



Causes of cancer
Replication errors
Cancer is a genetic disease in the sense that it is induced by modification of genetic information (the nucleotide sequence) in genes involved in regulating cell proliferation resulting in uncontrolled cell proliferation, a landmark of tumour. Since cell proliferation is tightly regulated by both negative and positive controls, modification of more than two genes is usually required for tumour formation. Furthermore, cancer is a malignant tumour with potential of invasion into healthy tissue and metastasis. Modification of several other genes such as genes involved in apoptosis, metabolism, cell–cell contact, and so on, is also required for rapid proliferation, metastasis and escape from immune response (Fig. 1).

Fig. 1. Schematic view of available strategies for therapeutic targeting of the various ‘hallmarks of cancer’ with drugs that interfere with each of the acquired capabilities necessary for tumour growth and progression. Adapted with permission from Hanahan and Weinberg (Reference Hanahan and Weinberg2012).
Modification could be caused by replication errors; this is, however, very rare because the high fidelity of DNA replication is maintained by DNA polymerase itself, its proofreading functions and other surveillance systems such as mismatch-repair systems (Kunkel, Reference Kunkel2004; Bębenek and Graczyk, Reference Bębenek and Graczyk2018). Therefore, modification of particular sets of several genes in one cell needs accumulation of errors upon many cell divisions over the course of a lifespan. Consequently, older people are more prone to get cancer. However, such events are still very rare under normal conditions. Loss of replication fidelity due to inherited and sporadic mutations in replication error repair systems (Bębenek and Graczyk, Reference Bębenek and Graczyk2018) is a prerequisite for cancer formation. Individuals with inherited mutations in mismatch repair enzymes seem prone to developing colorectal cancer. Another cause that increases replication errors is DNA damage as we will touch upon below.
DNA damage and error-prone repair
DNA damage by exogenous and endogenous agents can result in mutation (De Bont and van Larebeke, Reference De Bont and Larebeke N2004; Tubbs and Nussenzweig, Reference Tubbs and Nussenzweig2017). DNA is chemically fragile; every minute in every cell, it undergoes various chemical modifications, including oxidation, alkylation, deamination, elimination of nucleobases and disruption of the phosphate backbone (De Bont and van Larebeke, Reference De Bont and Larebeke N2004). Some modifications, such as 8-oxoguanine (8-oxoG) and O-6 methyl guanine (O6-methylG), increase the likelihood of replication error by forming non-canonical base pairs (8-oxoG with A and O6-methylG with T). Other modifications are bypassed by less accurate DNA polymerases (translesion replication), promoting mutation (Sale, Reference Sale2013).
Cells harbour multiple redundant DNA repair systems that help them avoid forming tumours (Sale, Reference Sale2013). DNA damage that occurs on one of the two DNA strands can be repaired using the undamaged complementary strand as a template. By contrast, double-strand breaks cannot be fixed in this manner and are instead repaired by non-homologous end-joining (NHEJ), microhomology-mediated end-joining or homologous recombination (HR) (Jasin and Rothstein, Reference Jasin and Rothstein2013; Wright et al., Reference Wright, Shah and Heyer2018). NHEJ is mutagenic, whereas HR is comparatively error-free (Tubbs and Nussenzweig, Reference Tubbs and Nussenzweig2017). Inherited and sporadic defects in repair enzymes predispose carriers to cancer formation, and such mutations are observed in many cancers (Tubbs and Nussenzweig, Reference Tubbs and Nussenzweig2017). Behaviours that cause accumulation of DNA damage, such as smoking or excessive sun exposure, also increase cancer risk.
