

Modern crocodiles, caiman, gharials and alligators are the only living members of Crocodyliformes, a group of pseudosuchian archosaurs whose evolutionary history spans about 200 million years (Whetstone & Whybrow Reference Whetstone and Whybrow1983; Benton & Clark Reference Benton, Clark and Benton1988; Bronzati et al. Reference Bronzati, Montefeltro and Langer2015; Mannion et al. Reference Mannion, Benson, Carrano, Tennant, Judd and Butler2015). The Middle Jurassic was a key period in their evolution, with thalattosuchians rapidly diversifying in the marine realm (e.g., Gasparini and Chong Reference Gasparini and Chong1977; Young et al. Reference Young, Brusatte, Ruta and Andrade2010, Reference Young, Bell, Andrade and Brusatte2011; Wilberg Reference Wilberg2015). However, much less is known about the non-pelagic crocodyliforms from this time. Apparently, terrestrial and freshwater species were stuck in a low point of their diversity, although this could be an artefact of the notoriously poor Middle Jurassic tetrapod fossil record (Tennant et al. Reference Tennant, Mannion and Upchurch2016a). Sparse sampling during this interval complicates our understanding of the diversification patterns of non-pelagic crocodyliforms (Bronzati et al. Reference Bronzati, Montefeltro and Langer2015; Mannion et al. Reference Mannion, Benson, Carrano, Tennant, Judd and Butler2015; Tennant et al. Reference Tennant, Mannion and Upchurch2016b), meaning that new Middle Jurassic fossils are critical.
Indeed, crocodyliforms from terrestrial and nearshore assemblages during the Middle Jurassic remain poorly understood compared with those from other geological intervals. During the Late Jurassic, most non-pelagic crocodyliforms in Europe were goniopholidids and atoposaurids (Andrade et al. Reference Andrade, Edmonds, Benton and Schouten2011; Young et al. Reference Young, Tennant, Brusatte, Challands, Fraser, Clark and Ross2016a; Tennant et al. Reference Tennant, Mannion and Upchurch2016b, Reference Tennant, Mannion and Upchurchc), whereas in other parts of the world, small-bodied sphenosuchian and protosuchian-grade taxa were widely distributed (Reference GaoGao 2001; Clark et al. Reference Clark, Xu, Forster and Wang2004; Göhlich et al. Reference Göhlich, Chiappe, Clark and Sues2005; Pol et al. Reference Pol, Rauhut, Lecuona, Leardi, Xu and Clark2013). By the Early Cretaceous, European faunas were composed of a more diverse assemblage of neosuchians, including goniopholidids, pholidosaurids, bernissartiids, putative atoposaurids and hylaeochampsids (e.g., Salisbury & Naish Reference Salisbury, Naish and Batten2011; Sweetman et al. Reference Sweetman, Pedreira-segade and Vidovic2015). Ghost lineage analyses imply that, depending on the systematic position of the Cretaceous clades, some major neosuchian lineages may have originated during the dark period of the Middle Jurassic (e.g., Bernissartiidae, Hylaeochampsidae, Paralligatoridae and Pholidosauridae: Bronzati et al. Reference Bronzati, Montefeltro and Langer2015; Turner Reference Turner2015; Tennant et al. Reference Tennant, Mannion and Upchurch2016b).
The problem, however, is that there are currently few known places where Middle Jurassic, non-pelagic crocodyliforms can be found. Jurassic fossil-bearing beds of terrestrial, freshwater and lagoonal origins are sparsely distributed in China (Gao Reference Gao2001; Maisch et al. Reference Maisch, Matzke, Pfretzschner, Sun, Stohr and Grossmann2003; Fu et al. Reference Fu, Ming and Peng2005), Kyrgyzstan (Averianov Reference Averianov2000), parts of Europe (Evans & Milner Reference Evans, Milner, Fraser and Sues1994; Kriwet et al. Reference Kriwet, Rauhut and Gloy1997; Knoll et al. Reference Knoll, Cuny, Mojon, López-Antoñanzas and Huguet2013; Knoll & López-Antoñanzas Reference Knoll and López-Antoñanzas2014; Young et al. Reference Young, Tennant, Brusatte, Challands, Fraser, Clark and Ross2016a) and Russia (Kuzmin et al. Reference Kuzmin, Skutschas, Grigorieva and Krasnolutskii2013), which has yielded some crocodyliform fossils. Furthermore, there are undescribed crocodyliform teeth of this age from Thailand (Tong et al. Reference Tong, Buffetaut and Suteethorn2002) and possible remains from Madagascar (Dal Sasso & Pasini Reference Dal Sasso and Pasini2003; Flynn et al. Reference Flynn, Fox, Parrish, Ranivoharimanana, Wyss, Harris, Lucas, Spielmann, Lockley, Milner and Kirkland2006), but these are fragmentary. The vast majority of Middle Jurassic crocodyliform fossils are isolated specimens from microvertebrate localities – limited evidence that makes it difficult to assign these fossils to higher-level clades, which in turn hinders our understanding of the origin and early evolution of these groups that would later go on to great diversity and success in the Cretaceous and beyond.
