


Introduction
In a world with biodiversity under increasing threat from anthropogenic activities, it can be difficult to prioritise the limited resources available to conservationists. Big, charismatic species may receive a disproportionate share of resources at the expense of small, nondescript or inaccessible species (Leader-Williams and Dublin Reference Leader-Williams and Dublin2000). Systematic, objective ways of determining conservation status are thus increasingly important in a world in which conservation is easily driven by emotional rather than logically defensible criteria.
The understanding of bird distributions and how they are changing is crucial for effective bird conservation (Gaston Reference Gaston2003). In order to determine the conservation status of any species, information is needed on range size, population size and population trend (IUCN Standards and Petitions Subcommittee 2014). Furthermore, information on habitat integrity and population fluctuations is also considered where available. However, these data are lacking for most species globally.
We explored ways of facilitating the conservation assessment process in regions of the world with atlas data. Here we focus on southern Africa, defined for this purpose as South Africa, Lesotho and Swaziland. In South Africa alone, there are over 600 species of breeding birds in a country of 1.2 million km2, making biodiversity monitoring a challenging task (Taylor et al. Reference Taylor, Peacock and Wanless2015). However, the region is increasingly known internationally for its high-quality long-term, large-scale public participation projects. Such citizen science projects make it possible for observations made by many different people to be pooled and analysed as a whole (Cohn Reference Cohn2008). The first and second Southern African Bird Atlas Projects (SABAP1, 1987–1992, and SABAP2, 2007–present) are among Africa’s biggest biodiversity databases, and provide overviews of avian distribution across southern Africa approximately 20 years apart (Harrison et al. Reference Harrison, Underhill and Barnard2008). Spatial records in this database show changes in species’ distributions (range), and by comparing reporting rates (the proportion of checklists reporting a species - a proxy for relative abundance) between these projects we can estimate population change. This information has been used to examine issues of conservation interest, including the influence on birds of climate change (Walther and Niekerk Reference Walther and Niekerk2014); identification of non-climatic drivers of range change (Péron and Altwegg Reference Péron and Altwegg2015a) and changes in timing of migration (Bussière et al. Reference Bussière, Underhill and Altwegg2015), as well as other questions of ecological interest (e.g. Péron and Altwegg Reference Péron and Altwegg2015b, Péron and Altwegg Reference Péron and Altwegg2015c, Cooper et al. Reference Cooper, Wannenburgh and Cherryin press). A few publications (e.g. Kemp et al. Reference Kemp, Herholdt, Whyte and Harrison2001, Lee and Barnard Reference Lee and Barnard2015) have used this information to inform conservation decisions for some species, but despite evidence that reporting rate declines are related to observed population decline (Amar et al. Reference Amar, Cloete and Whittington2015), there has been little testing of the robustness of these measures and they have not been used formally at a national scale to inform conservation decisions.
We explore the capacity of this atlas project to support conservation status assessment by overviewing the information that can be obtained from the SABAP projects for 58 southern African endemic and near-endemic bird species. We present population change indices together with a measure of statistical confidence in these, as well as range-size, range-size change and range connectivity metrics. This information should be important for those evaluating the conservation status of these species in southern Africa.
Methods
SABAP data
The SABAP data sets consist of bird lists compiled by birding citizen scientists. SABAP1 used quarter degree grid cells (15’x15’) as the sampling unit, corresponding to standard southern African 1:50,000 topographical maps (Harebottle et al. Reference Harebottle, Smith, Underhill and Brooks2007). The SABAP2 spatial sampling unit is the 5’x5’ pentad. There are nine pentads nested within each grid cell, so we aggregated the data of the second phase, SABAP2, at the quarter degree resolution in order to compare both phases. Birders were asked to submit lists of all species that they saw or heard during visits to grid cells of between two hours and five days. As of 2014, SABAP2 data existed for over 3,000 grid cells, with country-wide coverage illustrated used in our analysis displayed in Figure 1.