HR: double-edged roles
HR involves exchange of strands between two homologous DNAs, and its main step is catalysed by Rad51 (Jasin and Rothstein, Reference Jasin and Rothstein2013; Wright et al., Reference Wright, Shah and Heyer2018). A schematic presentation of the Rad51-promoted strand-exchange mechanism is given in Fig. 2. By exchanging a broken strand for an undamaged homologous strand, HR can repair double-strand breaks usually without causing any genetic modification and, in this way, prevent mutation. HR also restores a stalled replication fork due to replication error or non-treated DNA damage. However, the role of HR in preventing cancer was underestimated until the discovery that BRCA2, the mutation of which predisposes to breast and ovarian cancers with high penetrance (Prakash et al., Reference Prakash, Zhang, Feng and Jasin2015; Heeke et al., Reference Heeke, Pishvaian, Lynce, Xiu, JR, Chen, Baker, Marshall and Isaacs2018), is involved in HR by direct interaction with Rad51 (Mizuta et al., Reference Mizuta, LaSalle, Cheng, Shinohara, Ogawa, Copeland, Jenkins, Lalande and Alt1997).
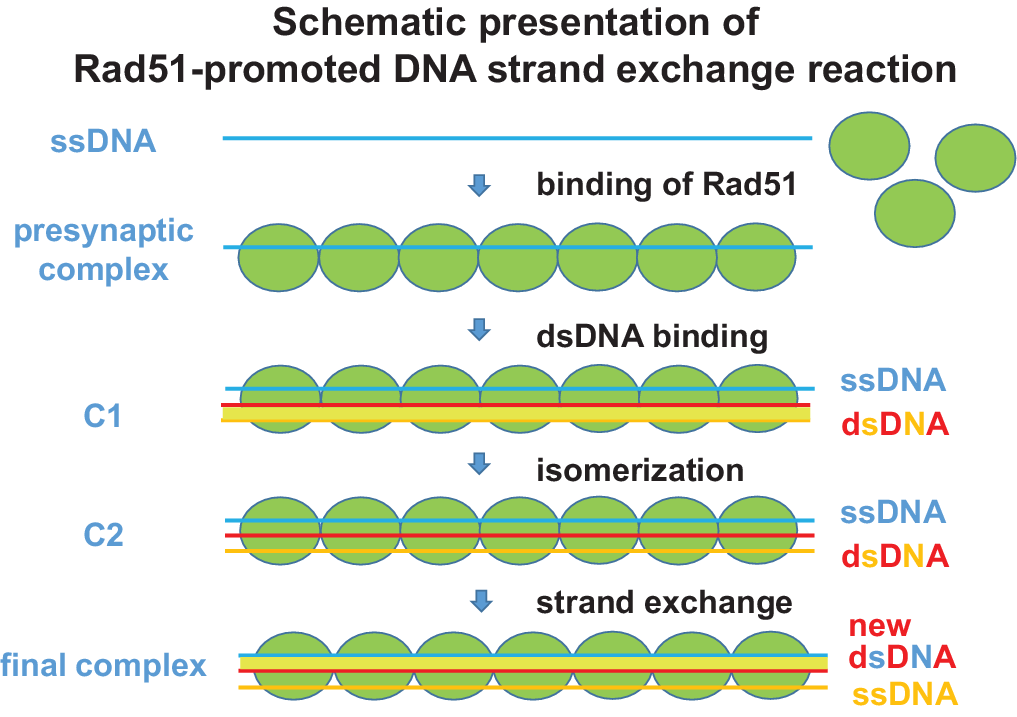
Fig. 2. Schematic presentation of Rad51-promoted DNA strand-exchange reaction. Reaction steps observed by kinetic analysis (Ito et al., Reference Ito, Murayama, Takahashi and Iwasaki2018) are shown. Rad51 molecules (green balls) bind to single-stranded DNA (blue line) to form a presynaptic filament. The filament binds a double-stranded DNA (red/orange lines) to form complex C1 and changes the DNA conformation (complex C1) before complete strand exchange. For simplicity, helical shapes of Rad51 filament and double-stranded DNA are not shown. Structures of C1 and C2 complexes are yet unknown.
