


Introduction
Conservation authorities have had a difficult time suppressing conflict between humans and elephants Loxodonta africana, a challenge that is widespread and problematic in both Africa and Asia. In Africa, in particular, the resolution of conflict has become a political challenge, as rural chiefdoms may resist mitigation efforts (Osborn & Parker, Reference Osborn and Parker2003), which could threaten conservation programmes (Hoare, Reference Hoare1999; Sitati & Walpole, Reference Sitati and Walpole2006). In the last 2 decades research has increasingly turned to investigating the causes of human–elephant conflict (Naughton-Treves, Reference Naughton-Treves1997; Hoare, Reference Hoare1999; Smith & Kasiki, Reference Smith and Kasiki2000; Parker & Osborn, Reference Parker and Osborn2001; Mosojane, Reference Mosojane2004; Sitati et al., Reference Sitati, Walpole, Smith and Leader-Williams2003, Reference Sitati, Walpole and Leader-Williams2005; Harris et al., Reference Harris, Russel, Van Aarde and Pimm2008; Jackson et al., Reference Jackson, Mosojane, Ferreira and Van Aarde2008; Graham et al., Reference Graham, Douglas-Hamilton, Adams and Lee2009). Although some studies make reference to the possible role of elephant pathways as spatial variables in human–elephant conflict (Smith & Kasiki, Reference Smith and Kasiki2000; Mosojane, Reference Mosojane2004; Chiyo et al., Reference Chiyo, Cochrane, Naughton and Basuta2005; Sitati & Walpole, Reference Sitati and Walpole2006), no research to date has investigated the spatial relationship between crop-raiding location and pathways.
Some pathways have been used for 50 years (Haynes, Reference Haynes2006), with their position representative of long-term movements and feeding patterns of elephant populations (Agnew, Reference Agnew1966). Williamson (Reference Williamson1975) recorded elephant trails following the troughs of Pleistocene dunes in Zimbabwe. In Botswana, Child (Reference Child1968) observed that seasonal movement by elephants was reflected by well-defined pathways, especially those leading to pans. Pathways or trails have been mentioned anecdotally in connection with watering holes or drinking areas (Moss, Reference Moss1988; Payne Reference Payne1998). Shannon et al. (Reference Shannon, Matthews, Page, Parker and Smith2009) found that the most heavily used elephant pathways were close to water. Crop damage caused by elephants in the Okavango Delta can be along established elephant pathways (NRP, 2006). K. Leggett (pers. comm.) confirms that desert elephants use pathways seasonally between feeding areas, as well as daily to access preferred drinking spots along the Hoanib and Hoarusib Rivers in Namibia.
In the equatorial forests of central Africa, forest elephants L. africana cyclotis play an important role in ecosystem dynamics by opening up clearings, structuring tree-species composition through seed deposition and by creating forest pathways that are subsequently used by other species (Weinbaum et al., Reference Weinbaum, Nzooh, Usongo and Laituri2007). Forest elephant trails can be tens of km long and several m wide (Blake & Inkamba-Nkulu, Reference Blake and Inkamba-Nkulu2004. It has been suggested that migrations of forest elephants may follow regular pathways rather than being random movements (Turkalo & Fay, Reference Turkalo and Fay1995). Elephant trails in the Congo connect forest clearings, which are important social gathering sites, and different sized pathways have different functions, with larger trails used for long-distance faster travel and smaller, more sinuous, pathways used for foraging and accessing resources (Van Leeuwe & Gautier-Hion, Reference Van Leeuwe and Gautier-Hion1998).
The identification of elephant pathways, and the landscape elements they connect, may offer insights into habitat requirements in a fragmented landscape. Much like seasonal corridors, pathways could facilitate daily movement, increase feeding efficiency by reducing travelling times to high-nutrient, clustered and stable food sources. Here we report on African savannah elephant L. africana africana movement along pathways, the landscape features (water, forage, refuges) connected by pathways, and the spatial correlates (distance to nearest settlement, refuge, road, protected area, river and elephant pathways) of raided field locations. Our study offers new insights into the spatial variables of human–elephant conflict and provides guidance for land-use planning initiatives and conservation managers.