Figure 1. Area and intensity of coverage during the two atlas periods. The colour represents the number of lists reported for each grid cell, with dark grey (red online) high and light grey (blue online) low. SABAP2 coverage is shown to 30 May 2014, as the project is ongoing.
We consider those species with > 70% of range or population within South Africa, Lesotho and Swaziland as near-endemics, as listed by Birdlife South Africa (BLSA; Lotz et al. Reference Lotz, Allan, Bowie, Chittenden, Cohen, Dowsett, Gibbon, Hardaker, Marais, Peacock, Retief, Ryan, Smit-Robinson and Taylor2014). We used this list to obtain the national and global conservation status (‘Least Concern’ [LC] to ‘Endangered’ [EN]) for these species. We restrict our analysis to this subset of the > 840 species in the SABAP database, as population trends identified for endemics and near-endemics can be more accurately inferred from this analysis than for species with significant ranges outside the survey area.
Of the 69 endemics on the above list, we consider 58 after excluding those with recent taxonomic splits. Several species in the BLSA checklist have been split since SABAP1 and are represented as two or more species in SABAP2. We do not consider new southern African species split from species with a combined range that extends beyond the study area. This includes: Hottentot Buttonquail Turnix hottentottus, Karoo Thrush Turdus smithi, Cape Parrot Poicephalus robustus, and the Long-billed Lark Certhilauda spp. complex. The data used in this analysis were accessed from the SABAP2 database over 29–30 May 2014.
Reporting rate
The reporting rate is the number of times a species was reported in a grid cell divided by the total number of checklists for that grid cell. There is evidence that reporting rates are monotonically related to abundance (Amar et al. Reference Amar, Cloete and Whittington2015, Griffioen Reference Griffioen2001, Robertson et al. Reference Robertson, Simmons, Jarvis and Brown1995). Reporting rate data are publicly available for each species from http://sabap2.adu.org.za/ and are a useful first step in quantifying changes in abundance (e.g. Huntley et al. Reference Huntley, Altwegg, Barnard, Collingham and Hole2012). For between-atlas period comparisons we select only the subset of data that were sampled on at least two occasions during each atlas period (n = 2,005 grid cells). We calculate a summary reporting rate change metric based on the average reporting rate across all grid cells for each project for which a species was ever present:
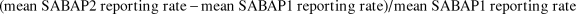 $$\left( {{\rm{mean}}\,SABAP2\,reporting\,rate - {\rm{mean}}\,SABAP1\,reporting\,rate} \right)/{\rm{mean}}\,SABAP1\,reporting\,rate$$
$$\left( {{\rm{mean}}\,SABAP2\,reporting\,rate - {\rm{mean}}\,SABAP1\,reporting\,rate} \right)/{\rm{mean}}\,SABAP1\,reporting\,rate$$
where positive values indicate increase, and negative values indicate decrease. We express this ratio as a percentage. The premise behind this metric is based on the concept of the regression to the mean: while extreme results on a site by site basis certainly exist, the mean across the population should tend towards a stable range of values.
As reporting rate change within a grid cell is undefined if the species was not recorded in that cell during SABAP1, due to division by zero, we create a standardised index of population change that allows us to present variation in change across a species range. For each grid cell we calculated the population change metric as follows:
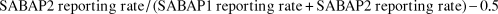 $$SABAP2\,reporting\,rate\,/\,\left( {SABAP1\,reporting\,rate + SABAP2\,reporting\,rate} \right) - 0.5$$
$$SABAP2\,reporting\,rate\,/\,\left( {SABAP1\,reporting\,rate + SABAP2\,reporting\,rate} \right) - 0.5$$
This metric returns a value between -0.5 and 0.5, with values > 0 indicating increases, and values < 0 indicating declines (adapted from Amar et al. Reference Amar, Redpath, Sim and Buchanan2010). A population change map for each species is available as electronic supplementary information, together with reporting rate and range change maps. For an overview of population trends across southern Africa for this set of species, we calculate the mean of population change across all species from within each grid cell as a population change map. We correlate the population change metric against each of the further metrics described below using Pearson’s product-moment correlations in R (R Core Team 2015).