Here we report a new crocodyliform lower jaw from the Middle Jurassic of the Isle of Skye, Scotland, UK (Fig. 1). This study is part of an ongoing investigation of the world-class Middle Jurassic vertebrate fossil record of Skye, led by the PalAlba Group (Brusatte et al. Reference Brusatte, Young, Challands, Clark, Fischer, Fraser, Liston, Macfadyen, Ross, Walsh and Wilkinson2015, Reference Brusatte, Challands, Ross and Wilkinson2016; Brusatte & Clark Reference Brusatte and Clark2015; Young et al. Reference Young, Tennant, Brusatte, Challands, Fraser, Clark and Ross2016a). Previously identified crocodyliforms from Skye include indeterminate remains (Evans et al. Reference Evans, Barrett, Hilton, Butler, Jones, Liang, Parish, Rayfield, Sigogneau-Russell, Underwood, Barrett and Evans2006; Wills et al. Reference Wills, Barrett and Walker2014) and a partial skeleton (Evans & Waldman Reference Evans and Waldman1996) from Cladach a'Ghlinne (Fig. 1B) and, most recently, a partial lower jaw assigned to Theriosuchus sp. (Young et al. Reference Young, Tennant, Brusatte, Challands, Fraser, Clark and Ross2016a) from Valtos (Fig. 1B). The new specimen here represents a hitherto unknown small-bodied taxon from the Middle Jurassic Skye tetrapod assemblage, and has implications for the diversity and ecology of non-pelagic crocodyliforms of the poorly-sampled Middle Jurassic.
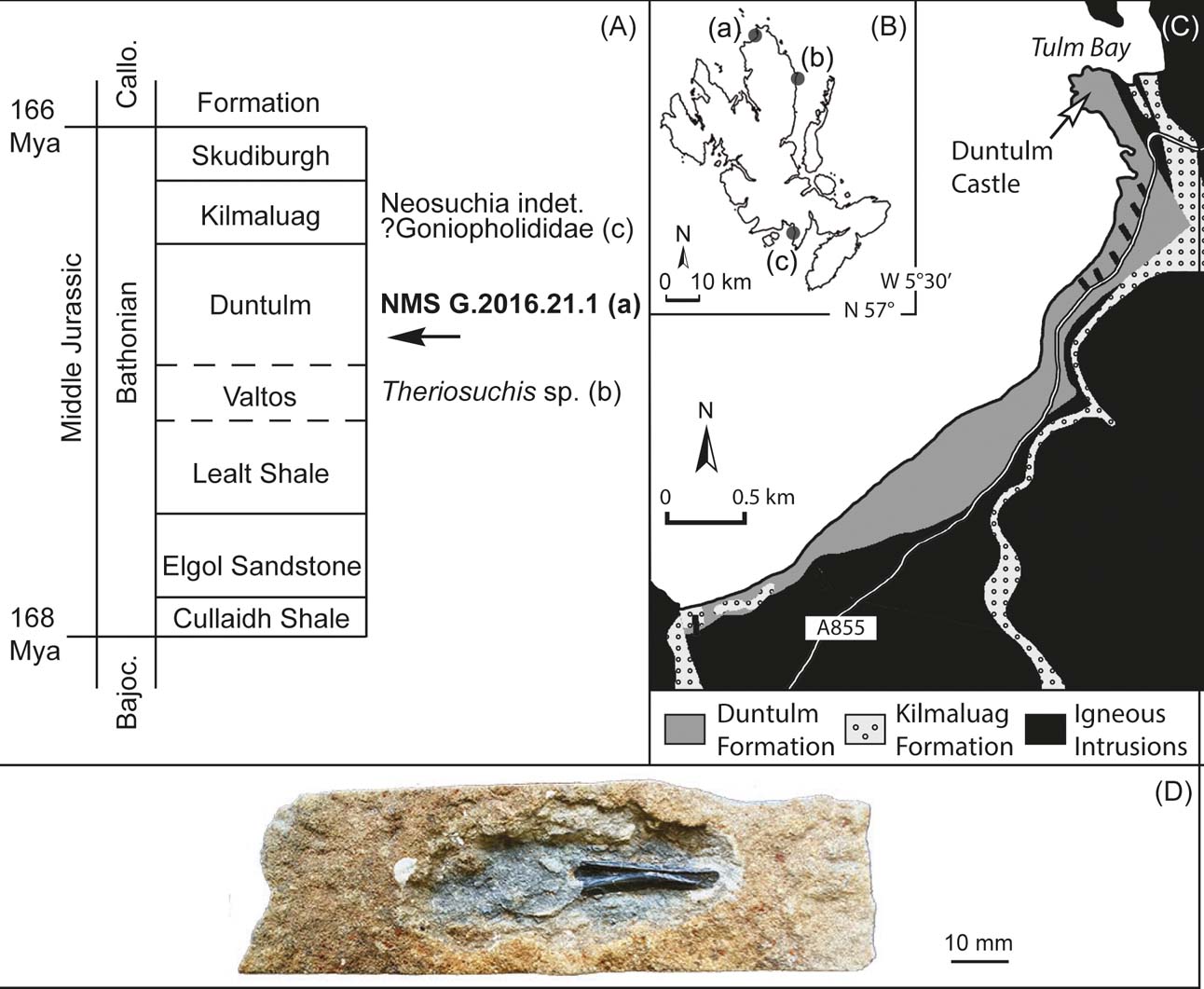