Study area
The c. 190 km2 Kwandu Conservancy occupies the centre of the Caprivi Strip (Fig. 1) in north-east Namibia. Lying central to the Kavango-Zambezi Transfrontier Conservation Area, the Kwandu Conservancy and its southern conservancy neighbours comprise a priority corridor for elephant movement from Botswana's Chobe National Park into Zambia's Sioma Ngwezi National Park and Angola's Luiana Partial Reserve. The Kwandu Conservancy is situated between two protected areas: Bwabwata National Park in the west, and Caprivi State Forest in the east. The international boundary of Zambia's Sioma Ngwezi National Park and Angola's Luiana Partial Reserve forms the northern boundary. The Kwandu Conservancy is considered a human–elephant conflict hotspot (Hanks, Reference Hanks2006).

Fig. 1 The study area within southern Africa's Kavango-Zambezi Transfrontier Conservation Area (KAZA TFCA). The rectangle on the inset indicates the location of the main map in north-east Namibia.
The most densely populated conservancy in Namibia, the Kwandu Conservancy is inhabited by 4,300 people. Community-based natural resource management is practised, and elephants are protected by law. Kongola is the largest village. Villages, crop fields, schools and clinics are mostly sited adjacent to the main north–south gravel road. Cultivation is practised in the wet season (November–April), when farmers plant maize, millet and sorghum, which are harvested in April and May. The greater Kwandu Basin is marked by the presence of fossil dunes where shallow, seasonally flooded areas form in the dune troughs and ancient river valleys. The Kwandu River is the western boundary of the Kwandu Conservancy, and both people and elephants rely on the river for water in the dry season (May–October). During the wet season surface water is widely available in numerous waterholes, pans and omuramba (ancient river valleys) but by May most of the waterholes have dried up. Rainfall is variable, occurring mainly in summer months between November and April. Mean annual rainfall is 600 mm. Soils are predominantly Kalahari sands and nutrient-poor (Robertson, Reference Robertson2005). The landscape is flat, at altitudes of 930–1,100 m (Mendelsohn & Roberts, Reference Mendelsohn and Roberts1997). Broad-leafed savannah characterizes the Caprivi Strip. Mopane-Burkea and teak woodland, mixed shrubland and omuramba grassland dominate the area, with mature woodlands (Acacia spp.) occurring in the region (Mendelsohn & Roberts, Reference Mendelsohn and Roberts1997).
Methods
Fieldwork to locate and explore elephant pathways was initiated from the Caprivi State Forest border, with the assistance of local game guards. Only permanent pathways, with floors devoid of vegetation and regularly used by elephants, were selected (Blake & Inkamba-Nkulu, Reference Blake and Inkamba-Nkulu2004). Regular use of pathways by elephants was confirmed through visual observation of dung, feeding damage and elephant footprints. Pathways that did not show any of these signs were excluded from the analyses. Pathways were followed from the border to the Kwandu River in the west. To record pathway length for spatial analyses in ArcGIS v. 9.3 (ESRI, Redlands, USA), coordinates (with a global positioning system, GPS), elevation, and pathway width were logged at 100-m intervals. Land use and presence of water pans, fields, roads, crossing points and prints of other species were also noted. Elephant activity along pathways and crossing points was documented over 120 observation days during the peaks of the wet and dry seasons of 2008. Fresh spoor was recorded daily between 06.00 and 07.00. To avoid recounting old spoor, footprints were eradicated after each count, and bull and breeding-herd spoor (including that of offspring) were noted. Male and female elephant groups were distinguished by the presence or absence of footprints of elephants < 6 years old (Balasubramanian et al., Reference Balasubramanian, Bhaskaran, Swaminathan, Desai, Daniel and Datye1995; Chiyo & Cochrane, Reference Chiyo and Cochrane2005).
Ground surveys were used to register all reported crop-raiding incidents in the peak wet season months (March–April) of 2008, when crops mature. Two hundred and fifty crop fields were georeferenced. Each field was assigned a random number drawn from a uniform distribution. Fields were sorted according to random numbers and the first 100 fields were selected as a random sample. We divided fields into three categories reflecting the protocol of Integrated Rural Development and Nature Conservation, Namibia: if elephants had been present in the field, the incident was recorded as either ‘raided’ (> 25% of field damaged) or ‘non-raided, elephant presence’ (< 25% of field damaged). If no elephants had been to the field, it was categorized ‘non-raided, elephant absence’. Corner points of the fields recorded with a GPS were used to create polygons in ArcGIS and the centre of each field was determined using the centroid tool. The 100 fields were mapped and distance measures from each field centre to the nearest pathway, settlement, tar and gravel road, river, forest refuge and protected area (spatial correlates tested in previous human–elephant conflict research; e.g. Naughton-Treves, Reference Naughton-Treves1997; Hoare, Reference Hoare1999; Smith & Kasiki, Reference Smith and Kasiki2000; Sitati et al., Reference Sitati, Walpole, Smith and Leader-Williams2003, Reference Sitati, Walpole and Leader-Williams2005) were determined using ArcGis. One–way ANOVA was used to test for association between whether crop fields were raided by elephants or not, and distance to the nearest settlement, protected area, forest refuge, tarmac road, gravel road, river and pathway, and field elevation.