As a final visual representation of change, based on list data for the set of endemic birds, we calculate the ratio of lists with a species recorded to lists without that species for each atlas period. This is the presence/absence ratio (presented in Cunningham et al. Reference Cunningham, Madden, Barnard and Amar2016). The log of the mean of these metrics across all grid cells plotted against each other allows a visualization of species that are doing well in SABAP2 compared to SABAP1 as a function of range. We emphasise that species close together on the resulting chart do not necessarily have similar populations, as the reporting rates are influenced by species detectability; for instance, large or vocal species are likely reported more frequently than expected, given density.
The standard statistic for the equality of two proportions (z-score; Underhill and Bradfield Reference Underhill and Bradfield1998) can be used as an index to measure confidence in change in relative abundance that accounts for the number of lists submitted for each grid cell for each period. The following is the formula as described in Underhill and Brooks (Reference Underhill and Brooks2014):
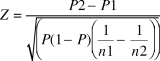 $$Z = {{P2 - P1} \over {\sqrt {\left( {P\left( {1 - P} \right)\left( {{1 \over {n1}} - {1 \over {n2}}} \right)} \right)} }}$$
$$Z = {{P2 - P1} \over {\sqrt {\left( {P\left( {1 - P} \right)\left( {{1 \over {n1}} - {1 \over {n2}}} \right)} \right)} }}$$
where P1 and P2 are the reporting rates from SABAP1 and SABAP2 respectively, n1 and n2 are the numbers of checklists on which the reporting rates are based, and P, reporting rate, is given by:
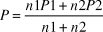 $$P = {{n1P1 + n2P2} \over {n1 + n2}}$$
$$P = {{n1P1 + n2P2} \over {n1 + n2}}$$
We calculate the mean of the z-score for the grid cells in a species range as an index of confidence in the direction of population change for each species: large negative values indicate evidence for population decline, large positive values indicate evidence for population increase. Values close to zero indicating unclear status: populations could be declining, increasing or not changing.
Population change metric validation with dynamic occupancy modelling
Treating reporting rate as a proxy for abundance relies on the premise that variation in detection probability is largely due to variation in abundance. This assumption is shared with other abundance estimators that are based on detection / non-detection data (Péron and Altwegg Reference Péron and Altwegg2015a, Royle and Nichols Reference Royle and Nichols2003) and appears to be reasonable for the SABAP data (Huntley et al. Reference Huntley, Altwegg, Barnard, Collingham and Hole2012, Robertson et al. Reference Robertson, Simmons, Jarvis and Brown1995). We also assume that the trends in species abundance in areas not well covered (notably the arid central western regions) were similar to those in well-covered areas. As we cannot validate these assumptions and it has been shown that simple metrics can produce biased trend estimates when sampling is not considered (Isaac et al. Reference Isaac, Strien, August, Zeeuw and Roy2014), we test the population change metric described above against ‘probability of reporting’ change between atlas periods based on 191 common species from dynamic occupancy modelling methods proposed by Bled et al. (Reference Bled, Nichols and Altwegg2013) and presented in Péron and Altwegg (Reference Péron and Altwegg2015a). These models attempt to account for variation in detectability that is a consequence of observer, habitat and season. However, due to the large number of variables these models are unstable for species with low reporting rates and small ranges i.e. most of the species in our set of endemic birds.
We present correlation coefficients at the community level using mean values for each species based on the summary metrics explained above and probability of reporting change for the set of 191 bird species. We also examine correlation between population change scores and z-scores with probability of reporting change for each of the 20 endemic species at the QDGC level within the set of 191 birds. Lastly, we examine the relationship between the correlation coefficient output for the last analysis with the 20 endemic species with the log-normalised number of QDGCs to examine the influence of range size on these comparisons.