On the other hand, strand exchange can promote loss of heterozygosity and aberrant genetic arrangement (Godin et al., Reference Godin, Sullivan and Bernstein2016). In our cells, most genes exist in pairs, one inherited from each parent. Mutations are rare and usually affect only one copy of a gene, and it is likely that any given mutation will be masked by the presence of the ‘healthy’ gene, with the exception of dominant phenotypes such as predisposition to breast cancer caused by BRCA2 mutation. Loss of heterozygosity causes the mutant phenotype to become manifest. Thus, HR can in this way also contribute to cancer formation (Godin et al., Reference Godin, Sullivan and Bernstein2016). HR may also occur between two non-identical but similar DNA parts. This promotes deletion of DNA, when two parts are on the same DNA and chromosome translocation when two parts are on two different chromosomes (Bishop and Schiestl, Reference AJR and Schiestl2002). These exchanges can occur in human because the human genome is so large that there can be many similar DNA sequences. Furthermore, the human genome contains many repetitive sequences. Therefore ‘erroneous’ HR may be a cause of cancer formation. In Fig. 3, the origin of the double-edged roles of Rad51 is schematically presented. Obviously, it is important to understand in detail the recognition mechanism of homologous DNA and its accuracy. Such knowledge would be also useful for the application of the CRISPR technique, which relies on accuracy of HR to incorporate external DNA at an appropriate site in chromosome.

Fig. 3. The double-edged roles of Rad51-mediated homologous DNA recombination and associated normal, benign processes in an organism (blue) and below (in red) error-associated processes potentially connected to tumour formation.
Rad51 and cancer
Basic functions of Rad51
Rad51 is a member of the RecA family of proteins, which also includes RecA, RadA and Dmc1. It plays a key role in HR: searching out and pairing homologous DNA sequences and then promoting strand exchange between them (Fig. 2). Rad51 function in cells requires several accessory proteins (Bishop and Schiestl, Reference AJR and Schiestl2002; Carreira and Kowalczykowski, Reference Carreira and Kowalczykowski2011; Prakash et al., Reference Prakash, Zhang, Feng and Jasin2015; Heeke et al., Reference Heeke, Pishvaian, Lynce, Xiu, JR, Chen, Baker, Marshall and Isaacs2018; Jia et al., Reference Jia, Song, Dong, Hao, Zhao, Li and Tong2019), and its activity is regulated by the interactions with regulatory proteins and post-translational modifications. Rad51 is phosphorylated by cancer-related kinases (Daboussi et al., Reference Daboussi, Dumay, Delaco and Lopez2002; Chabot et al., Reference Chabot, Defontaine, Marquis, Renodon-Corniere, Courtois, Fleury and Cheraud2019). Hypoxia in cancer cells is reported to downregulate Rad51 (Bindra et al., Reference Bindra, Schaffer, Meng, Woo, Måseide, Roth, Lizardi, Hedley, Bristow and Glazer2004). In addition to defects in the accessory proteins of Rad51 involved in HR, which are frequently observed in cancer cells, alterations of Rad51 itself have also been observed in cancer patients (Antoniou et al., Reference Antoniou, Spurdle, Sinilnikova, Healey, Pooley, Schmutzler, Versmold, Engel, Meindl, Arnold, Hofmann, Sutter, Niederacher, Deissler, Caldes, Kämpjärvi, Nevanlinna, Simard, Beesley, Chen, Neuhausen, Rebbeck, Wagner, Lynch, Isaacs, Weitzel, Ganz, Daly, Tomlinson, Olopade, Blum, Couch, Peterlongo, Manoukian, Barile, Radice, Szabo, Pereira, Greene, Rennert, Lejbkowicz, Barnett-Griness, Andrulis, Ozcelik, Gerdes, Caligo, Laitman, Kaufman, Milgrom, Friedman, Domchek, Nathanson, Osorio, Llort, Milne, Benítez, Hamann, Hogervorst, Manders, Ligtenberg, Ouweland, Peock, Cook, Platte, Evans, Eeles, Pichert, Chu, Eccles, Davidson, Douglas, Godwin, Barjhoux, Mazoyer, Sobol, Bourdon, Eisinger, Chompret, Capoulade, Bressac-de Paillerets, Lenoir, Gauthier-Villars, Houdayer, Stoppa-Lyonnet, Chenevix-Trench and Easton2008).
