Inflammatory bowel disease (IBD) is a condition with uncertain aetiology. However, recent studies have determined that factors such as genetic disposition, stress and diet are involved in disease pathogenesis(Reference Innis, Pinsk and Jacobson1). As many as one in 100 individuals in Europe suffer one or more often prolonged episodes of IBD in their lifetime(Reference Lakatos2). In normal intestinal epithelial cells, equilibrium between tolerance to microflora and immune response to pathogenic bacteria is constantly maintained. In these circumstances cytokines play an essential role in gut homeostasis(Reference Monteleone, Peluso, Fina, Caruso, Andrei, Tosti and Pallone3). Evidence suggests that dysregulation of this fine balance leads to the chronic inflammatory state that is characteristic of IBD(Reference Caradonna, Amati, Magrone, Pellagrino, Jirillo and Caccavo4). Destruction of the intestinal mucosa in IBD is associated with amplified production of several key pro-inflammatory cytokines including TNF-α, IL-12 and IL-1β(Reference Bouma and Strober5). TNF-α secretion leads to activation of further cytokine production by macrophages and in addition can act directly on intestinal epithelial cells initiating the changes in epithelial permeability and ion transport that lead to mucosal inflammation and lesions(Reference Marano, Lewis, Garulacan, Soler and Mullin6–Reference Satsu, Ishimoto, Nakano, Mochizuki, Iwanaga and Shimizu8).
PPAR nuclear hormone receptors are well-characterised lipid-activated transcription factors. There are three PPAR isoforms: PPARα which is expressed in the liver, heart, intestine and kidney and plays a role in fatty acid oxidation and inflammation; PPARγ which is expressed in the adipose tissue, colon, immune cells and adrenal gland and plays a role in inflammation and fatty acid metabolism; PPARβ/δ which is expressed in a wide range of tissues but whose function is poorly defined(Reference Yu, Correll and Vanden Heuvel9–Reference Kersten, Desvergne and Wahli11). Lipids bind and activate PPAR in the intestine and mediate anti-inflammatory effects by antagonising the actions of NF-κB, a pro-inflammatory transcription factor, leading to down regulation of pro-inflammatory markers including IL-1β, TNF-α and IL-12 and up regulation of pro-inflammatory cytokines such as IL-10(Reference Yu, Correll and Vanden Heuvel9, Reference Loscher, Draper, Leavy, Kelleher, Mills and Roche12).
Conjugated linoleic acid (CLA) refers to the positional and geometric isomers of linoleic acid. Research indicates that cis-9, trans-11-CLA (c9, t11-CLA) may have anti-inflammatory effects in a range of pro-inflammatory conditions including IBD(Reference Loscher, Draper, Leavy, Kelleher, Mills and Roche12–Reference Hontecillas, Wannemeulher, Zimmerman, Hutto, Wilson, Ahn and Bassaganya-Riera14). Our group recently demonstrated, that c9, t11-CLA suppresses NF-κB activation in dendritic cells(Reference Loscher, Draper, Leavy, Kelleher, Mills and Roche12). Also, a pig feeding study demonstrated that dietary CLA delayed the onset of IBD(Reference Hontecillas, Wannemeulher, Zimmerman, Hutto, Wilson, Ahn and Bassaganya-Riera14, Reference Bassaganya-Riera and Hontecillas15). It is thought that CLA mediates its anti-inflammatory effects by suppressing arachidonic acid-mediated eicosanoid production and/or acting as a PPAR ligand. Dietary CLA is mainly derived from beef and dairy produce. The predominant dietary isomer c9, t11-CLA accounts for approximately 90 % of total CLA(Reference Riserus, Vessby, Arnlov and Basu16) and is synthesised from trans-vaccenic acid (TVA), the predominant trans fatty acid in milk fat and meat, through the action of stearoyl-CoA desaturase (SCD)(Reference Griinari, Corl, Lacy, Chouinard, Nurmela and Bauman17). SCD typically converts palmitic acid to palmitoleic acid and also stearic acid to oleic acid(Reference Ntambi18). This action influences cellular functions such as membrane fluidity and consequently SCD is highly expressed in the colonic epithelium(Reference Mahfouz, Valicenti and Holman19) where it could alter cellular fatty acid composition. Our group have previously demonstrated that the rate of conversion of TVA to c9, t11-CLA is inhibited by other dietary fatty acids such as EPA and 3,10-dithia stearic acid in Caco-2 cells(Reference Renaville, Mullen, Moloney, Larondelle, Schneider and Roche20). The objective of the present study was to determine the bioconversion of TVA into c9, t11-CLA in Caco-2 cells, a well-characterised cell model of the intestinal epithelium. Furthermore the study aimed to compare the relative effects of TVA and c9, t11-CLA on markers of inflammation and lipid metabolism, within the context of IBD in the Caco-2 intestinal epithelial cell model.