Results
The 12 elephant pathways connected the Kwandu River with the Caprivi State Forest and the Bwabwata National Park, crossing three land-use types: forest, agricultural land and floodplain. Pathways were used at night, allowing elephants to traverse the anthropogenic landscape in relative safety. Pathways tended to follow an east–west orientation and had low curvilinearity. They connected resource patches such as preferred drinking or crossing points on the Kwandu River (Plate 1), with waterholes, maize fields and crossing points on the main north–south road. Waterholes were found along 50% of the pathways (Table 1). However, 83% of these waterholes were dry by May (the end of the wet season). Female elephants avoided pathways in areas of medium and high settlement density (< 100 people km−2) in the south, Mendelsohn & Roberts, Reference Mendelsohn and Roberts1997), preferring the northern pathways further away from settlements (areas with < 10 people km−2). Teak woodland and shrubland constitute the major vegetation type in the forest and includes tree species important for elephant diet in the wet and dry seasons. Crop fields are a mixture of fallow and planted fields, with maize the dominant crop type. Pathways become less defined (i.e. non-linear) after entering crop fields but increasingly linear when approaching road-crossing points and when leading to a river. Crossing points at roads remained 100% consistent, with elephants always crossing at the same locations for all observations. Pathways were used by other species, including the hyaena Crocuta crocuta, hippopotamus Hippopotamus amphibius, leopard Panthera pardus, jackal Canis mesomelas, wild cat Felis libyca, duiker Sylvicapra grimmia, kudu Tragelaphus strepsiceros and bushpig Potamochoerus larvatus, especially in the north, in areas of decreased human disturbance.

Plate 1 Elephant Loxodonta africana pathways within the Kwandu Conservancy were linear, with floors devoid of vegetation (a, b, e). Pathways connected habitat and resource patches such as waterholes (c) and preferred drinking spots on the river (f) with crossing points on the Kwandu Conservancy road (d).
Table 1 Characteristics of the 12 recorded elephant Loxodonta africana pathways, with density of adjacent human settlements, pathway length, width and elevation, number of branches off a pathway, number of adjacent waterholes, whether males or both males and females used the pathway, and season in which the pathway was used.
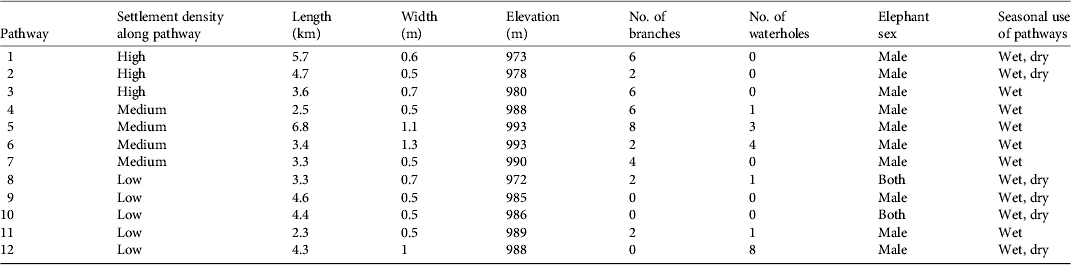
Pathway activity was significantly higher in the peak wet season months (March–April; 60 records) than in the second dry season months (September–October; eight records), with peak activity in April (Fig. 2). Data records of elephant footprints showed that pathway use was highest in the southern, high-density settlement areas and in the extreme northern low-density settlement area Monthly rainfall and elephant incident data for 2006–2008 showed a clear correlation between timing of rainfall and elephant raids, with raids occurring after above-average rainfall events (Spearman Rank correlation = 0.685). Crop raiding occurred as crops ripened towards the end of the rainy season, with a peak of incidents in April. Of the 168 recorded incidents, only 24% comprised actual raids (> 25% of field destroyed). Maize was the most frequently planted crop (n = 100), and the most affected by elephant incidents. Mature crops were raided more frequently than interim or immature crops (χ2 = 8.32, P = 0.00392). Bulls were responsible for 100% of reported incidents, and all incidents occurred at night. Bull group size increased with incident type, with larger groups responsible for raids (z = 2.91, P < 0.01). Seventy-five percent of fields raided in 2007 were targeted again in 2008 (χ2 = 11.95, P = 0.02). Distance to protected area, forest refuge, tar and gravel roads, river and elevation were insignificant as spatial variables in explaining field-raiding position. Although statistically not significant, fields lying close to settlements tended to be raided less frequently than fields at a distance from settlements. Fields close to pathways were visited by elephants more frequently and raided significantly more often than fields that lay further away (Fig. 3).
