



Introduction
Our knowledge of the early Eocene (Ypresian) Odonata of the insect-rich Okanagan Highlands deposits of southern British Columbia, Canada, and north–central Washington State, United States of America, has expanded rapidly since they were first reported in 2019, most recently by the efforts of local collectors in the villages of Princeton, British Columbia (Allenby Formation) and Republic, Washington (Klondike Mountain Formation). These now include 32 species and 15 genera (one collective) in three suborders (Archibald and Cannings Reference Archibald and Cannings2019, Reference Archibald and Cannings2021a, Reference Archibald and Cannings2022; Archibald et al. Reference Archibald, Cannings, Erickson, Bybee and Mathewes2021). Here, we describe three more species based on recently discovered fossils, and we name two of them.
Material and methods
We examined three fossil wings in Okanagan Highlands lacustrine shale, BBM-P000020A and B and BBM-P000021 from the Allenby Formation near Princeton, British Columbia, Canada, and SR 23-005-003A and B from the Klondike Mountain Formation at Republic, Washington, United States of America.
We follow the terminology for wing venation as used by Garrison et al. (Reference Garrison, von Ellenrieder and Louton2010). By “linear,” we mean that a vein is not zigzagged.
Figures were made with Adobe Photoshop (Adobe, San Jose, California, United States of America) from photographs taken in the laboratory of Parks Canada (Vancouver, British Columbia) with a digital camera mounted on a Zeiss microscope (Zeiss, Oberkochen, Germany). Drawings were made from these with Adobe Illustrator and are taken from both the part and counterpart, except for Allenby gen. and sp. A, for which a counterpart was not recovered.
Institutional abbreviations are: BBM – Beaty Biodiversity Museum, University of British Columbia, Vancouver; PDMA – Princeton and District Museum and Archives, Princeton, both in British Columbia, Canada; and SR – Stonerose Interpretive Center, Republic, Washington, United States of America. We use the Republic locality code B4131 of the Burke Museum of Natural History and Culture (Seattle, Washington), and the Stonerose Interpretive Center.
Character states of compared taxa are placed in square brackets.
Localities
The fossils examined are from the Vermilion Bluffs Member of the Allenby Formation near the village of Princeton, British Columbia (Read Reference Read2000), and exposure B4131 of the Tom Thumb Tuff Member of the Klondike Mountain Formation in the village of Republic, Washington. These are localities of the Okanagan Highlands series of early Eocene (Ypresian) lacustrine shale basins that are scattered across about 1400 km from northwestern British Columbia to north–central Washington. They bear rich fossil records of life in the montane forests surrounding the lakes in which they were preserved. The Okanagan Highland localities mentioned here were temperate uplands with estimated upper microthermal climates during the Early Eocene Climatic Optimum, the warmest sustained interval of the Cenozoic Era (Greenwood et al. Reference Greenwood, Archibald, Mathewes and Moss2005). Although such microthermal climates are today associated with cold winters, in the climatically equable Eocene, Okanagan Highland forests experienced mild winters with few or no frost days (see Archibald et al. Reference Archibald, Greenwood, Smith, Mathewes and Basinger2011 for a summary).
Uranium–lead analyses indicate that both were deposited in the latter half of the Ypresian: the Vermilion Bluffs Member at 51.85 ± 0.85 Ma from samples collected at the “Billy’s Family Restaurant” exposure about 4 km northeast of the fossil site where BBM-P000020 and BBM-P000021 were found, and exposure B4131 of the Tom Thumb Tuff Member of the Klondike Mountain Formation in Republic, Washington, where SR 23-005-003 was found at 51.18 ± 0.09 Ma (both Rubino et al. Reference Rubino, Leier, Cassel, Archibald, Foster-Baril and Barbeau2021).