Range and range change
Between atlas periods, ranges of some species expanded while others have contracted. To capture a snapshot of net gain or loss in range, we use the following calculation based on grid cells in which a species has been recorded:
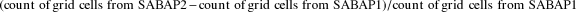 $$\left( {count\,of\,grid\,cells\,from\,SABAP2 - count\,of\,grid\,cells\,from\,SABAP1} \right)\,/count\,of\,grid\,cells\,from\,SABAP1$$
$$\left( {count\,of\,grid\,cells\,from\,SABAP2 - count\,of\,grid\,cells\,from\,SABAP1} \right)\,/count\,of\,grid\,cells\,from\,SABAP1$$
Plotting reporting rate change against range change is useful for gauging how well a species is doing compared to other species.
To exclude that range where perhaps a species was vagrant or possibly incorrectly recorded in SABAP1, we excluded grid cells that had > 50 lists but only 1 record in SABAP1 and call this core range. We calculated core range change as above for range change; but this is a stricter measure of range change.
We calculated the total number of grid cells where a species was recorded over both atlas periods. This value multiplied by the approximate area covered by a grid cell, 729 km2, we call the species SABAP range, which we consider a surrogate for Extent of Occurrence (EOO; the minimum convex polygon encompassing all known normal occurrences of a particular species). We compare these to ranges from BirdLife International species accounts from the BirdLife Data Zone (http://www.birdlife.org/datazone/home) using standard correlation tests in R (R Core Team 2015). We also calculate the area of pentads from which a species has been recorded and treat this finer scale reporting as an indication of Area of Occupancy (AOO; the subset of the EOO where the species actually occurs).
Connectivity index
For each species we calculated a range connectivity score. Each grid cell where a species was recorded was scored for the presence of the species in the four neighbouring grid cells to the north, south, west and east, being those grid cells with greatest surface area contact. The maximum score is four for a grid cell surrounded by other occupied grid cells, while an isolated grid cell will have a score of 0. For each species we record the mean connectivity score across the species range. This index may be influenced by detection probability: a species with a checker-board pattern might occur widely but be hard to detect (e.g. Peregrine Falcon Falco peregrinus). As this score is a function of the area of a species range, we correct by dividing the connectivity score by the log of the number of grid cells in which a species occurs.
Results
Population change for southern African endemics in relation to reporting rate and range change
Displaying population metrics for this set of species allows one to examine individual species trends in the broader context of species trends for the region. In the example of what we call southern African endemics, population trends were mixed, with mean reporting rate lower for 33 species, and higher for the remaining 25 (mean population change = -0.06 ± 0.09). There was a correlation between range change and reporting rate change (t = 6.7, P < 0.01, df = 56, Figure 2); as, generally, if a species is no longer reported from a grid cell this is reflected in both metrics. A species with reporting rate change < -30% and range change < -30% (bottom left, Figure 2, Figure 3) may be a species of conservation concern based on IUCN criteria A (population size reduction), where population reduction (measured over the longer of 10 years or three generations) is greater than 30%. Several species with positive range and reporting rate change metrics are still species of conservation concern, as there may be reasons other than population change metrics for considering their status (e.g. population size and fragmentation).

Figure 2. Reporting rate change for 58 South African endemic bird species plotted against change in reported range between SABAP1 and SABAP2. Point size represents the absolute value of the mean z-score.

Figure 3. Reporting rate change for seven South African endemic bird species plotted against change in range between SABAP1 and SABAP2. This figure shows the lower left hand corner of Figure 2 in more detail - species qualifying as those of conservation concern due to range and population decrease. Size of the points is weighted by mean z-score.