Rad51 antagonises radio- and chemo-therapies of cancer
Rad51 is thought to protect cancer cells against radio- and chemo-therapies by repairing DNA damage induced by these treatments – ‘Rad’ derived from radiation (Prakash et al., Reference Prakash, Zhang, Feng and Jasin2015). Consequently, cancers with defects in HR are more efficiently treated by such therapeutic modalities. In fact, inhibition or downregulation of Rad51 increases the efficiency of radiotherapy and chemotherapy (Tsai et al., Reference Tsai, Kuo, Chiu, Y-C and Lin2010). In this context, many small molecules that could act as inhibitors of Rad51 activity are worth investigating (Huang and Mazin, Reference Huang and Mazin2014; Chen et al., Reference Chen, Cai, Li and Wu2017).
Rad51 promotes cancer and cancer progression
When overexpressed, Rad51 can also promote cancer, and hyperactive Rad51 mutations have been detected in the cells of solid tumours. Multiple studies have reported Rad51 overexpression in various cancers (Godin et al., Reference Godin, Sullivan and Bernstein2016). Furthermore, these studies show that elevated Rad51 expression is correlated with reduced patient survival. Thus, Rad51 promotes progression of cancer, increasing its malignancy by stimulating metastasis and increasing resistance to chemotherapy. Accordingly, Rad51 is considered promising as a target for treatment of cervical carcinoma, breast cancer and non-small-cell lung cancer (Tsai et al., Reference Tsai, Kuo, Chiu, Y-C and Lin2010; Huang and Mazin, Reference Huang and Mazin2014; Chen et al., Reference Chen, Cai, Li and Wu2017; Jia et al., Reference Jia, Song, Dong, Hao, Zhao, Li and Tong2019).
Many cancers have epigenetic deficiencies in various DNA repair genes, likely resulting in higher rates of unrepaired DNA damage. The overexpression of Rad51 seen in many cancers may reflect compensatory Rad51 overexpression (as in BRCA1 deficiency) and elevated rates of HR repair to deal at least partially with excess DNA damage.
Challenge of understanding Rad51-promoted DNA strand-exchange reaction
Appropriate inhibition of Rad51
As noted above, Rad51 has been proposed as a target for cancer treatment. Indeed, some Rad51 inhibitors are effective against some cancers. However, because of the double-edged character of Rad51, inhibition of its activity may also have negative effects, for example, promoting more malignant cancers. Many currently available Rad51 inhibitors prevent formation of Rad51 filament on DNA, the first step of the reaction (Huang and Mazin, Reference Huang and Mazin2014; Chen et al., Reference Chen, Cai, Li and Wu2017). Consequently, damage then remains unaddressed and may end up being processed by a mutagenic repair system such as NHEJ. Therefore, it would be preferable to inhibit the reaction at an intermediate stage that is toxic to cancer cells. Such toxic Rad51 filaments have been reported in yeast (Veaute et al., Reference Veaute, Jeusset, Soustelle, Kowalczykowski, Le Cam and Fabre2003; Symington and Heyer, Reference Symington and Heyer2006). However, it is necessary to fully understand the reaction mechanism in order to inhibit Rad51 activity at the appropriate step to avoid negative effects.