Systematics
Order Odonata
cf . Suborder Cephalozygoptera
cf . Family Dysagrionidae
cf . Subfamily Dysagrioninae
collective genus Dysagrionites Archibald and Cannings
Figure 1
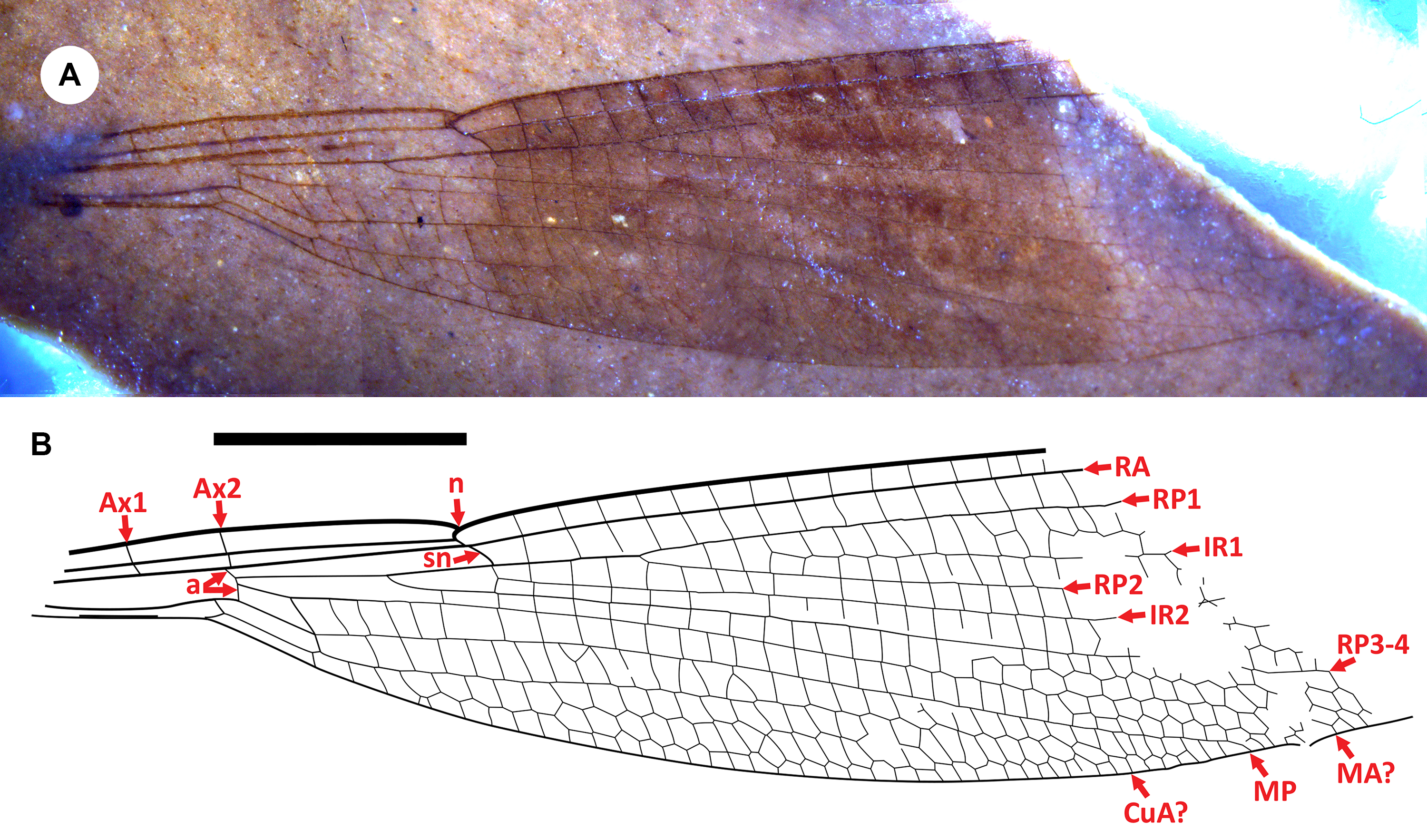
Figure 1. Dysagrionites allenbyensis n. sp. A, holotype BBM-P000020: A, photograph of the part (A-side); B, drawing from the part and counterpart. Both to scale; the scalebar is 5 mm.