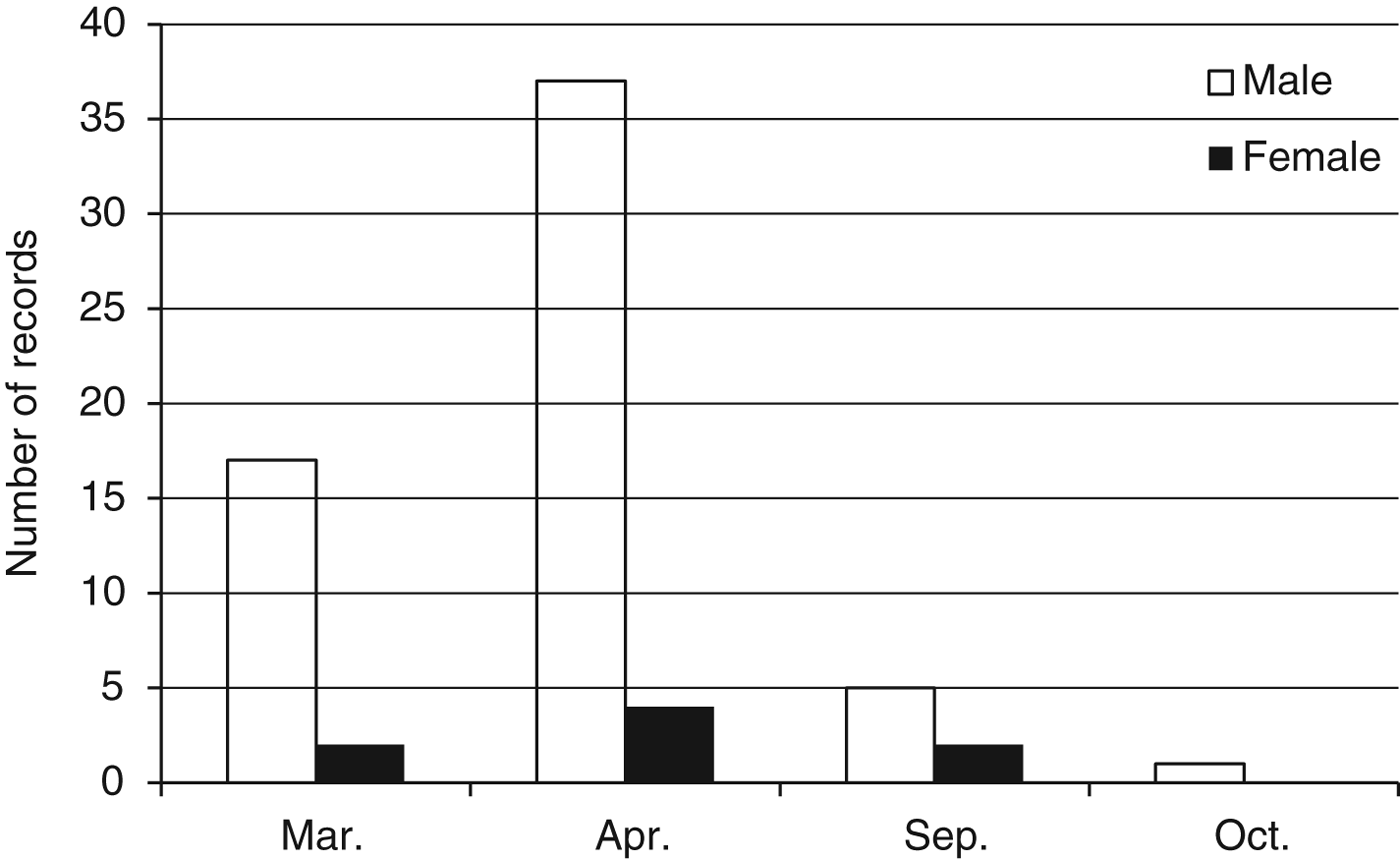
Fig. 2 Number of records of use of pathways in the wettest (March and April) and driest (September and October) months by male and female elephants.
