is ordinarily a difficult calculation, which is also influenced by neural noise. However, in some circumstances individual factors are so strong that prediction would be almost certain. If someone is hungry and is presented with a pizza, it is very likely that they would eat it. If someone had immense pleasure from cocaine and was offered it again, they might take it again even though cognitively they know that this will only make the situation worse in the long run.
Consciousness and Voluntariness
From all we know about the brain, it is constantly in action, and it appears that many different things are being processed simultaneously. For example, as was already pointed out, there are multiple, continuous, external and internal inputs. Additionally, many different thought processes can be going on simultaneously. If you are asked a question, such as someone’s name, and cannot immediately think of it, you can go on and think about something else, and, often, sooner or later, the name will “pop into your head.” This is likely the product of ongoing searching. If given a difficult decision, you might say, “I will think about it and let you know later.” Even if you do not devote considerable time thinking about it at a conscious level, you will be able to come to a decision. Much of this brain activity is unconscious. Only one thing at a time, or rarely two things, will bubble up into consciousness. And the stream of consciousness does not always flow smoothly; the topic may change quickly and not always logically from one to the next.
What is in consciousness must be in some way what is important at the time and is a result of the process called attention. Attention can be bottom up or top down. A strong external stimulus will usually bottom up into consciousness regardless of what else is going on. However, if a soldier is paying top-down attention to fighting with an enemy, he might not notice that he has been shot in the leg.
The individual elements of consciousness are called qualia, and one quale, relevant for movement disorders and other reasons, is voluntariness [Reference Hallett6]. Much of the time, it would be fair to say that persons are not thinking about whether a movement is voluntary. Things happen. For most movement, people generally think that they are the “agent” of the movement; that is, they willed the movement and it occurred. This is the sense of agency or, specifically, self-agency. Self-agency presumably requires both a sense of willing, a feedforward signal, and the sense that the willed movement occurred, a feedback signal. If willing precedes the specific action that was willed, then the quale of agency can be created (see Figure 1.1).
The sense of agency utilizes a brain network with a prominent role for the temporoparietal junction (TPJ) [Reference Nahab, Kundu and Gallea7]. Presumably this feedforward–feedback process is happening all the time but does not create a quale because it is so routine. However, if the process does not work correctly – for example, a movement occurs for which there is no feedforward signal – that might bubble up to consciousness as a surprise, generating the quale of an involuntary movement. Whether movements are voluntary or involuntary is often a concern to the patient and to the movement disorder neurologist.
Behavioral Movement Disorders
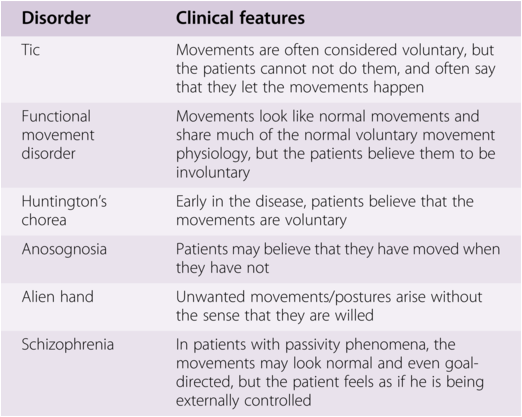
Many of the patients seen by movement disorder neurologists have some behavioral aspects and some are even “primarily” behavioral. Several features are easier to explain than others, and some examples will be given here to illuminate general as well as specific issues. These disorders overlap to some extent to the “disorders of volition” (see Table 1.1) [Reference Hallett6, Reference Kranick and Hallett8].