Remarks. Archibald and Cannings (Archibald et al. Reference Archibald, Cannings, Erickson, Bybee and Mathewes2021) created the genus-level collective taxon Dysagrionites to function as a holding bin for fossil odonates referrable to the Dysagrioninae (Cephalozygoptera, Dysagrionidae) that can be clearly separated from species of that subfamily but whose nominal generic affinity is unclear by the preservation and completeness of their currently known specimens. Collective taxa are regulated by the International Commission on Zoological Nomenclature (International Trust for Zoological Nomenclature 1999, for example, articles 42.2.1 and 42.3.1).
Archibald et al. (Reference Archibald, Cannings, Erickson, Bybee and Mathewes2021) referred three species to the parataxon: Dysagrionites delinei Archibald and Cannings and Dysagrionites sp. A and Dysagrionites sp. B. They erred, however, in that the parataxon definition required that those species are established as members of the Dysagrioninae, and although this strongly appears to be the case, these fragmentary fossils lack enough of the character states that, in combination, firmly place them in the family and, therefore, in this subfamily. Simonsen et al. (Reference Simonsen, Archibald, Rasmussen, Sylvestersen, Olsen and Ware2024) subsequently established a diagnosis of Dysagrionidae using wings alone; however, known specimens of Dysagrionites species still lack sufficient morphology for confirmation. We emend the collective taxon definition by adding “or tentatively so”: This genus-level collective group name is proposed for any fossil species referable to the Dysagrioninae, or tentatively so, that can be clearly defined at the species level, separating them from other species of the subfamily, but whose orthotaxon generic affinity is unclear by the incomplete preservation of its currently known specimens. Dysagrionites then includes these three species and D. allenbyensis, described here.
If our assumption that D. allenbyensis is a dysagrionid is correct, it belongs to the Dysagrioninae, as the origin of its RP3–4 is about two-thirds the distance between the arculus and nodus and IR2 originates at the subnodus [Petrolestinae: RP3–4 originates more proximally, closer to, or at, the point midway between the arculus and subnodus, and IR2 originates close to, or on, RP3–4; subfamily diagnoses were summarised by Archibald et al. Reference Archibald, Cannings, Erickson, Bybee and Mathewes2021].
Allenbya holmesae Archibald and Cannings is the only previously known non-anisopteran odonate from the Allenby Formation. It is also tentatively treated as a dysagrionid and cephalozygopteran but is distinct enough to be assigned to a nominal genus (Archibald and Cannings Reference Archibald and Cannings2022). It is also a fragmentary wing, but in this case, the distal portion is preserved. It is distinct from D. allenbyensis; for example, IR1 is zigzagged, and the postnodal and postsubnodal crossveins are not aligned.
Dysagrionites allenbyensis sp. nov.
ZooBank Registration number: urn:lsid:zoobank.org:act:E1F8815D-1BB4-4090-99EE-08A9273CEE79
Figure 1
Type material. Holotype BBM-P000020A and B (part and counterpart). A well-preserved wing with colouration, missing the apical portion. Found by Beverley Burlingame on 2 April 2024 and donated to the Beaty Biodiversity Museum, where it is housed.
Diagnosis. The wing of Dysagrionites allenbyensis is most like those of Okanopteryx Archibald and Cannings (three species as discussed below) and Stenodiafanus Archibald and Cannings (one species as discussed below). It is distinct from those of Okanopteryx by: CuA space expanded to three cells wide for much of its length from subquadrangle to terminus [O. jeppesenorum Archibald and Cannings maximum four; O. fraseri Archibald and Cannings usually two, sometimes a single row of three]; pigmented fascia wide, extending from nodus to at least 15 and probably 17 cells beyond it [O. macabeensis Archibald and Cannings nine (holotype) to 11]. It is distinct from Stenodiafanus westersidei Archibald and Cannings by dark fascia [S. westersidei: hyaline throughout] and CuA space width [S. westersidei: maximum two cells in equivalent region preserved].