Materials and methods
Cell culture
The human colon carcinoma cell line (Caco-2) was purchased from the European Collection of Animal Cell Cultures (ECACC, Salisbury, Wilts, UK). Caco-2 was grown from passage 50–57. Cells were maintained in Dulbecco's modified Eagle's media (Gibco BRL, Paisley, Renfrewshire, UK) supplemented with l-glutamine (2 mm; Sigma-Aldrich, Poole, Dorset, UK), penicillin (100 μg/ml; Sigma-Aldrich), streptomycin (100 μg/ml; Sigma-Aldrich) and fetal calf serum (10 %; Gibco BRL) at 37°C in a humidified 5 % CO2 atmosphere. Cells were split using trypsin-EDTA solution (Sigma-Aldrich). After reaching confluence, cells were seeded at a density of 5 × 105 and grown for 14 d in the presence of fatty acids. c-9, t-11-CLA (Cayman Chemicals) and TVA (Sigma-Aldrich) were dissolved in dimethyl sulfoxide (DMSO; Sigma-Aldrich) and added to cells at a concentration of 50 μm. Media was changed and supplemented with fatty acids or DMSO vehicle control once per d during the course of growth and development. Cell passages were used between 52 and 53. On day 7 of culture, relevant cells were treated with lipopolysaccharide (LPS) at 0·1 mg/ml (Escherichia coli 127:B8; Sigma-Aldrich) to induce inflammation. Supernatant fractions were removed after 24 h and IL-12p70, TNF-α and IL-6 concentrations were measured using DuoSet ELISA kits (R&D Systems, Abingdon, Oxon, UK) according to the manufacturer's instructions.
Fatty acid analysis
Lipid was isolated from cells according to the Bligh & Dyer method(Reference Bligh and Dyer21).
Cells were washed in cold PBS and pelleted for 5 min at 800 g before lipid extraction. Acid methylation induces isomerisation of CLA in a time- and temperature-dependent manner, therefore the base–acid methylation parameters were optimised as described by Park et al. (Reference Park, Albright, Liu, Storkson, Cook and Pariza22) to minimise isomerisation of cis–trans bonds. The extracted lipid fractions were dried under N2 and then methylated with NaOCH3, which was followed with a 4 % solution of HCl in methanol; this was carried out at 50°C for 20 min. Tricosanoic acid (23 : 0) methyl ester was used as an internal standard. Deionised water saturated with hexane (95:5 water–hexane; v/v) and hexane were added to the fatty acid methyl esters (FAME) and following centrifugation the top layer containing FAME in hexane was removed and transferred to tubes containing approximately 0·75 g Na2SO4. Following centrifugation a sample of the supernatant fraction (500 μl) containing FAME was diluted with hexane. The fatty acids were then separated by GC using a Varian 3800 GC (Varian Instruments) equipped with a CP-Sil 88 capillary column (100 m × 0·25 mm internal diameter, 0·2 μm film thickness; Chrompack, The Netherlands) and a Varian 8400 autosampler. The injector and the flame ionisation detector were kept at constant temperatures of 250 and 260°C, respectively. The column oven temperature was held at 40°C for 2 min, increased at 20°C/min to 80°C and held for 2 min, increased to 160°C at 20°C/min, to 220°C at 4°C/min, and to 240°C at 2°C/min and held for 8 min. The total run time was 43 min, and the carrier gas used was H2.