Table 1.1 Some disorders of volition [Reference Hallett, Wolters and Baumann37] (with permission of author and publisher)
| Disorder | Clinical features |
|---|---|
| Tic | Movements are often considered voluntary, but the patients cannot not do them, and often say that they let the movements happen |
| Functional movement disorder | Movements look like normal movements and share much of the normal voluntary movement physiology, but the patients believe them to be involuntary |
| Huntington’s chorea | Early in the disease, patients believe that the movements are voluntary |
| Anosognosia | Patients may believe that they have moved when they have not |
| Alien hand | Unwanted movements/postures arise without the sense that they are willed |
| Schizophrenia | In patients with passivity phenomena, the movements may look normal and even goal-directed, but the patient feels as if he is being externally controlled |
Behavioral Abnormalities due to Parietal-Premotor Disorders (Apraxia and Task-Specific Disorders)
There are many types of apraxia; the one most commonly recognized is ideomotor apraxia in which there appears to be the loss of a motor memory for a skilled movement [Reference Wheaton and Hallett9, Reference Park10]. The patient seems to understand what movement is to be made, there is no responsible deficit of language, and the motor machinery is in good enough shape to make the movement, but the proper sequence of actions is not generated. Bedside testing is generally done with external movements, either asking someone to make a particular movement (such as “show me how you would use a hammer”) or to mimic the examiner making a novel movement. However, such patients often cannot make the complex movement even in the natural context and might be impaired in activities of daily living. Such an apraxia, for example, is characteristic in patients with corticobasal syndrome (see Chapter 32).
Patients with focal hand dystonia commonly begin with a derangement of just a single task, such as only writing or only playing the piano. Other tasks are done normally. When attempting the task, the motor coordination falls apart and a dystonic spasm intervenes. This is referred to as task specificity. It is similar to ideomotor apraxia in that there is a failure of a learned motor program.
There is good evidence that skilled movements are stored in the brain in the parietal area or, perhaps more accurately, in parietal-premotor networks, mainly in the left hemisphere, for both right or left hand movements [Reference Bohlhalter, Hattori and Wheaton11]. Basic observations as to how movements are learned in human studies show that activity in parietal-premotor pathways increases as movements are learned. As the movements are learned to the point of automaticity, the activity declines but the connectivity in the network becomes stronger [Reference Wu, Chan and Hallett12]. In praxis movements, such as handwriting [Reference Horovitz, Gallea, Najee-Ullah and Hallett13], the activation of left-sided parietal-premotor pathways can be identified with both neuroimaging and electroencephalography (EEG) studies (see Figure 1.2).

Figure 1.2 Functional magnetic resonance imaging (fMRI) from normal subjects making transitive movements (mimicked movements that employ a tool) with right and left hands. Axial and coronal sections are shown. There are prominent activations in parietal, premotor, and supplementary motor cortex, more on the left side even with left sided movement.
So, in apraxia there appears to be a general failure of the parietal-premotor pathway. Lesions of the parietal area are common causes of the disorder, but premotor lesions can also be responsible as well as “pure disconnections” between the two areas. In writer’s cramp, we identified that the specific writing parietal-premotor connection is hypoactive [Reference Gallea, Horovitz, Ali Najee-Ullah and Hallett14], but why this is accompanied by dystonic spasm is not understood.
Behavioral Abnormalities due to Paralimbic Disorders (Tics)
Tics are on the border between voluntary and involuntary movements. If pressed, most adults who have tics say that they are voluntary movements made to reduce an inner psychic tension or a sensory feeling. When the movement is made, the person feels temporarily better, but then the tension or sensation begins rising again. However, the movements are done so automatically that ordinarily there is no sense of willing or even recognition that the tic occurred. Children with tics have a more difficult time explaining the nature of their movements but will generally say that they are involuntary. When a movement leads to a good feeling, it can be considered rewarding, and it is possible that tics are perpetuated in part due to operant conditioning. Thus, a tic could be thought of as an undesirable habit [Reference Graybiel15].
The sensory feeling that provokes the tic is called a sensory tic, and little work has been done to understand it. Patients feel that they are particularly sensitive to sensory stimuli; they become very annoyed, for example, with tags on shirts. However, the psychophysics of their sensory perception, including thresholds, is normal, suggesting a possible failure of habituation [Reference Belluscio, Jin, Watters, Lee and Hallett16].