There are four species currently listed as ‘Least Concern’ that merit further investigation into their conservation status: Ground Woodpecker Geocolaptes olivaceus, Drakensberg Rockjumper Chaetops aurantius, Sentinel Rock Thrush Monticola explorator and Gurney’s Sugarbird Promerops gurneyi (Figure 3). All these species are associated with upland areas or the grassland biome, as are the three species with existing threatened status in Figure 3. Confidence in population change (mean z-score) of species of conservation concern was lowest for Botha’s Lark Spizocorys fringillaris, but the total number of grid cells where the species was ever recorded was only 20. The associated range change for Botha’s Lark between atlas periods was -64%. The total core range of this species was 15 grid cells, with a range change of -50%: an alarming apparent contraction.
The seven species identified in Figure 3 as species of conservation concern are all species for which the confidence measure (mean z-score) across grid cells was within the lower quartile of values for the total set of species (Table 1). In addition to the above species, those species with large measures of confidence in decrease (negative z-scores) were: Orange-breasted Sunbird Anthobaphes violacea, Cape Rockjumper Chaetops frenatus, Protea Seedeater Crithagra leucoptera, Pied Starling Lamprotornis bicolor and Grey-winged Francolin Scleroptila africana. For these species we are more confident there are population declines possibly as they are associated with areas with large atlas efforts, although the magnitude of these declines may not necessarily meet IUCN criteria for threatened status without concurrent declines in reporting rate. Presence/absence ratios for Protea Seedeater, Grey-winged Francolin and Pied Starling are low (Figure 4) and appear among the set of species faring most poorly according to this measure.

Figure 4. Presence/absence ratios for 58 endemic bird species for each atlas period. Species on the negative end of the x-axis are generally infrequently reported, while those on the positive side are commonly reported: negative values indicate species reported from less than 50% of cells. Shading represents the 95% confidence interval of the regression between the values on the two axes. Species below the 1:1 line (black diagonal) are species reported less frequently in SABAP2. Selected species classified as Least Concern with a lower reporting rate in SABAP2 are labelled, as are selected species with threatened status with higher reporting rates in SABAP2.
The set of species for which we are most confident of population declines are those associated with grassland and fynbos (a biome restricted mostly to the Western and Eastern Cape provinces (Cowling Reference Cowling1995); Table 1, Figure 5). The overview map of population change suggests endemic species as a whole are faring particularly poorly around Swaziland and north-eastern South Africa. The fynbos biome (a fire-driven ecosystem dominated by shrubs, geophytes and the Restionaceae family; Cowling Reference Cowling1995) in South Africa’s most south-western corner is also an area with largely negative trends.

Figure 5. Mean population change across all species within each grid cell (left panel). N for each grid cell is indicated by endemic species richness (right panel). Grids not included in this analysis due to insufficient coverage (<2 lists for both atlas periods) are white with black points.
Population change metric validation with dynamic occupancy modelling
For a set of 191 passerine species for which probability of reporting change was calculated from dynamic occupancy modelling (Péron and Altwegg Reference Péron and Altwegg2015a), at the community level there was a significant positive correlation with both our population change metric (rs = 0.46, t = 7.1, df = 189, P < 0.001) and mean z-scores (rs = 0.56, t = 9.3, df = 189, P < 0.001). However, in the analysis at the species level for the 20 endemic species for which we had occupancy estimates at the grid level, 13 species showed a significant positive correlation between the dynamic occupancy modelling probability of reporting change and population change; while 15 species showed significant correlation with z-scores. Lastly, the size of range seemed to influence this relationship as there was a significant negative correlation between correlation coefficient output from the above analyses and range size for the 20 endemics (rs = -0.79, t = -5.3, df = 18, P < 0.001) suggesting this relationship between occupancy model metrics and our metrics is weak for species with smaller ranges.
Range
The SABAP ranges of endemic and near-endemic species in southern Africa were generally large (> 20,000 km2; an IUCN threshold criteria for determining threatened species status). SABAP range and published EOO values were highly correlated, with those from SABAP lower on average (BLI EOO 348,306 km2; SABAP range 291,939 km2; t = 16.7, P < 0.01, df = 56). Only two species had total ranges < 20,000 km2: Botha’s Lark and Rudd’s Lark Heteromirafra ruddi. Overall, occupied area as calculated from pentad data was on average 23.6% of that of SABAP range. Apart from Rudd’s and Botha’s Larks, only Mountain Pipit Anthus hoeschi had the pentad area close to 2,000 km2, representing that AOO threshold under which a species might meet conservation status criteria. There was no correlation between population change and SABAP range (t = 0.1, P = 0.91, df = 56); or the pentad area from which a species was recorded (t = 1.4, P= 0.16, df = 56).