Figure 1 Locality and geological context of NMS G.2016.21.1: (A) stratigraphic occurrences of fossil crocodyliforms discovered from the Great Estuarine Group; (B) map of the Isle of Skye, showing three localities of known fossil crocodyliforms: (a)=Duntulm; (b)=Valtos; (c)=Cladach a'Ghlinne; (C) geological context at the Duntulm locality (a); (D) the new specimen NMS G.2016.21.1.
Note, that herein we use the term Hylaeochampsidae in a broad sense, referring to taxa previously referred to Hylaeochampsidae (compare this clade's taxonomic composition in Buscalioni et al. Reference Buscalioni, Piras, Vullo, Signore and Barbera2011; Puértolas-Pascual et al. Reference Puértolas-Pascual, Canudo and Moreno-Azanza2014; Narváez et al. Reference Narváez, Brochu, Escaso, Pérez-García and Ortega2015; Turner Reference Turner2015; Schwarz et al. Reference Schwarz, Raddatz and Wings2017). We use the term ‘putative hylaeochampsid’ for species with no consensus on their phylogenetic position; i.e., Pachycheilosuchus trinquei Rogers, Reference Rogers2003 and Pietraroiasuchus ormezzanoi Buscalioni et al., Reference Buscalioni, Piras, Vullo, Signore and Barbera2011, as some phylogenetic analyses fail to recover them within Hylaeochampsidae or within a derived neosuchians position (e.g., Narváez et al. Reference Narváez, Brochu, Escaso, Pérez-García and Ortega2015; Tennant et al. Reference Tennant, Mannion and Upchurch2016c). In addition, a recent study using the Turner (Reference Turner2015) matrix recovered the susisuchids Susisuchus and Isisfordia to be paraphyletic, and both to be within Hylaeochampsidae sensu lato (Schwarz et al. Reference Schwarz, Raddatz and Wings2017), highlighting the uncertainty in the taxonomic composition of this clade at present.