Description. As above and the following: RP3–4 origin at two-thirds the distance from arculus to subnodus. Postnodal and postsubnodal crossveins almost all aligned. Apex of wing and pterostigma unknown by preservation. No oblique crossvein O. CuA progressively zigzagged in distal quarter; MA zigzagged in distal third (final portion not fully preserved); IR1 almost linear from its origin three cells from origin of RP-2; other main veins linear (but distal portions missing). CuA space expanded in the middle, three cells wide for most of its length. Ax1, Ax2 preserved, arculus aligned with Ax2; subnodus distinctly angled, normal obliquity. Membrane hyaline except for dark fascia mid-wing from nodus to the level of at least 15 postnodal crossveins and probably 17 in total. Geologic shear forces may at times somewhat distort fossils at this exposure, and so the apparent narrowness of the wing might be an artefact (Archibald and Cannings Reference Archibald and Cannings2021b; Archibald et al. Reference Archibald, Mathewes and Aase2023a).
Etymology. The specific epithet is a toponym referring to the Allenby Formation.
Range and age. Vermilion Bluffs Member of the Allenby Formation about 4 km southwest of the village of Princeton, British Columbia. Ypresian (see above).
Remarks. Dysagrionites allenbyensis is immediately notable for its broad, dark fascia mid-wing. Within the Dysagrionidae, such colouration is found only in species of Okanopteryx (Archibald et al. Reference Archibald, Cannings, Erickson, Bybee and Mathewes2021). The genus is known by three species from Okanagan Highlands localities: O. macabeensis (type species) and O. fraseri from the McAbee locality near Cache Creek, British Columbia, and O. jeppesenorum from Republic, Washington (Archibald et al. Reference Archibald, Cannings, Erickson, Bybee and Mathewes2021). Okanagrion hobani Archibald and Cannings (Dysagrionidae) is also known from both Republic and McAbee but has not been found in the Allenby Formation.
The preserved veins of BBM-P000020 agree with those that, in combination, are partly diagnostic of Okanopteryx: no accessory antenodal crossveins; the subnodus is distinctly angled, with normal obliquity; the origin of IR2 is rather evenly spaced between RP1–2 and RP3–4; MA is linear to mid-wing, then becomes increasingly zigzagged; MA and MP are close at margin; CuA is increasingly zigzagged from about mid-wing; the postnodal and postsubnodal crossveins are almost all aligned; and there is no intercalary vein in the CuA–A space. However, other character states necessary to assign a wing to the genus in combination with the above cannot be evaluated on this incomplete fossil: the shape and size of the pterostigma; presence of an oblique brace vein; number of cells wide in the costal space distad the pterostigma; and a distinct convergence of RA, RP1, IR1, RP2 at apex. Dysagrionites allenbyensis does not greatly differ from O. macabeensis in particular and could be a regional variant of that species, close in distance (ca. 150 km) and time (age estimates overlap: McAbee: 52.10 ± 0.26 Ma, Vermillion Bluffs: 51.85 ± 0.85 Ma; both Rubino et al. Reference Rubino, Leier, Cassel, Archibald, Foster-Baril and Barbeau2021).
These preserved character states are also found in Stenodiafanus (see diagnosis in Archibald et al. Reference Archibald, Cannings, Erickson, Bybee and Mathewes2021). The broad, dark fascia of BBM-P000020 might easily be present convergently in a species of that genus, and the width of the CuA space could vary among its species as it does among those of Okanopteryx. Stenodiafanus westersidei is known only from the Klondike Mountain Formation, about 150 km distant and close in time (estimated ages overlap: Klondike Mountain B4131 at 51.18 ± 0.09 Ma and Vermillion Bluffs as above; both Rubino et al. Reference Rubino, Leier, Cassel, Archibald, Foster-Baril and Barbeau2021). McAbee, the Allenby Formation, and the Klondike Mountain Formation had similar climates and forests (Greenwood et al. Reference Greenwood, Archibald, Mathewes and Moss2005; Archibald et al. Reference Archibald, Greenwood, Smith, Mathewes and Basinger2011).