Range connectivity
Metrics of connectivity varied widely among the set of endemic bird species. Species identified as those of conservation concern by Lotz et al. (Reference Lotz, Allan, Bowie, Chittenden, Cohen, Dowsett, Gibbon, Hardaker, Marais, Peacock, Retief, Ryan, Smit-Robinson and Taylor2014) dominated the cluster of species with high fragmentation and small ranges, both corrected and uncorrected (Table 1, Figure 6). There was a significant positive correlation between the corrected connectivity score and population change (t = 2.6, P = 0.01, df = 56) with species with negative population change also those species with low connectivity. Pied Starling, a widely distributed arid-zone generalist, had the highest connectivity overall. On the other hand, the arid-zone Cinnamon-breasted Warbler Euryptila subcinnamomea had the lowest connectivity score.

Figure 6. Connectivity (left-hand panels) and corrected connectivity (connected score/log(range); right hand panels) for southern African endemic bird species. The lower two charts are the lower left sections of the upper charts, indicating species with small ranges and low connectivity; QDGC = quarter degree grid cells.
Discussion
Citizen science projects like bird atlas projects have an important role to play in biodiversity conservation (Robertson et al. Reference Robertson, Cumming and Erasmus2010). In this article we have shown ways in which species atlas data can be used to develop population parameters that can assist conservation assessment of bird species. However, we regard our analysis as only one approach to be used alongside other lines of evidence when assessing the conservation status of species. In our case study from southern Africa, SABAP2 is a dynamic dataset which facilitates the exploration of numerous ecological and conservation questions. It has been effective as ‘an early warning system’ to alert conservationists of species in trouble (Barnard and Villiers Reference Barnard and Villiers2012, Lee and Barnard Reference Lee and Barnard2012).
Population change
Both Loftie-Eaton (Reference Loftie-Eaton2014) and Péron and Altwegg (Reference Péron and Altwegg2015a) found evidence that the species sets that they examined using atlas data were reported more frequently during SABAP2. The latter study of 191 widely distributed species reported only increases in probability of recording between atlas periods. In contrast to these two studies, several endemic species from our study show evidence for population declines.
In southern Africa, the five worst-faring birds by standardised population change have high affinities to the grassland biome or mountain regions. Of these, Botha’s Lark has been identified as ‘Endangered’. However, we identify four species listed as ‘Least Concern’ that show changes in population status and range size which qualify them as species of conservation concern: Ground Woodpecker, Sentinel Rock Thrush, Drakensberg Rockjumper and Gurney’s Sugarbird. It has been noted that Gurney’s Sugarbird is adversely impacted by inappropriate fire regimes (de Swardt Reference de Swardt2010), but there is little published on the other species. It has previously been shown using SABAP data that the species diversity of grassland birds generally, and globally threatened grassland birds in particular, is significantly and negatively correlated with the extent of afforestation (Allan et al. Reference Allan, Harrison, Navarro, van Wilgen and Thompson1997). Furthermore, climate envelope modelling suggests that fynbos and grassland bird species are among those most at risk from global climate change (Huntley and Barnard Reference Huntley and Barnard2012). Our analysis suggests that some species of this biome are in detectable decline over the relatively short period of time between atlas periods.