mRNA expression analysis
Gene expression was determined following 24 h LPS stimulation. Total RNA was extracted from Caco-2 cells using the guanidinium thiocyanate–phenol–chloroform method (TRIZOL Reagent®; Molecular Research Center, Inc., Cincinnati, OH, USA). A quantity of 2 μg of RNA was treated with RNase-free DNase to remove contaminating genomic DNA (DNase I Amplification Grade; Invitrogen, Paisley, UK). Total RNA was then reverse transcribed (High Capacity cDNA Reverse Transcription Kit; Applied Biosystems, Warrington, Cheshire, UK). Reaction without RNA and without reverse transcription enzyme was carried out as a negative control. A pool of cDNA samples was serially diluted 1:2 in Rnase-free water to produce a standard curve for the TaqMan® PCR assay. TaqMan PCR was performed for TNF-α, IL-6, IL-12p35, PPARγ and PPARα using PreDeveloped Assay Reagent Kits. Each 25 μl reaction contained 20 ng cDNA, 2X TaqMan Universal PCR Mastermix, forward and reverse primers and TaqMan probe. All reactions were carried out in duplicate with the ABI 7700 Sequence Detection System (Perkin-Elmer Applied Biosystems) using the following conditions: 50°C for 2 min and 95°C for 10 min followed by forty cycles of 95°C for 15 s and 60°C for 1 min. Target gene mRNA expression was normalised to the internal control glyceraldehyde-3-phosphate dehydrogenase (GAPDH) gene and expressed relative to the vehicle control treatment (DMSO) for the non-LPS-stimulated cells.
Statistics
All experiments were preformed in triplicate. Statistical analysis was performed with DataDesk 6.1 (Data Description, Inc., New York, NY, USA). One-way ANOVA determined significant differences between the three fatty acid treatments. A probability of P < 0·05 was considered statistically significant. Additional analysis focusing on differences between TVA and CLA used an unpaired t test to determine significance between fatty acids. There was no statistical difference between vehicle control (DMSO)-treated cells and cells alone, therefore vehicle control cells were used as the reference treatment.
Results
Cellular fatty acid composition and stearoyl-CoA desaturase activity
The fatty acid composition of TVA- and c9, t11-CLA-treated cells compared with control cells is presented in Table 1. TVA treatment significantly increased Caco-2 cell TVA and c9, t11-CLA levels (P < 0·005). The biotransformation of TVA into CLA was almost equivalent to that achieved with c9, t11-CLA treatment; however, the levels of c9, t11-CLA achieved in TVA-treated cells was significantly lower (P = 0·007). Treatment with TVA and c9, t11-CLA significantly decreased the level of several major fatty acids (16 : 1, 18 : 0, cis-18 : 1n-9 and cis-18 : 1n-11); 14 : 0, 16 : 0 and 18 : 0 were significantly reduced by the TVA treatment only.
Table 1 The effect of trans-vaccenic acid (TVA) (50 μm) and cis-9, trans-11-conjugated linoleic acid (CLA) (50 μm) treatment for 7 d on Caco-2 fatty acid composition and the 18 : 1 and 16 : 1 stearoyl-CoA desaturation index†
(Mean values and standard deviations for three experiments)

ND, not determined.
Mean value is significantly different from that for the dimethyl sulfoxide control:
*P < 0·05,
**P < 0·01 (ANOVA).
† Fatty acids are expressed as mol%; fatty acid species occurring less than 1 mol% in all groups were omitted.
‡ Mean value is significantly different from that for mol% of cis-9, trans-11-CLA within the same treatment group (P < 0·01; Student's t test for unpaired values).
The 18 : 1 and 16 : 1 SCD desaturation ratios, an estimate of SCD activity, calculated as 18 : 1/(18 : 1+18 : 0) and 16 : 1/(16 : 1+16 : 0)(Reference Renaville, Mullen, Moloney, Larondelle, Schneider and Roche20), respectively, were significantly increased to the same extent by both TVA and c9, t11-CLA compared with the DMSO vehicle control-treated cells.