Institutional abbreviations. AMNH, American Museum of Natural History, New York, USA; IPFUB, Institut für Paläontologie der Freie Universität Berlin, Berlin, Germany; NHMUK, Natural History Museum, London, UK; NMS, National Museums Scotland, Edinburgh, UK; PC-1, provisional specimen number for Pietraroiasuchus ormezzanoi Buscalioni et al., Reference Buscalioni, Piras, Vullo, Signore and Barbera2011 at Museo Regionale di Scienze Naturali di Torino, Italy; SMU, Shuler Museum of Paleontology, Southern Methodist University, Dallas, Texas, USA.
1. Systematic Palaeontology
Crocodyliformes Benton & Clark, Reference Benton, Clark and Benton1988
Mesoeucrocodylia Whetstone & Whybrow, Reference Whetstone and Whybrow1983
Neosuchia Benton & Clark, Reference Benton, Clark and Benton1988
Neosuchia cf. Hylaeochampsidae
Specimen. NMS G.2016.21.1: an isolated and incomplete, three-dimensionally preserved left lower jaw, including much of the dentary and part of the splenial. The specimen preserves the complete anterior margin of the dentary, but it is broken posteriorly at the 14th alveolus.
Possible unity with hylaeochampsids. Neosuchia cf. Hylaeochampsidae (NMS G.2016.21.1) resembles the putative hylaeochampsids Pachycheilosuchus and Pietraroiasuchus in having shallow dentary alveoli, mildly heterodont dentition and a proportionally short mandibular symphysis extending to the anterior border of the 4th alveolus.
Differential diagnosis. NMS G.2016.21.1 differs from other neosuchians in having an extremely reduced lingual projection along the mandibular symphyseal suture and in having neurovascular foramina present medial to the tooth row, with a one-to-one association with the tooth sockets from alveoli 5–9. NMS G.2016.21.1 differs from Pachycheilosuchus in having a Meckelian canal that is markedly constricted at the 7th alveolus, and it differs from Pietraroiasuchus in having dentary alveoli that are tightly arranged. Furthermore, the new specimen (NMS G.2016.21.1) differs from all other hylaeochampsids in that its 6th dentary alveolus is smaller than the 5th and 7th and slightly raised from the tooth row.
Locality. The specimen was collected from bed 46 of the type section of the Duntulm Formation at the southern part of the exposures [NG407 734] at Cairidh Ghlumaig, Isle of Skye, Scotland, United Kingdom (Morton & Hudson Reference Morton, Hudson and Taylor1995, figs 35–36, table 2; Fig. 1C).
The Duntulm Formation (Bathonian, Middle Jurassic) is part of the Great Estuarine Group, (Anderson Reference Anderson1948), which crops out in portions of the northwest Highlands of Scotland, including the Isles of Skye, Raasay, Eigg and Muck (Harris & Hudson Reference Harris and Hudson1980). It consists of sedimentary rocks of marginal marine and non-marine origin, deposited in the Hebrides Basin during the Bathonian of the Middle Jurassic (Harris & Hudson Reference Harris and Hudson1980).
Seven formations belong to the Great Estuarine Group (Fig. 1A). The upper formations of the Group, following an initial transgression across the fluvial deltas of the Valtos Formation, were deposited during a phase of marine regression (Andrews Reference Andrews1985). It shows a transition from shelly limestone, shale and mudstone in the Duntulm Formation (Andrews & Walton Reference Andrews and Walton1990), to interbedded shale, fine-grained limestone and dolomite (with desiccation cracks) and fine-grained sandstones in the freshwater lagoonal Kilmaluag Formation and, finally, to silty clay, coarse sandstone and intraformational conglomerates in the fluvial Skudiburgh Formation (Andrews Reference Andrews1985).
The Duntulm Formation represents the nearest approach to marine conditions within the Great Estuarine Group. The strata from which NMS G.2016.21.1 was collected have one of the most marine-dominated molluscan faunas within the Duntulm Formation as a whole. In these beds, the small oyster Praeexogyra, which forms nearly monotypic shell beds that dominate much of the formation, is joined by bivalve genera such as Camptonectes, Placunopsis, Corbula and Modiolus, and by echinoid spines, all indicative of shallow marine environment (Andrews & Walton Reference Andrews and Walton1990). The trace fossil Thalassinoides indicates the presence of burrowing crustaceans (Myrow Reference Myrow1995).