Subfamily incertae sedis
Genus Paradysagrion gen. nov.
ZooBank Registration number: rn:lsid:zoobank.org:act:9540B488-96D6-4697-BCCE-82C7BAE16E10
Figure 2
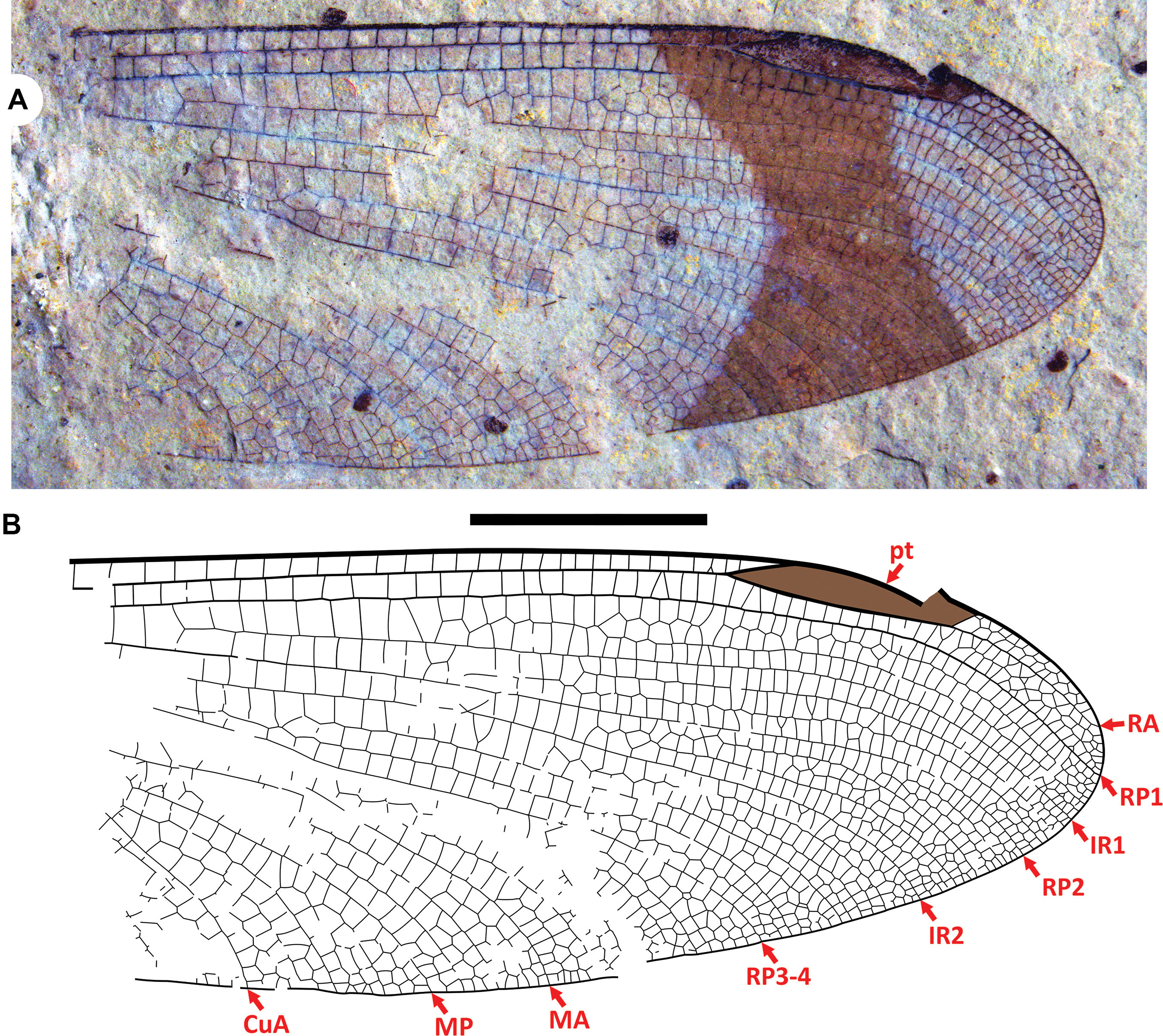
Figure 2. Paradysagrion sosbyae gen. and sp. nov. holotype SR 23-005-003: A, photograph of the part (A-side); B, drawing from the part and counterpart. Both to scale; the scalebar is 5 mm.