Rad51 will be crucially needed in future CRISPR-based cancer therapy
Double-strand-break repair by HR is initiated by 5′-to-3′ strand resection; in humans, the DNA nuclease cuts back the 5′ end of one strand to generate a 3′ single-stranded DNA overhang. The development of CRISPR technology for both cancer gene diagnostics and therapeutics (including gene-edited T-cells) will rely on insight into the native cellular repair system. This may represent one of the most important motivations for investigating Rad51 (and RecA, as model system) in cancer research. When Aaron Klug invented zinc finger-based sequence-specific artificial double-strand endonucleases, he noted that a bottleneck in progress toward gene-correction therapy would be the need to entrust insertion of the desired DNA into the native cellular recombination machinery, that is, Rad51 (Deltcheva et al., Reference Deltcheva, Chylinski, Sharma, Gonzales, Chao, Pirzada, Eckert, Vogel and Charpentier2011; Jinek et al., Reference Jinek, Chylinski, Fonfara, Hauer, Doudna and Charpentier2012).
Reaction mechanism
The mechanisms of recombination enzymes appear similar in all organisms (Ito et al., Reference Ito, Murayama, Takahashi and Iwasaki2018; Takahashi, Reference Takahashi2018). In eukaryotes, the Rad51 protein plays a central role in homologous recombinational repair. Specifically, Rad51 catalyses strand transfer between a damaged sequence and its undamaged homologue to allow re-synthesis of the damaged region. In addition, it collaborates with Dmc1, another recombinase, in proper segregation of chromosomes in meiosis. For the reaction, the enzyme binds to a single-stranded region of DNA to form a well-organized filamentous complex with DNA (Fig. 2). This single-stranded (ss) DNA–RecA filament then interacts with a double-stranded (ds) DNA to form an ssDNA–RecA–dsDNA complex, and if the two DNAs have identical or nearly identical sequences, strand exchange occurs. All reaction steps occur in a long nucleoprotein filament.
The structure of ssDNA–RecA filament has been determined by X-ray crystallographic analysis (Chen et al., Reference Chen, Yang and Pavletich2008) and other techniques such as linear dichroism and electron microscopy (Stasiak et al., Reference Stasiak, Di Capua and Koller1981; Norden et al., Reference Norden, Elvingson, Kubista, Sjöberg, Ryberg, Ryberg, Mortensen and Takahashi1992). The structure of ssDNA–Rad51 filament seems similar (Xu et al., Reference Xu, Zhao, Xu, Zhao, Sung and Wang2017). In the filament, Rad51 monomers are arranged in a helical manner around ssDNA, which also forms helix and ready to receive dsDNA (Reymer et al., Reference Reymer, Frykholm, Morimatsu, Takahashi and Nordén2009). DNA is elongated about 50% in length. By contrast, the structure of ssDNA–RecA–dsDNA (and ssDNA–Rad51–dsDNA) complex has not been determined although linear dichroism and modeling indicate a similar protein arrangement (Reymer et al., Reference Reymer, Frykholm, Morimatsu, Takahashi and Nordén2009). Kinetic analysis shows the presence of two ssDNA–Rad51–dsDNA complex intermediates (Fig. 2) (Ito et al., Reference Ito, Murayama, Takahashi and Iwasaki2018). Recognition of sequence homology seems to occur before formation of Watson–Crick base pairing between ssDNA and complementary strand separated from dsDNA (Gupta et al., Reference Gupta, Folta-Stogniew, O’Malley, Takahashi and Radding1999). The reaction can start at any part with six-base homology (Anand et al., Reference Anand, Beach, Li and Haber2017; Takahashi, Reference Takahashi2018).
Despite the great importance of recombinases in the context of human health (e.g. cancer, gene therapy and sterility) and many years of intense research, the mechanisms of homology search and strand exchange are not yet understood at the atomic level. Many questions, including how the Rad51 filament binds a second DNA, recognizes and searches sequence homology and why Rad51 stretches the DNA, remain unanswered. An improved understanding of the mechanistic details of these fundamental processes could hopefully clarify the accuracy of HR and its double-edged roles in both cancer prevention and formation. Such knowledge will also be useful for the CRISPR technology, in which incorporation of new DNA relies on the cell’s native recombination machinery.