Several species with grassland affinities show signs of positive population change: Cloud Cisticola Cisticola textrix, Melodious Lark Mirafra cheniana, Mountain Pipit and Southern Bald Ibis Geronticus calvus. However, long term monitoring suggests that Southern Bald Ibis continues to show moderate declines at breeding sites, with a breeding population of < 2,000 pairs (C. Henderson, unpubl. data). This species is a colonial breeder that often forages in groups. Bird atlas data may not be a sufficiently sensitive early warning tool for population declines of charismatic or predictably flocking species, as their abundance is not directly recorded and so even large declines in mean flock size would not be reflected in some cases. However, this does suggest that Pied Starling and Grey-winged Francolin, species fitting this description where declines have been observed, may be worthy of special attention. Mountain Pipit shows an unusual situation that reporting rate change was very positive between SABAP1 and SABAP2, which may be a consequence of small range size, for which these summary metrics become unstable. The range for Mountain Pipit showed moderate decrease, and total current range may be under 20,000 km2 within South Africa. Melodious Lark, currently with IUCN red list status ‘Near Threatened’ attributed to moderately rapid population decline, is likely stable. Our analysis supports the most recent local regional ranking of ‘Least Concern’ (Taylor et al. Reference Taylor, Peacock and Wanless2015).
Species with high affinity for forest generally showed little sign of population decreases. Knysna Warbler Bradypterus sylvaticus appears to have expanded its range eastwards despite lower coverage in this part of the species range during SABAP2. This species still exhibits a low degree of range connectivity due to its reliance on isolated forest patches. Knysna Woodpecker Campethera notata was the species that fared the best of all the endemics by various criteria. It is classified as ‘Near Threatened’ due to historical loss of range from the east coast, coupled with small estimated populations within protected areas. However, population is currently stable, and in the absence of further threats this species might be classified as ‘Least Concern’.
Two arid zone species showed core range change declines of > 35%: Sclater’s Lark Spizocorys sclateri and Black-eared Sparrow-lark Eremopterix australis. Cinnamon-breasted Warbler showed reporting rate declines of >30% and very low range connectivity. There are concerns for bird populations of southern Africa’s arid zones, as this area is experiencing dramatic increases in extreme heat events (Cunningham et al. Reference Cunningham, Kruger, Nxumalo and Hockey2013). However, the area has been poorly covered during SABAP2 and by contrast Red Lark Calendulauda burra showed increases in reporting rate and moderate range increases. There is thus a continued need for improved atlas coverage of dry regions before concrete conclusions can be made regarding arid-zone species using atlas data.
Birds with affinity for fynbos generally also fared poorly, with Protea Seedeater Crithagra leucoptera and Cape Rockjumper showing declining trends. Cape Rockjumper has been identified as vulnerable to warming due to climate change (Lee and Barnard Reference Lee and Barnard2015), while Protea Seedeater declines can be attributed to decrease in mature Protea sp. and associated food stands as well as nesting sites (Lee and Barnard Reference Lee and Barnard2014). Several species with fynbos and grassland affinity appear to be faring poorly, including Black Harrier Circus maurus, Ground Woodpecker and Grey-winged Francolin. By contrast, Cape Bulbul Pycnonotus capensis, Victorin’s Warbler Cryptillas victorini and Cape Grassbird Sphenoeacus afer show positive population change trends.
Range
The area encompassed by grid cells best fits the IUCN definition of Extent of Occurrence (EOO), defined as that area that can measured by a minimum convex polygon and which contains all sites of occurrence. However, for species with fragmented range due to poor coverage, this area would currently under-represent the technical definition of EOO. Species with EOO < 20,000 km2 may qualify for endangered status if this range is also severely fragmented combined with continuing observed decline in population metrics or extreme population fluctuations. The two species meeting these criteria are Rudd’s and Botha’s Larks, both currently classified as ‘Endangered’.