Diagnosis. Preserved wing portion most similar to that of Dysagrion Scudder by: pterostigma about 6.3 times longer than wide [Dysagrion species: 4.0–7.5]; many intercalated linear (or nearly so) supplementary sectors between main veins from RP1 to probably CuA (see below; cf. figures of Archibald et al. Reference Archibald, Cannings, Erickson, Bybee and Mathewes2021, cited below); IR1 linear from origin as in Dysagrion fredericii Scudder (see Archibald et al. Reference Archibald, Cannings, Erickson, Bybee and Mathewes2021, fig. 6), slightly zigzagged in Dysagrion pruettae Archibald and Cannings (see Archibald et al. Reference Archibald, Cannings, Erickson, Bybee and Mathewes2021, figs. 17–18) and Dysagrion lakesii Scudder (see Archibald et al. Reference Archibald, Cannings, Erickson, Bybee and Mathewes2021, fig. 5), fully zigzagged in Dysagrion packardii Scudder (see Archibald et al. Reference Archibald, Cannings, Erickson, Bybee and Mathewes2021, fig. 4).
It differs from wings of Dysagrion by: (1) RA, RP1, IR1 subparallel to apex, apex between terminus of RA, RP1 [Dysagrion: converging at apex, RA at or near apex]; (2) CuA (presumed, see below) linear to termination [Dysagrion: heavily zigzagged]; (3) no brace vein [present in D. pruettae; region poorly or not preserved in other species]; (4) RA–RP1 space between pterostigma and apex three cells wide in most of distal half, seven at margin [Dysagrion: one]; (5) C–RA space distal to pterostigma two cells wide [Dysagrion: one]; (6) RP1, IR1, and RP2 spaces, each with two linear supplementary sectors, terminate with similar widths [Dysagrion: without supplementary sectors, and RP1, IR1 terminate very close, IR1, RP2 not]; and (7) subnodus distinctly angled with normal obliquity [Dysagrion: reverse obliquity].
Description. As for its only species, below.
Etymology. The genus name is formed from the Greek -para, “alongside,” and Dysagrion, referring to similarities of its wings to those of that genus. Gender, neuter.
Remarks. Dysagrion (cf. Archibald et al. Reference Archibald, Cannings, Erickson, Bybee and Mathewes2021, figs. 4–6, 16–18) is also known from exposure B4131 of the Klondike Mountain Formation. Its most similar species, D. fredericii, is from the Green River Formation of Wyoming, also in the latter portion of the Ypresian.
Paradysagrion sosbyae sp. nov.
ZooBank Registration number: urn:lsid:zoobank.org:act:AC2D2DFA-47B2–46C6–8EE3-B564791E4C72
Figure 2
Type material. Holotype SR 23-005-003A and B (part and counterpart) by monotypy. The well-preserved distal half of a wing found by Emmalin Sosby, 28 May 2023, at exposure B4131 of the Tom Thumb Tuff Member of the Klondike Mountain Formation in the village of Republic, Washington. Housed in the Stonerose Interpretive Center collections, Republic, Washington.
Diagnosis. See genus diagnosis.
Description. Holotype wing. As in genus diagnosis and: dark, hourglass-shaped fascia in pterostigma region; postnodal, postsubnodal crossveins not aligned. C–RA space apical to pterostigma close to half the width of adjacent RA–RP1 space.
Etymology. The specific epithet is derived from the surname of Emmalin Sosby, who found and donated the fossil, in recognition of her contribution. Gender, feminine.