The lack of breakthroughs in HR research could have several explanations. One possibility could be the fact that the reaction differs from many other enzymatic reactions in terms of substrate size. Rad51 exchanges strands of long DNA segments, and many Rad51 molecules must assemble into a very long filament to catalyse the reaction. Consequently, elucidating reaction mechanisms is challenging. Furthermore, Rad51 interacts with two DNA molecules to promote strand exchange between them. The details of how Rad51 interacts with these DNAs, especially the second one, and associated topological challenges, remain elusive. For example, we do not know how the second DNA enters the filament to interact with the first DNA, which is completely coated by Rad51. Similarly, it remains unclear how Rad51 separates the two strands of double-stranded DNA to promote strand exchange.
Contributions of other factors than hydrogen bonds to DNA base pair recognition
A provocative possibility is that our current model of DNA recognition based on hydrogen bonded Watson–Crick base pairs is somehow incomplete, requiring some new theoretical approach to guide more systematic experiments. Here key factors including hydrophobic catalytic effects may have been overlooked. Conventionally, we imagine that the HR in DNA involves the formation of base pairs mediated by hydrogen bonds between two strands. However, hydrogen bonding alone cannot explain the discrimination of DNA with few mismatches relative to a completely complementary strand. Both Rad51 and RecA initiate the HR reaction when the two DNAs present sequence identity over as many as six to eight contiguous bases (Anand et al., Reference Anand, Beach, Li and Haber2017; Takahashi, Reference Takahashi2018). The energetic difference between binding of DNA with one mismatch and with a completely matched DNA is too small (less than 20%) if we consider only hydrogen bond formation between bases. Still RecA manages to eliminate such a singly mismatched DNA in more than 80% of the cases (Takahashi, Reference Takahashi2018). This may be because the selection is not made thermodynamically but kinetically (Bazemore et al., Reference Bazemore, Folta-Stogniew, Takahashi and Radding1997; Takahashi, Reference Takahashi2018). A matched DNA binds to the Rad51/DNA filament much faster than one containing a mismatch, as has also been observed in hybridization contexts (Jensen et al., Reference Jensen, Orum, Nielsen and Nordén1997). This is apparently also the case with the selection of complementary base by DNA polymerase during replication (Oertell et al., Reference Oertell, Harcourt, Mohsen, Petruska, Kool and Goodman2016). However, the hydrogen bonds become rapidly weaker with distance, and it is difficult to imagine that hydrogen bonds alone could play a decisive role in the kinetic selection process. We here propose that other factors be involved.
We know today that DNA polymerase selects the correct DNA base without requiring complete hydrogen bond formation (Kool, Reference Kool2000), and that certain modified DNA bases that cannot form hydrogen bonds with a partner are still selected by polymerases. Moreover, several studies have reported artificial base pairs involving non-canonical bases that do not form any hydrogen bonds with their partners (Henry and Romesberg, Reference Henry and Romesberg2003; Yamashige et al., Reference Yamashige, Kimoto, Takezawa, Sato, Mitsui, Yokoyama and Hirao2012), and that DNA containing such a third base pair (in addition to A–T and G–C pairs) can still be correctly replicated without significant error. In these base pairs, geometrical complementarity of two bases appear to be important (Yamashige et al., Reference Yamashige, Kimoto, Takezawa, Sato, Mitsui, Yokoyama and Hirao2012). For such a recognition mechanism, also the correct mutual orientation of the two bases is crucial. This orientation effect may be the result of a precise steric guidance due to base stacking with adjacent bases. A similar mechanism, we propose, may be involved in the Rad51-promoted DNA recognition and strand exchange.