The urge to tic is similar to normal urges such as the urge to scratch an itch or the urge to blink when trying to keep the eyelids open. fMRI studies of the urge to blink show that the anterior insula is particularly active [Reference Berman, Horovitz, Morel and Hallett17]. Using an event-related design with fMRI, it has been demonstrated that the anterior insula and the anterior cingulate are active prior to tics (see Figure 1.3) [Reference Bohlhalter, Goldfine and Matteson18]. Additionally, comparing the resting brain activity while awake, when tics are occurring, and asleep, when tics are rare, there is also increased activity of the anterior insula and cingulate [Reference Lerner, Bagic and Boudreau19]. Hence, the urge to tic may well arise in these structures.

Figure 1.3 fMRI of axial (A), coronal (B), and sagittal (C and D) views in event-related design of spontaneous tics. The upper row shows significant activations (P < 0.05, corrected for multiple comparisons) of paralimbic areas (anterior cingulate cortex and insular region bilaterally) before tic onset; these activations were much less prominent at tic onset (lower row).
There is an EEG signature called the Bereitschaftspotential or BP that can be identified in the 1.5 s or so prior to a voluntary movement [Reference Shibasaki20, Reference Shibasaki and Hallett21]. It is a slowly rising negativity that begins fairly symmetrically around the vertex, and as the movement approaches the negativity rises a bit faster and, at least for right-sided dominant hand movements, the potential peaks more over the left sensorimotor cortex. The early part of the potential, BP1, arises from the supplementary motor area (SMA) and the lateral premotor cortex, both parts of area 6. The later part of the potential, BP2, emphasizes more the premotor cortex and the primary motor cortex (M1). With tics, there is either no BP at all or just a BP2 [Reference van der Salm, Tijssen, Koelman and van Rootselaar22]. This suggests that tics originate with only minimal involvement of area 6. Perhaps the anterior insula and anterior cingulate access the motor cortex directly.
Behavioral Abnormalities due to the Loss of “Self-Agency” (Functional Movement Disorders)
Functional movement disorders come in many varieties, both positive and negative. Virtually any other movement disorder can be mimicked. The negative disorders of weakness and paralysis are common, but typically present to neurologists, for example, seeing strokes, neuromuscular disease, or multiple sclerosis. At the other extreme, paroxysmal hyperkinetic functional movements are often categorized as functional seizures (psychogenic non-epileptic seizures). The underlying etiology for such disorders is complex and multifactorial, and requires considering a biopsychosocial model, including genetic factors, stress responsivity, childhood trauma, and the current social structure [Reference Drane, Fani and Hallett23–Reference Perez, Edwards and Nielsen25].
The psychiatric etiology is most commonly considered to be conversion, where, in Freudian terms, a psychological symptom is converted to a somatic symptom. This is an unconscious process and the movement is not voluntary. Alternate etiologies are factitious and malingering, where the movement is voluntary, but the patient says that they are involuntary. At present, we have no clinical or laboratory way of separating these entities, except by secret surveillance. For the rest of this discussion, the etiology will be assumed to be conversion.
Functional movements appear to utilize brain mechanisms very close to those used by ordinarily voluntary movements [Reference Baizabal-Carvallo, Hallett and Jankovic26, Reference Hallett, Hallett, Stone and Carson27]. In functional paralysis, transcranial magnetic stimulation (TMS) of the motor cortex produces normal motor-evoked potentials (MEPs), indicating a normal motor cortex and pathway all the way to the muscle. When imagining movement of a body part, the MEPs should enhance, but in functional paralysis, the MEPs are reduced, suggesting an inhibitory influence on M1. When apparently trying to move the paretic limb in functional paralysis, fMRI shows activity changes in frontal lobe areas. Perhaps the inhibition comes from the frontal lobe.
In functional tremor, one important observation is that the tremor can be entrained by voluntary rhythmic movement of another body part. This suggests that the functional tremor generator is likely shared with the voluntary generator. Similarly, functional tremor is typically synchronous in different limbs, different from other tremors such as seen in Parkinson’s disease (PD) and essential tremor.
Functional myoclonus has an electromyography (EMG) pattern similar to quick voluntary movements in terms of EMG burst length and antagonist muscle relationships. When stimulus-induced, it behaves like a normal reaction time movement in terms of mean latency and variability of latency, unlike cortical or brainstem myoclonus where the latency is very short with little variability. Importantly, there is often a typical BP preceding the functional myoclonus, indicating preparation for movement in area 6 (see Figure 1.4).