Allenby Odonata gen. and sp. A
Figure 3
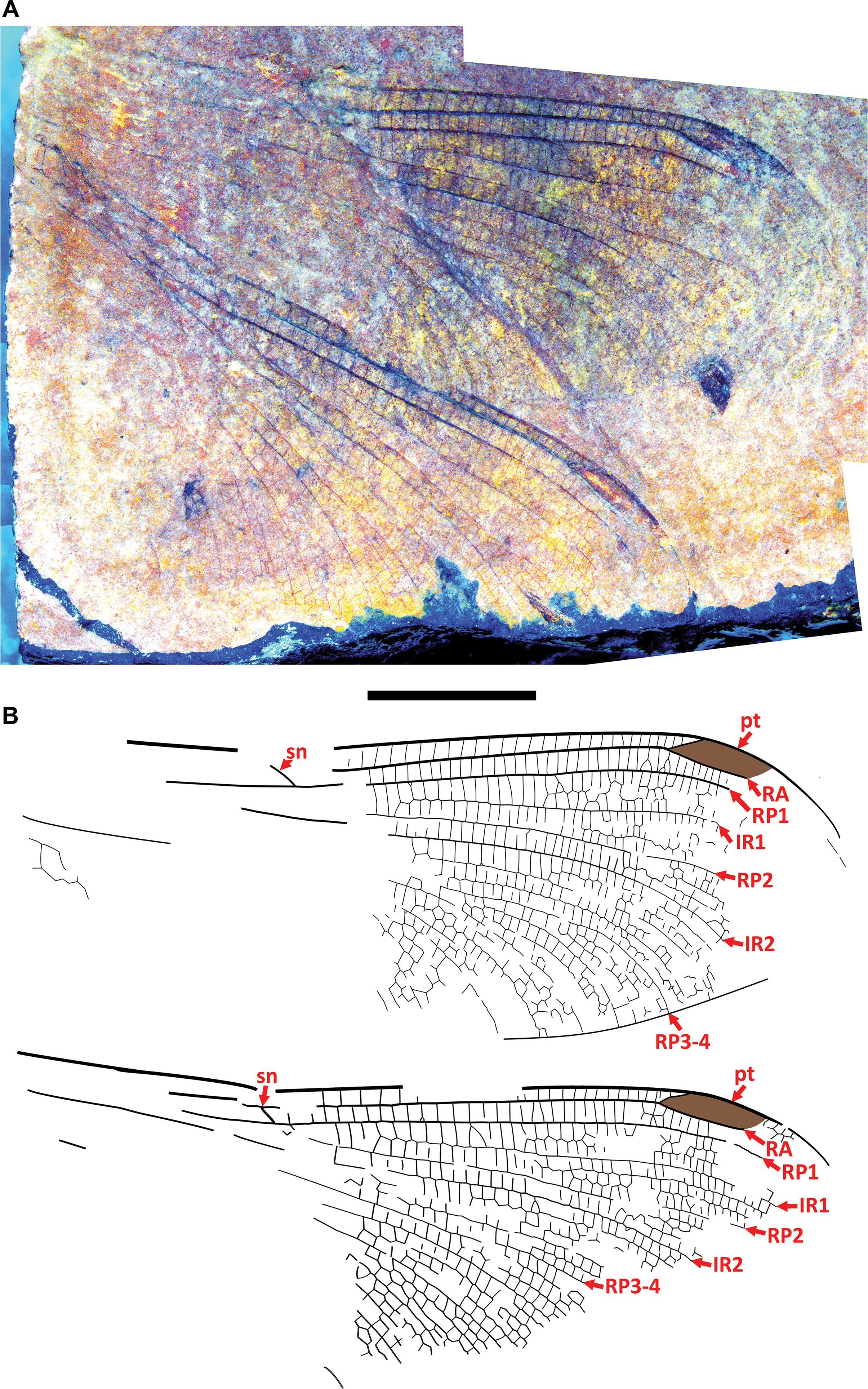
Figure 3. Allenby Odonata gen. and sp. A, BBM-P000021: A, photograph; B, drawing. Both to scale; the scalebar is 5 mm.