Range connectivity
Species with small, highly fragmented ranges are traditionally those species most at risk from a conservation perspective (Bolger et al. Reference Bolger, Alberts and Soule1991). Sclater’s Lark, Cinnamon-breasted Warbler and Black-eared Sparrow-lark are three arid-zone species with low scores. The scores of arid zone specialists may be influenced by poor coverage in the arid western and interior of South Africa. On the other hand, forest species like Chorister Robin-chat Cossypha dichroa, Forest Buzzard Buteo trizonatus and Knysna Warbler would be expected to have a fragmented distribution as Afromontane forest is a naturally fragmented biome in South Africa. The low connectivity for the upland or grassland species Yellow-breasted Pipit Anthus chloris and Sentinel Rock-Thrush is unexpected give the extent of their respective preferred biomes.
Conclusions
We have shown how species atlas datasets, using the example of the Southern African Bird Atlas Project, can be used to extract simple population metrics for use in developing conservation status assessments, even where detailed research on species is unavailable. Our set of southern African endemic and near-endemic bird species shows evidence for population declines among several species. This fits the global pattern that range-restricted species are more vulnerable to patterns of global change. Across southern Africa there is concern that range-restricted species will increasingly have no climate envelope space in which to move (Huntley et al. Reference Huntley, Altwegg, Barnard, Collingham and Hole2012).
While we concentrate on southern African endemic bird species, this region hosts considerable populations of bird species with global conservation status that we have not considered in this assessment, such as Blue Swallow Hirundo atrocaerulea, Bearded Vulture Gypaetus barbatus and several crane species. While some of the techniques introduced here, such as standardised population change in conjunction with z-scores can be used for these species, there are further caveats to the interpretation of these species, as movements and range changes elsewhere are difficult to account for.
In order to interpret atlas data, greater use should be made of statistics that standardise reporting rates over as wide an area as possible, and greater use should be made of occupancy modelling that accounts for various issues related to detection arising from observer and seasonal affects (e.g. Bled et al. Reference Bled, Nichols and Altwegg2013), for species with sufficiently large ranges. The metrics we used in this study rely on the assumption that the probability of detecting a species at a site is dominated by its local abundance and otherwise reasonably constant (Guillera-Arroita et al. Reference Guillera-Arroita, Lahoz-Monfort, Elith, Gordon, Kujala, Lentini, McCarthy, Tingley and Wintle2015). In contrast, occupancy- and related models allow for modelling the observation process in more detail (Altwegg et al. Reference Altwegg, Wheeler and Erni2008, MacKenzie et al. Reference MacKenzie, Nichols, Royle, Pollock, Bailey and Hines2006, Royle and Nichols Reference Royle and Nichols2003). However, for species with small ranges compared to the spatial sampling unit, statistically separating the observation process from the biological process can be challenging. In these cases, we argue that comparisons based on raw data can still be useful, provided that they are interpreted with appropriate care. In spite of the difficulties in interpretation of changes in reporting rates between SABAP1 and SABAP2, it is likely that if the SABAP2 results for a species shows decreased reporting rates (or complete absence) over large parts of its range, this reflects genuine range change, as comparisons are more likely to be conservative than to exaggerate increases or decreases (Loftie-Eaton Reference Loftie-Eaton2014). With SABAP2 entering its seventh year with consistent reporting for the last five years, this project will in the near future provide information on population change in its own right. There is thus every reason to continue to encourage the citizen scientists who collect these data to continue doing so, and thus add value to one of Africa’s largest, and certainly most accessible and vibrant biodiversity databases.
Supplementary Material
To view supplementary material for this article, please visit https://doi.org/10.1017/S0959270916000307
Acknowledgements
Our profound thanks to the many citizen scientists who together have submitted more than 13 million records of bird distribution to SABAP1 and 2. This manuscript would not have been possible without the work of Les Underhill, Doug Harebottle, Michael Brooks and the SABAP2 team, and is part of the considerable evidence of the value of these contributions. Ernst Retief commented on a draft version of this paper. Funding for this study was received from Birdlife South Africa, the Percy FitzPatrick Institute of African Ornithology, and National Research Foundation (NRF) grants to Res Altwegg and to Phoebe Barnard. The NRF accepts no liability for opinions, findings and conclusions or recommendations expressed in this publication.