The stability of double-stranded DNA is supported mainly by base stacking between neighbouring bases, rather than by formation of A–T and G–C base pairs (Yakovchuk et al., Reference Yakovchuk, Protozanova and Kamenetskii2006). We recently demonstrated that the semi-hydrophobic environment created by ethylene glycol ethers, such as poly-ethylene glycol (PEG), significantly weakens DNA base-stacking strength but not the hydrogen bonding, and also promotes the DNA strand-exchange reaction even in the absence of any recombinase protein (Feng et al., Reference Feng, Sosa, Mårtensson, Jiang, Tong, Dorfman, Takahashi, Lincoln, Bustamante, Westerlund and Nordén2019). Interestingly, both RecA and Rad51 elongate DNA by unstacking one base pair of every three, evidencing a weakening of the base-stacking forces (Chen et al., Reference Chen, Yang and Pavletich2008; Xu et al., Reference Xu, Zhao, Xu, Zhao, Sung and Wang2017; Sun et al. Reference Sun, McCorvie, Yates and Zhang2020). Interestingly, we also note that several hydrophobic residues are present in the proximity of the DNA-binding sites of both Rad51 and RecA (the so-called L1 and L2 loops), and, thus, we speculate that these, just like PEG, can provide a hydrophobic environment that may catalyse the strand-exchange reaction.
Perspective
Obviously, Rad51 is involved in causing many, possibly a majority, of our common cancers. At present, however, this is simply a statement of correlation, and the exact causal mechanism is yet unknown. It is obvious that dealing with the genome, either by natural heterozygotic gene mixing, which is the basis of Darwinian evolution, or the repair of random errors is serious business and prone to introduce potentially malignant genetic aberrations. Unsolved questions regarding both the eukaryotic Rad51 and its simpler, more ‘ancestral’ protein RecA in prokaryotes have persisted despite over 40 years of intense research. This may be partly explained by the inherent complexity of the HR reaction, which involves a large number (10–100) of Rad51 protein units bound to a stretch of DNA. However, it is also possible that our understanding of recombination mechanisms remains incomplete. One factor recently proposed to be important for the interactions and stability of DNA is hydrophobic effects in the vicinity of DNA, which may be involved in catalysing crucial steps in the HR reaction (Ito et al., Reference Ito, Murayama, Takahashi and Iwasaki2018) via a destabilized base stacking (Feng et al., Reference Feng, Sosa, Mårtensson, Jiang, Tong, Dorfman, Takahashi, Lincoln, Bustamante, Westerlund and Nordén2019). Such an effect might explain the bewildering observation that in contrast to DNA replication and transcription, where nucleotides are added and tested one by one, homologous DNA recombination through strand exchange is testing whole sequences, six or more bases in length, for complementarity, requiring the elimination of partially mismatched yet thermodynamically stable intermediates. These are just a few of the elusive points; many questions remain regarding mechanism of gene recombination. For these reasons, and in light of the strong connection between cancer and Rad51, we conclude that fundamental research on Rad51 is a prerequisite for progress in cancer research.
Open Peer Review
To view the open peer review materials for this article, please visit http://doi.org/10.1017/qrd.2020.13.



 Open access
Open access
Comments
Comments to Author: Nordén & Takahashi write a Perspective with a thought-provoking title: “Understanding Rad51 function is a prerequisite for progress in cancer research”. Cancer is an extremely complex molecular and cellular phenomenon.From their own point of view, they summarized how Rad51 plays a double-edged role: 1) positive role to protect further DNA damage, repairing it through homologue recombination preventing cancer development, 2) negative role to prevent chemotherapy treatment and participating cancer further development.
In the Abstract and a variation of it later in the Conclusion, they state that “Another, more dramatic reason for lack of understanding could be that our model of DNA interaction based on hydrogen bond-directed base pair formation is somehow fundamentally wrong, mandating a complete overhaul of the way we think of reactions involving DNA. Since Rad51 recognizes sequence identity of DNA over several bases, one base mismatch could be tolerated when we consider only hydrogen bond energy. This is not the case. We propose that certain hydrophobic effects, recently discovered destabilizing stacking of nucleobases may play a central role in this context.”
This reviewer considers their statement very provocative and thought-provoking.But the statement is too strong to be readily accepted by the research community.It is better to state that the current understanding and emphasis of importance of hydrogen bond for conferring DNA stability is incomplete. The accumulated experimental data requires new theoretical insight to guide experiments into how hydrophobic interaction also plays a key role for DNA stability and in homologue recombination.