Cis-9, trans-11-conjugated linoleic acid but not trans-vaccenic acid modulates markers of the inflammatory phenotype
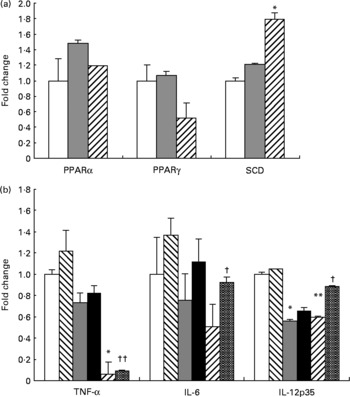
Fig. 1 (b) presents the effect of c9, t11-CLA and its precursor TVA on TNF-α IL-6, and IL-12 mRNA expression. Before LPS stimulation TNF-α and IL-12p35 mRNA levels were significantly down regulated in the c9, t11-CLA-treated cells (P < 0·05, P < 0·01, respectively). TVA-treated cells also had lower pre-LPS IL-12p35 and TNF-α mRNA levels; however, only the former was significant compared with vehicle control cells (P < 0·01). Following 24 h LPS stimulation TNF-α, IL-6 and IL-12p35 mRNA expression was significantly reduced in the c9, t11-CLA-treated cells compared with control cells (P < 0·01, P < 0·05 and P < 0·05, respectively). In contrast, TVA did not show an equivalent anti-inflammatory effect, despite significant conversion to c9, t11-CLA, whereby TVA only significantly reduced IL-12p35 mRNA levels (P < 0·05).

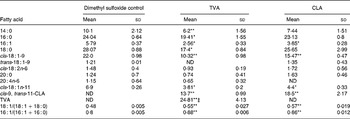
Fig. 1 Effects of trans-vaccenic acid (TVA) and cis-9, trans-11-conjugated linoleic acid (c9, t11-CLA) on inflammatory and lipid marker mRNA expression in Caco-2 cells. Expression of PPARα, PPARγ and stearoyl-CoA desaturation (SCD) (a), and TNF-α, IL-6 and IL-12p35 (b) was measured using Taqman real-time PCR. mRNA levels were normalised to glyceraldehyde-3-phosphate dehydrogenase and results were expressed as fold induction relative to dimethyl sulfoxide vehicle control for unstimulated cells (0 h). (□), Control; (![]() ), TVA; (
), TVA; (![]() ), CLA; (
), CLA; (![]() ) control+lipopolysaccharide (LPS); (■), TVA+LPS; (
) control+lipopolysaccharide (LPS); (■), TVA+LPS; (![]() ), CLA+LPS. Data are the mean values for three experiments, with standard deviations represented by vertical bars. ANOVA was used to compare TVA- and c9, t11-CLA-treated cells with control. Mean value is significantly different from that for control: *P < 0·05, **P < 0·01. Mean value is significantly different from that for control+LPS: † P < 0·05, †† P < 0·01.
), CLA+LPS. Data are the mean values for three experiments, with standard deviations represented by vertical bars. ANOVA was used to compare TVA- and c9, t11-CLA-treated cells with control. Mean value is significantly different from that for control: *P < 0·05, **P < 0·01. Mean value is significantly different from that for control+LPS: † P < 0·05, †† P < 0·01.
IL-12p70 cytokine concentrations were significantly reduced in c9, t11-CLA-treated Caco-2 cells compared with control cells (8·25 (sd 0·42) pg/ml and 9·12 (sd 0·32) pg/ml, respectively) (P < 0·05). TVA-treated Caco-2 cells did not produce significantly less IL-12p70 (8·9 (sd 0·52) pg/ml). Neither fatty acid treatment had a significant effect on IL-1β or TNF-α concentration (data not shown).
Cis-9, trans-11-CLA but not trans-vaccenic acid modulated markers of lipid metabolism in Caco-2 cells
The differential effect of TVA and c9, t11-CLA on molecular markers of lipid metabolism is presented in (Fig. 1 (a)). SCD mRNA levels were significantly increased in c9, t11-CLA-treated cells (P < 0·05), but not TVA-treated cells, compared with control Caco-2 cells. Whilst PPARα and PPARγ are important biomarkers of lipid metabolism that also modulate inflammation, mRNA was not significantly altered by either fatty acid intervention.
