






Crossref Citations
This article has been cited by the following publications. This list is generated based on data provided by Crossref.
Tu, Bingjie
Zhang, Qingying
Liu, Xiaobing
Yu, Shaopeng
Xu, Nan
Liu, Jia
and
Liu, Changkai
2025.
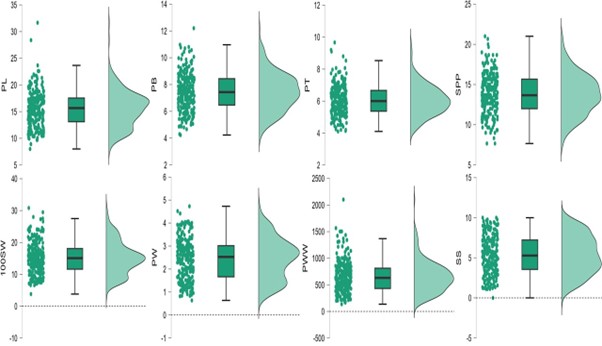
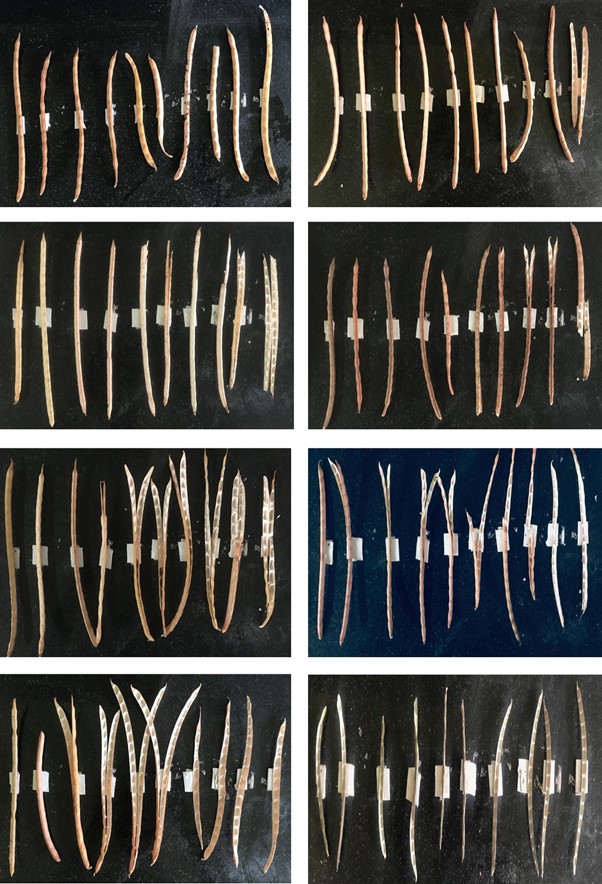
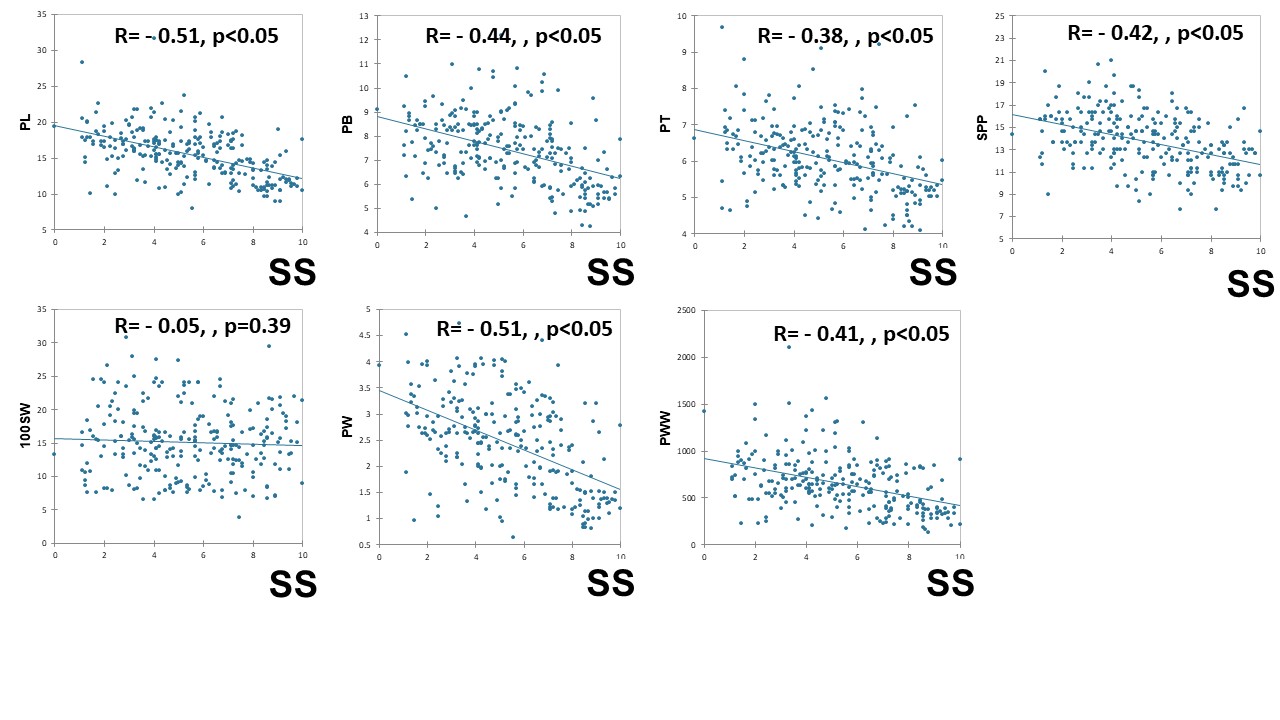
Agronomic and pod traits in relation to pod shattering in cultivated soybeans.
Czech Journal of Genetics and Plant Breeding,
Vol. 61,
Issue. 2,
p.
67.