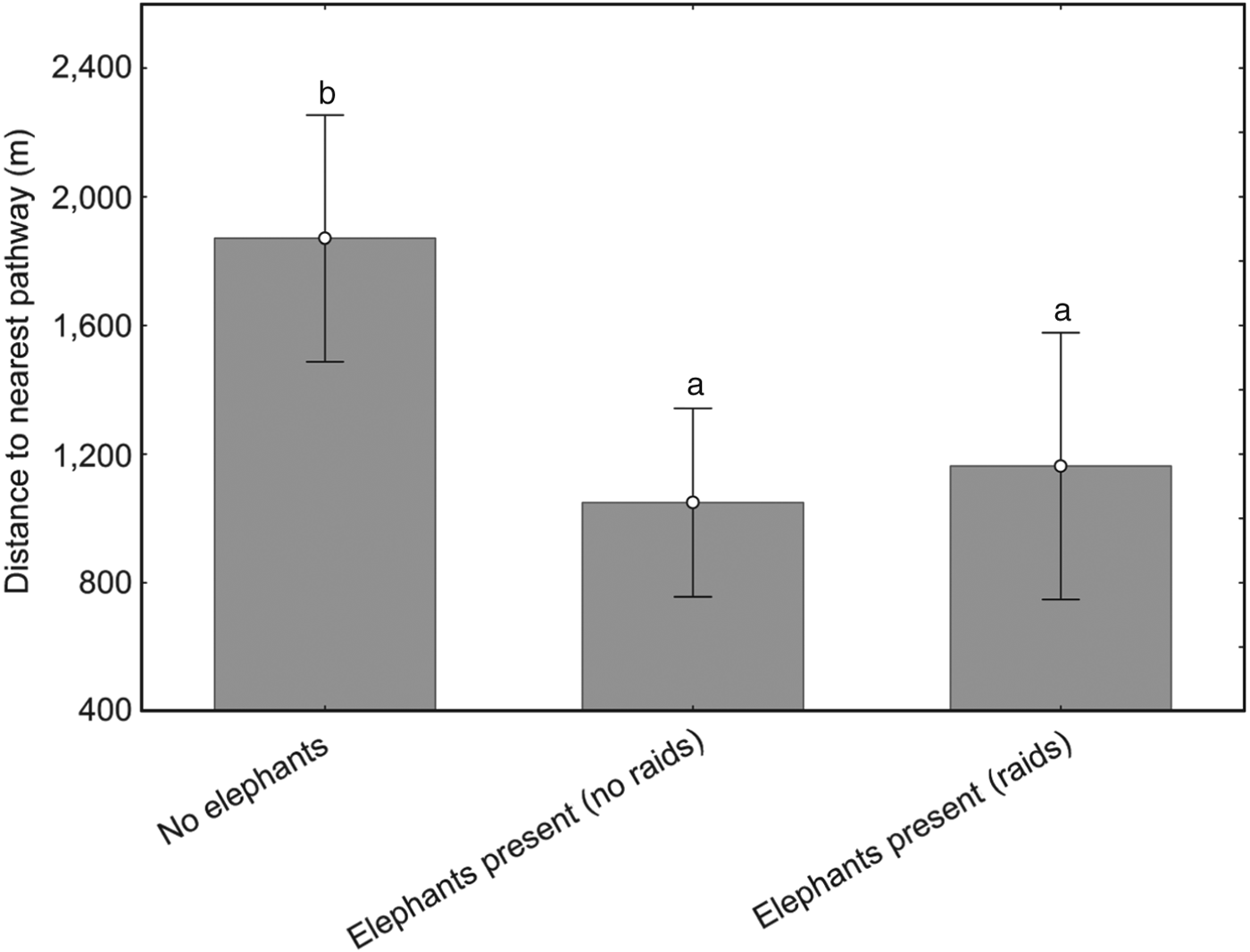
Fig. 3 Fields close to pathways were visited by elephants more frequently and raided significantly (P < 0.01) more often than fields that lay further away. Bars with different letters above are significantly different.