The importance of hydrogen bond in DNA base pairing is indisputable: i) faithful DNA replication with extremely rare errors, ii) RNA transcription from DNA with very high fidelity and iii) the genetic code for specific tRNA anticodon to recognize the codon mRNA are all require specific hydrogen bond recognition.The base stacking through the hydrophobic interaction in DNA is absolutely critical for DNA stability and hybridization, but hydrophobic interaction cannot confer specificity of base pairing.It is known that DNA hybridization is largely driven by hydrophobic interaction in water, non-aqueous solvent can interfere the hybridization both thermodynamically and kinetically. But for specific base pairing, namely, how each base recognizes its complementary base requires precise hydrogen bond recognition and perfect registration. The author is correct that hydrogen bonds contribute less to the DNA stability and is not the driving force for hybridization.The hydrogen bonds main function is to confer and confirm the base-pairing specificity, which is the essence of high fidelity of replication, transcription and translation.
Rad51 is a very interesting protein and plays many roles in cell. But its negative tole to promote cancer was less widely known. They wrote “When overexpressed Rad51 can also promote cancer, and hyperactive Rad51 mutations have been detected in the cells of solid tumors. Multiple studies have reported Rad51 overexpression in various cancers (Godin et al., 2016). Furthermore, these studies show that elevated Rad51expression is correlated with reduced patient survival.” In light of the latest clinical observations, paying closer attention and intense scrutiny to Rad51’s other role I cancer is required.
In order to truly understand the double-edged role of Rad51, additional experiments are needed including the detailed molecular structure with DNA in action.The latest CryoEM may be a useful tool to address such dauting structural question.But it will likely gain much insights.
A few points to consider:
1) In order to clearly emphasis of the double-edged role of Rad51, this reviewer suggests to add a Figure 3. This figure 3 can be a cartoon to clearly show Rad51 dual function: i) an important positive role in DNA homologue recombination, ii) negative role in participating cancer development, e.g. phosphorylated by cancer-related kinases.
2) On page 5, 2nd paragraph, line 8, “recombination may also occur between two no identical but similar DNA parts”, it is better to use “between two nonidentical but similar DNA parts”.
3) On page 10, 2nd paragraph, it is best to add a subtitle for this section to emphasize author’s points.
4) The statement “A third, more dramatic possibility is that something is wrong with our basic theory of DNA interactions, which may require a complete paradigm overhaul” is too strong.It could be re-written as “A more provocative possibility is that our current knowledge on DNA interaction strongly emphasis on hydrogen bond is incomplete, new theoretical insights to guide more systematic experiments are required. Other key factors including hydrophobic catalysis should not be overlooked.”
5) Page 10, the last paragraph, they state: “Another possibility is that hydrogen bond formation is less important, and that some other element governs selection”.A phrase could be inserted after less important “it may not be the driving force for DNA hybridization and double helix stability”.
6) Page 11, 2nd paragraph, “We must remember that the stability of double-stranded DNA”, “We must remember that” is unnecessary.
7) Page 12, the subtitle “Conclusion”.It is better to use “Perspective”.
8) First line, “Obviously, Rad51 is somehow involved”. It is not better not to use the word “somehow”.Somehow is not science.Science must explain “How”, not “Somehow”.If it is uncertain, the phrase could change to “Rad51 may be involved”.
9) Page 12, line 6, “Mysteries”, it is better to use “Unsolved questions”.
10) Line 9, “that our understanding of recombination mechanisms remains incomplete because something is fundamentally wrong with our basic theory of DNA interactions.” The sentence “because something is fundamentally wrong with our basic theory of DNA interactions” is not necessary since you immediately below proposed your own idea to complete the incomplete theory.
After the authors add Figure 3 to clearly show the double-edged role of Rad51, address these concerns and make careful revision, this reviewer recommends publication of this provocative and thought-provoking Perspective.