Discussion
CLA is a naturally occurring fatty acid with at least eight different isomers(Reference Eulitz, Yurawecz, Sehat, Fritsche, Roach, Mossoba, Kramer, Adlof and Ku23). c9, t11-CLA is the predominant isomer found in meat, milk and other food products from ruminant animals. Anti-inflammatory effects have been demonstrated for this isomer in a range of inflammatory and autoimmune disorders(Reference Kersten, Desvergne and Wahli11, Reference Ringseis, Gahler, Herter and Eder24, Reference Toomey, McMonagle and Roche25). However, relatively little is known about the biopotency of its precursor TVA, despite it being the predominant trans fatty acid in CLA-rich food sources(Reference Riserus, Vessby, Arnlov and Basu16). The occurrence of pro-inflammatory disorders is becoming increasingly common and is set to increase dramatically over the next decade(Reference Logan26). Negative side effects observed with conventional pharmaceutical treatment alongside growing evidence that Western-type diets are implicated in the aetiology of these conditions has led to rising interest in anti-inflammatory fatty acids such as c9, t11-CLA. Current evidence from cell and animal studies suggests that CLA may have a role to play in alleviating symptoms and maintaining remission in patients with IBD(Reference Bassaganya-Riera, Hontecillas and Beitz13, Reference Bassaganya-Riera, Reynolds, Martino-Catt, Cui, Hennighausen, Gonzalez, Rohrer, Benninghoff and Hontecillas27). In the present study we demonstrated that CLA attenuates markers of inflammation relevant to IBD. We observed that whilst there was significant production of c9, t11-CLA in Caco-2 cells supplied with TVA, therefore indicating that TVA was endogenously converted to c9, t11-CLA, this was at a significantly lower level than Caco-2 cells treated with c9, t11-CLA. Although there were indications in the data that TVA-treated cells followed the same anti-inflammatory trend as c9, t11-CLA-treated cells by lowering levels of several pro-inflammatory cytokines, these results were not significant. Therefore, TVA was not associated with the profound anti-inflammatory profile seen in c9, t11-CLA-treated cells. As the present results showed that some of the TVA was converted to c9, t11-CLA, the activity of SCD was estimated by calculating desaturation indices. Both TVA and c9, t11-CLA increased SCD desaturation indices in Caco-2 cells. Although there is little evidence to suggest that c9, t11-CLA affects SCD activity in intestinal cells, the present study demonstrates that this isomer has positive effects on both SCD mRNA levels and SCD desaturation indices. This increase may lead to a beneficial alteration of the SFA:unsaturated fatty acid ratio, which is important in regulating membrane fluidity. Abnormalities of this ratio have been implicated in a range of immune disorders(Reference Li, Birdwell and Whelan28).
Inflammatory cytokines such as TNF-α and IL-12p70 play a crucial role in the pathogenesis of IBD, therefore reduction in their expression and production by anti-inflammatory lipids could represent a viable treatment option. Results show that treatment with c9, t11-CLA significantly reduced TNF-α, IL-6 and IL-12p35 mRNA levels and IL-12p70 concentrations in Caco-2 cells. In contrast, TVA treatment did not have such potential as an anti-inflammatory fatty acid; it did reduce IL-12p35 mRNA levels but this was not associated with a significant effect on Caco-2 cell IL-12p70 concentrations. Based on previous work we would assume that the anti-inflammatory effect of c9, t11-CLA was mediated via NF-κB- and/or PPARγ-dependent mechanism(s) rather than at the level of eicosanoid synthesis, as there was no significant change in arachidonic acid levels following Caco-2 cell CLA treatment(Reference Loscher, Draper, Leavy, Kelleher, Mills and Roche12, Reference Hwang, Kundu, Shin, Lee, Lee and Surh29). However, there is evidence from the literature that c9, t11-CLA mediates anti-inflammatory effects independently of NF-κB via down regulation of TNF-α. Given the down regulation of TNF-α mRNA by c9, t11-CLA-treated cells it is likely that this may also be a major factor in the mediation of anti-inflammatory effects by c9, t11-CLA.
In conclusion, the results of the present study demonstrate that the precursor fatty acid TVA does not display the anti-inflammatory effects associated with c9, t11-CLA. In this case c9, t11-CLA treatment reduced Caco-2 cell TNF-α and IL-12 expression. These are up regulated in patients with IBD. This decrease provides evidence that supplementation of dietary CLA, but not TVA, may have positive anti-inflammatory effects relevant to IBD.
Acknowledgements
We thank Aiveen Marron for excellent technical assistance. The present study was funded by the Food Institutional Research Measure (project no. 5254; Department of Agriculture and Food, Dublin, Republic of Ireland). The authors have no financial conflict of interest.