Table 2 One–way ANOVA testing for association between whether crop fields (n = 100) were raided by elephants or not and distance to the nearest settlement, protected area, forest refuge, tar or gravel road, river and pathway, and field elevation.

Discussion
Many species use pathways, for a host of reasons. The rock hyrax Procavia capensis uses foraging trails within its home range (Estes, Reference Estes1992). The black rhinoceros Diceros bicornis in East Africa move along the ‘same well worn paths’ when moving to and from drinking areas (Ritchie, Reference Ritchie1963). Harvester ant Pogonomyrmex spp. foragers use a trunk-route system to facilitate exploitation of patchily distributed yet stable food resources (Holldöbler & Lumsden, Reference Holldöbler and Lumsden1980); trunk trails are consistent over time, with chemical and visual cues along trails contributing to trail persistence.
Hippopotamus pathways in swamp areas are aligned with the prevailing slope and develop into channel systems that keep river channels open. Main trails are connected to lateral trails that lead to grazing areas. Pathway use has been recorded in the forest elephant (Van Leeuwe & Gautier-Hion, Reference Van Leeuwe and Gautier-Hion1998), and Shannon et al. (Reference Shannon, Matthews, Page, Parker and Smith2009) mapped an entire savannah elephant pathway network in South Africa, recording an increased density of pathways in closed woodland close to water.
Our study is the first to demonstrate the spatial significance of pathways for elephant movements and crop-raiding location. We observed that females use pathways far from settlements to access water, whereas males use pathways throughout the year and irrespective of density of human settlements. Bulls used the pathways not only in the harvesting (wet) season when crops ripen but also at the peak of the dry season, and this suggests that elephants use pathways for reasons other than crop-raiding. Pathways for savannah elephants may be least-effort routes between daily and seasonal resources such as preferred drinking and resting areas, crossing points on roads, and foraging sites. Pathway use in Kwandu Conservancy therefore appears to maximize optimal foraging by facilitating movement in a disturbed matrix and linking predictable resources.
Maximizing optimal foraging
Area-concentrated search is considered vital in patchy environments (Fortin, Reference Fortin2003), where search mode is adapted to habitat structure. Search modes can either be intensive (with low travel speeds and high path sinuosity) or extensive (with high travel speeds along linear paths). Minimizing travel distance between resources is an obvious strategy to maximize the cost-benefit balance (Noser & Byrne, Reference Noser and Byrne2007). The procurement of food at the lowest energetic cost forms the basis of optimal foraging theory. The use of least-effort routes has been described in chimpanzees Pan troglodytes verus (Normand & Boesch Reference Normand and Boesch2009), baboons Papio ursinus (Noser & Byrne, Reference Noser and Byrne2007) and buffalos Syncerus caffer (Bar David et al., Reference Bar-David, Bar-David, Cross, Ryan, Knechtel and Getz2009), with animals displaying a goal-directed approach to out-of-sight food and water sources. Pathway position could reflect preferred elephant routes to and from favoured areas (Shannon et al., Reference Shannon, Matthews, Page, Parker and Smith2009).
In Kwandu Conservancy elephants foraged randomly within homogeneous crop fields but when travelling through the anthropogenic matrix movement was directional and non-random along clearly-defined pathways. Pathways may act as least-effort routes to stable and clustered nutrient sources such as fruiting trees, crops and mineral licks. In tropical forests distribution of forest elephant trails has been linked to high-nutrient food sources such as fruit and mineral deposits (Blake & Inkambu-Nkulu, Reference Blake and Inkamba-Nkulu2004) and elephant trails link important fruiting trees (Short, Reference Short1983; White, Reference White1992). In Botswana, Child (Reference Child1968) recorded trails leading to fruiting trees such as marula Sclerocarya birrea and mugongo Schinsiophyton rautaneii. In the Kwandu Conservancy and adjacent areas there are numerous species of fruiting trees favoured by elephants, including marula, Transvaal gardenia Gardenia volkensii, camelthorn Acacia eriloba, candlepod acacia Acacia hebeclada, jackalberry Diospyrus mespiliformis and rosewood Guibourtia coleosperma. Trees fruit between November and April (Roodt, Reference Roodt1998), coinciding with the wet season, which is also the time that farmers harvest their crops. As some of these trees grow in the greater Kwandu Conservancy area, and many of the riverine areas are densely settled, human–elephant interactions are inevitable. Bulls that lose part of their home range to crop fields can become crop-raiders (Balasubramanian et al., Reference Balasubramanian, Bhaskaran, Swaminathan, Desai, Daniel and Datye1995).