Figure 1.4 EEGs associated with functional jerks of trunk and self-initiated movements mimicking the jerks, arranged time-locked to the EMG discharge. Fifty traces of individual EEG records are shown on the top panel (single trial) and averaged waveforms are shown on the bottom (averaged). In single trials, blue and red colors indicate surface-negative and surface-positive, respectively. Note slowly rising surface-negative potential preceding both the functional jerks and the mimicking jerks. Data recorded by Dr. Zoltan Mari in the Human Motor Control Section, NINDS.
While the origin of functional movements is obscure, in several circumstances there is fMRI evidence of a hyperactive limbic system. The passive response of the right amygdala is increased to emotional faces, and several structures in the limbic system are hyperactive even with voluntary movements [Reference Voon, Brezing and Gallea28]. Another relevant fMRI observation is that the right TPJ is hypoactive with psychogenic tremor compared to voluntarily mimicked tremor [Reference Voon, Gallea and Hattori29]. As noted earlier, the TPJ is implicated in the sense of agency, and therefore, this hypoactivity might explain the loss of self-agency for the movements. It can be speculated that the lack of the normal feedforward signal from the aberrant movement intention may explain the abnormal TPJ activity.
If stress is one of the responsible factors in functional movements, abnormal signals from the limbic system can well be understood as the prime mover. A further question that typically arises is why different persons have different types of functional movement disorders. One possible explanation for this is that the brain often mimics what it knows. If a person knows someone with a stroke, they might have paralysis. If another person has a relative with PD, they might have a tremor. Some subjects even mimic themselves. This is common with functional seizures, where many of these patients also have organic seizures. Mimicry is a major function of the brain that aids in the process of motor learning and which also may be responsible for empathy [Reference Baird, Scheffer and Wilson30]. I feel your pain. The mirror neurons, those motor neurons that show responses when seeing specific movements, as well as when making those same movements, may be the underlying substrate for the mimicry [Reference Rizzolatti and Sinigaglia31].
Behavioral Abnormalities due to Reduced Internal Triggering (Akinesia, Hypokinesia, Bradykinesia)
Slowness of movement, or bradykinesia, is one of the major features of PD [Reference Bologna, Paparella, Fasano, Hallett and Berardelli32]. In general terms, the explanation fits well into the scheme of behavior put forward here. The basal ganglia support mainly the front half of the brain, and it is the front half where internal triggering of movement is generated. Hence, patients find it difficult to initiate movement (akinesia) and make slow (bradykinetic) and small (hypokinetic) movements. Patients must compensate for this difficulty by paying more attention to movements requiring more cortical resources. Externally triggered movements are much better because these appear to require less basal ganglia support. External triggering underlies the phenomenon of paradoxical kinesia [Reference Hallett33].
Another feature of the slowness in PD is the sequence effect, where repetitive movements get gradually slower or smaller. The sequence effect is easily seen with handwriting that gradually becomes more micrographic through the sentence. Additionally, the sequence effect commonly precedes a gait-freezing episode. This feature may be specific to PD; at least, it is not common in progressive supranuclear palsy (PSP) [Reference Ling, Massey, Lees, Brown and Day34]. The pathophysiology of this behavior is not known. It is not responsive to dopamine [Reference Kang, Wasaka and Shamim35] and hence can be a significant clinical problem even when the PD is generally responsive to oral therapy.
Behavioral Abnormalities due to Abnormal Operant Conditioning (Impulse Control Disorders)
PD patients may show a variety of abnormal behaviors, which are discussed in Chapter 13 in this book. Such behaviors include pathological gambling, pathological shopping, and punding [Reference Weintraub and Mamikonyan36]. These are all repetitive activities, which, similar to other forms of addiction, the patient continues to do even though recognizing that it might not be the best thing to do. These behaviors are related to abnormal dopamine functioning and operant conditioning creating undesirable habits.