Material. BBM-P000021, original PDMA collection number PMF.2023.001.0031. No counterpart. Distal portions of two wings with poorly preserved fragments of apical-most regions. Found by Beverley Burlingame on 4 August 2023 and donated to the Princeton and District Museum and Archives, then to the Beaty Biodiversity Museum, where it is now housed.
Description. Distal portion of the wing, as in remarks (below), and the following. Preserved portion hyaline. Pterostigma about four times longer than wide; no brace vein; most cells in much of anterior region about twice (or more) as high as wide; many linear (or nearly so) supplementary sectors between main veins obscuring identities of partially preserved main veins posterior to RP3–4.
Range and age. Vermilion Bluffs Member of the Allenby Formation about 4 km southwest of the village of Princeton, British Columbia. Ypresian (see above).
Remarks. The wings of BBM-P000021 are more generalised and difficult to assign to a genus and species than are those of other Allenby Formation non-anisopteran odonates. Some of its morphology, however, is informative: its pterostigma is the general size and shape found in many Okanagan Highlands Dysagrionidae; its postnodal and postsubnodal crossveins are not aligned; its C–RA space distad the pterostigma is more than one cell wide and most likely not more than two; it lacks a brace vein; it has a distinctly angled subnodus of normal obliquity; it has numerous linear supplementary sectors; IR2, RP3–4, and likely at least some other incompletely preserved main veins are rather strongly curved distally; and there are narrow cells in the posterior region of the wings, many twice as high as wide.
Although all preserved distal portions of the BBM-P000021 wings agree with those of the Dysagrionidae, the character states that might diagnose the species as a member of that family are found in the basal half of their wings, and some Zygoptera also have similar venation, particularly in the distal part of their wings (Archibald et al. Reference Archibald, Ware, Rasmussen, Sylvestersen, Olsen and Simonsen2023b; Simonsen et al. Reference Simonsen, Archibald, Rasmussen, Sylvestersen, Olsen and Ware2024). Therefore, we tentatively associate BBM-P000021 with similar odonates that are also known from the Ypresian, especially regionally, pending more complete future specimens.
By the above, BBM-P000021 is similar to the wings of Okanagrion threadgillae Archibald and Cannings (Dysagrionidae) (see Archibald et al. Reference Archibald, Cannings, Erickson, Bybee and Mathewes2021, fig. 20) from Klondike Mountain locality A0307, about 670 m from B4131; Primorilestes madseni Rust et al. (Dysagrionidae), from the early Ypresian Fur Formation of Denmark (Rust et al. Reference Rust, Petrulevičius and Nel2008, fig. 2); and Tynskysagrion brookeae Bechly et al. (Green River Formation, Wyoming, United States of America, placed by Bechly et al. Reference Bechly, Garrouste, Aase, Karr, Grande and Nel2020 in Eodysagrioninae, which they treated as a subfamily of Dysagrionidae but was later excluded from that subfamily by Simonsen et al. Reference Simonsen, Archibald, Rasmussen, Sylvestersen, Olsen and Ware2024). We, therefore, treat BBM-P000021 as Allenby gen. and sp. A, also tentatively in the Dysagrionidae and Cephalozygoptera.
Acknowledgements
We thank Emmalin Sosby, who found SR 23-005-003 and donated it to the Stonerose Interpretive Center, and Travis Wellman (Stonerose managing director) for loaning it to us; Beverley Burlingame, who found and donated BBM-P000020 to the Beaty Biodiversity Museum and BBM-P000021 to the Princeton and District Museum and Archives, Kathy Simpkins (PDMA curator of fossils) and Todd Davidson (PDMA director) for its donation to the Beaty Biodiversity Museum and for their long-term generous help; and Marlow Pellatt (Parks Canada, Vancouver) for use of the digital camera/microscope in his laboratory. S.B.A. thanks Rolf Mathewes (Simon Fraser University, Burnaby, British Columbia) for support.




 Open access
Open access