Linking predictable resources
Some pathways, particularly in the southern densely settled areas, were frequently used by elephant bulls in the wet season when moving between protected areas, foraging on ripening crops en route. Crop-raiding has been described as an extension of an elephant's optimal foraging strategy, with bulls making use of raised nutritive content (protein, calcium and sodium) of crops at the end of the wet season (Sukumar, Reference Sukumar1990). As in previous studies (Hoare, Reference Hoare1999; Jackson et al., Reference Jackson, Mosojane, Ferreira and Van Aarde2008; King et al., Reference King, Lawrence, Douglas-Hamilton and Vollrath2009) female elephants in this study displayed risk-averse behaviour and avoided settlement areas in peak harvesting time by restricting pathway use to the northern, less densely settled areas. The herds made use of the pathways to access the Kwandu River and protected areas to the west, as the majority of waterholes had dried up. Maize is non-randomly distributed in the landscape, and in the Kwandu Conservancy 75% of fields raided in one year would be targeted again in the following year (von Gerhardt, Reference von Gerhardt2011). Crop fields provide elephants with a clustered and stable, high-nutrient resource at the end of the wet season when nutritional demands are greatest, with bulls coming into musth and females in oestrus (Poole, Reference Poole1984). Crop-raiding, and human–elephant conflict, was found to be a function of season, with incidents only recorded by the Conservancy Office in the wet season, although elephants cross the Conservancy in the dry season to access the Kwandu River and Bwabwata National Park.
In addition, undisturbed riparian habitat with preferred fruiting trees (Sclerocarya, Garcinia, Diospyros, Bauhinia and Acacia spp.) and minerals are available on the western side of the Kwandu River within Bwabwata National Park. Salt licks are provided around pans in Bwabwata National Park (KVG, pers. obs.) and provide an important source of sodium. In the savannahs of the Central African Republic elephants seek out salt-rich soils around termitaria and waterholes and these areas are connected by trails (Ruggiero & Fay, Reference Ruggiero and Fay1994). Elephants, especially females, exercised geophagy in response to sodium deficiencies in Kalahari sand habitats (Holdo et al., Reference Holdo, Holt and Fryxell2009). Duffy et al. (Reference Duffy, Dai, Shannon, Slotow and Page2011) demonstrated that elephants use a direct movement strategy, using decreased path tortuosity and fewer turning angles when needing to get to a destination such as a water source. Waterholes were found in at least half of the pathways in Kwandu Conservancy, and these tended to dry up by the end of April. Elephants resident in the area used the pathways to access the Kwandu River to drink at night.
Facilitating movement in the matrix
Animals are known to have sinuous pathways in good-quality terrain, whereas they tend to move further and faster in unfavourable terrain (Crist et al., Reference Crist, Guerrtin, Wiens and Milne1992). Elephant movement tends to be more directional when resources are limited or habitat heterogeneity is high (Wittemyer et al., Reference Wittemyer, Polansky, Douglas-Hamilton and Getz2008). Habitat connectivity in the Kwandu Conservancy is patchy because of the presence of agricultural fields, with the spatial configuration of the road, settlements and crop fields creating a barrier to wildlife movement (Martin, Reference Martin2006). This environmental gradient of habitat quality should consequently decrease animal movement (Forman, Reference Forman1995). However, in this study temporal segregation of pathway use permitted the elephants to traverse the anthropogenic matrix safely. Elephants and other species were active along pathways at night, with human activity occurring during the day. Pathway use by both elephants and humans has also been noted by Carroll (Reference Carroll1988), with forest elephants and Aka pygmies in the Central African Republic using the vast network of elephant pathways connecting marshy clearings that have high mineral and water content. Other studies confirm nocturnal activity of elephants near smallholdings (Hoare, Reference Hoare1999; Sitati et al., Reference Sitati, Walpole, Smith and Leader-Williams2003; Graham et al., Reference Graham, Douglas-Hamilton, Adams and Lee2009) and research suggests that this is a risk-avoidance strategy (Galanti et al., Reference Galanti, Preatoni, Martinoli, Wauters and Tosi2006; Wittemyer et al., Reference Wittemyer, Polansky, Douglas-Hamilton and Getz2008) used by elephants in unprotected areas.
Fidelity to pathways in disturbed areas could be an effective behavioural strategy when speedy escapes and spatial awareness of shelter areas are required. Spatial awareness during flight behaviour is demonstrated by meerkats Suricata suricatta, for example, which use spatial memory rather than olfactory or visual cues in quickly locating bolt-holes when exposed to threat of predation (Manser & Bell, Reference Manser and Bell2003). In Kwandu Conservancy rural farmers use a combination of methods to discourage elephants from moving though agricultural areas: drumming, burning fires, throwing stones, shouting and firing guns. Ahlering et al. (Reference Ahlering, Millspaugh, Woods, Western and Eggert2011) recorded elevated levels of stress hormones in bulls moving though settlement areas. In Kwandu Conservancy single mature elephant bulls moving along pathways were most commonly observed (von Gerhardt, Reference von Gerhardt2011) but at the height of the harvesting season bull groups of 2–7 individuals of varying ages were recorded, with serious crop-raiding incidents attributed to groups of bulls. Intense burning and runaway bushfires are also a threat to elephants in the peak dry season months. Spatial memory and acuity in an anthropogenic matrix may be advantageous to elephants frequently exposed to disturbance.
Conclusion
Metapopulation theory underpins the management for the Kavango-Zambezi Transfrontier Conservation Area's network of protected areas. Such networks may be a mitigation measure against habitat isolation provided that network design and management is optimized and the spatial arrangement of landscape elements considered. Protected areas have been set aside for this purpose but it is within the surrounding matrix that the strength of the Kavango-Zambezi Transfrontier Conservation Area's effectiveness will be challenged. Dispersal and movement of individual elephants between protected areas is affected by the quality of the surrounding matrix, as well as by the spatial arrangement of habitat fragments beyond protected areas (Cumming, Reference Cumming, Ferguson and Hanks2010). Landscape elements such as elephant pathways, nodes (waterholes/pans) and stepping stones (remnant pockets of forest/forest reserves) in the surrounding matrix need to be identified and to be of sufficiently high quality to encourage elephant movement among patches. This will be especially relevant for herds of risk-averse females and their offspring.
Our study suggests that elephants use pathways when moving between protected areas and that these pathways play a significant role in the location of human–elephant conflict. Crops, fruiting trees, and preferred drinking and feeding areas are spatially fixed, and the elephant pathway network in a given area may provide insights into resource preference at the habitat scale. The spatial arrangement of landscape features and the location of pathways could provide useful tools for designing large conservation landscapes such as the Kavango-Zambezi Transfrontier Conservation Area, notably by indicating where to protect elephant pathways to rivers and where to direct efforts to reduce human–elephant conflict incidents effectively.
Klingelhoeffer (Reference Klingelhoeffer1987) suggested that pathways reflect ancient movement routes. Although it is known that elephant movement is non-random (Wittemyer et al., Reference Wittemyer, Polansky, Douglas-Hamilton and Getz2008; Loarie et al., Reference Loarie, van Aarde and Pimm2009), little is known about the relationship between elephant spatial memory and movement patterns. The extent to which elephant pathways determine elephant movements requires further research, especially in view of the inheritance of geographical knowledge through the matrilineal line.
Acknowledgements
We thank the Office of the President and Government of Namibia for permission to conduct research in the greater Kwandu area, staff from Integrated Rural Development and Nature Conservation (especially Richard Diggle, Beaven Munali and Robin Beatty) and the Kwandu Conservancy Office (especially Cordelia Muyoba and Kenneth Mufaso), and Johnny Clive for invaluable assistance during fieldwork. Stellenbosch University, South Africa, Conservation International, South Africa, and the Shikar Safari Club Foundation, USA, provided financial support. The opinions expressed in this paper are those of the authors and do not necessarily reflect those of Stellenbosch University, Conservation International or the Shikar Safari Club Foundation.
Biographical sketches
Katharina von Gerhardt's research interests lie in the conservation of the African elephant, and the management of conservation areas in Africa. Adriaan van Niekerk is interested in the development and application of geographical information systems and remote sensing techniques to support decisions concerned with land use, biogeographical, environmental and socio-economic problems. Martin Kidd is a statistical consultant at the University of Stellenbosch; he dealt with most of the statistical aspects of this research. Michael Samways researches synthetic management approaches to conservation, mostly at the landscape level. John Hanks is interested in elephant conservation and management linked to transfrontier conservation areas.