With that said, the Duntulm facies are not simply marine. Cyanobacterial (‘algal’) limestones interbedded with the oyster-bearing limestones show evidence of both subaerial exposure and freshwater flushing (Andrews Reference Andrews1986). Carbon, oxygen and strontium isotopic studies of the oysters show that the lagoons where they lived were at times isolated from the sea and suffered evaporation (Holmden & Hudson Reference Holmden and Hudson2003). An intercalation of strata bearing the bivalves Unio and Neomiodon, a few metres above where NMS G.2016.21.1 was discovered, emphasises the potential for freshwater input, confirmed by the isotopic investigations. A scenario depicting the overall environment was presented by Hudson & Trewin (Reference Hudson, Trewin and Trewin2003). Dinosaur fossils, most notably the trackways of large sauropods, are now known from the lower part of the formation (Brusatte et al. Reference Brusatte, Challands, Ross and Wilkinson2016), northeast of where NMS G.2016.21.1 was collected.
Bed 46, from which the crocodyliform specimen came, is sandier than the surrounding beds. Unlike them, it yields the freshwater alga Bothrococcus, probably washed in along with the sand (Andrews & Walton Reference Andrews and Walton1990). In NMS G.2016.21.1, the matrix surrounding the bone is greenish grey sandstone that weathers to a buff-orange colour (Fig. 1D). Besides the crocodyliform jaw, the matrix preserves fish scales, fish teeth and shells of the oyster Praeexogyra. The small oysters are preserved isolated and broken, confirming a high-energy nearshore environment. It is possible that the crocodyliform jaw was washed in; however, given that the dominant fauna of bed 46 is of marine-related genera, we consider NMS G.2016.21.1 most likely to have been a lagoonal species.
Preparation and scanning. Physical preparation exposed the medial surface of the jaw and most of the dorsal surface of the tooth row, whereas the lateral surface and the anterior tip of the dentary remain embedded within the matrix (Fig. 1D). We used high-resolution X-ray computed tomography (HRCT) to visualise the entire specimen, including the embedded portions (Fig. 2). We scanned the fossil in a custom-built machine (constructed and operated by Dr Ian Butler) at the School of GeoSciences, University of Edinburgh, using a 240 kv GE X-ray generator. The scan has a resolution of 44 μm, which was performed under 2.8 w power and with a 0.8 mm Aluminium filter.

Figure 2 CT models and line drawings of specimen NMS G.2016.21.1: (A–B) dorsal view; (C–D) lateral view; (E–F) medial view; (G–H) ventral view. Anatomical abbreviations: D1=first dentary tooth; D6=sixth dentary tooth; d.sp=dentary articulation surface for the splenial; mg=Meckelian groove; n. fr=neurovascular foramina; sp=splenial; sym=mandibular symphysis; v.sp=ventral articulation surface for the splenial. Scale bar=10 mm.
2. Description
The preserved lower jaw shows no obvious taphonomic distortion or post-mortem damage, apart from the break at the posterior end. The specimen is laterally compressed, measuring 28 mm anteroposteriorly, 6 mm dorsoventrally at the deepest point and less than 4 mm labiolingually (Fig. 2). The posterior margin terminates at the 14th dentary alveolus (D14); therefore, we cannot assess the total number of dentary teeth (Fig. 2A–B). The irregular shape of this posterior margin suggests it represents a fracture, rather than the anterior margin of the external mandibular fenestra. This means that the specimen preserves the anterior part of the left dentary and a small fragment of the splenial (Fig. 2E–H). The angular and surangular are not preserved, and we cannot determine whether a mandibular fenestra was present.
2.1. Dentary
In dorsal view, the dental arcade occupies almost the entire mediolateral width of the occlusal surface of the lower jaw. The anterior tooth row has a medial ridge extending between the 2nd and 9th alveoli (D2–D9), which provides an elongate articulation surface for the splenial. We estimate that the splenial extended as far anteriorly as the anterior border of the 6th alveolus (Fig. 2: v.sp). Neurovascular foramina are present medial to the tooth row. Anterior to the 5th alveolus (D5), these foramina are irregularly distributed. From D5 to D9, one foramen is located posterior to each alveolus in the raised tooth row (Fig. 2B).
In lateral view, the dentary has a straight dorsal margin and a concave ventral margin. It gradually tapers dorsoventrally towards the anterior tip and medially into the symphysis (Fig. 2C–D). A series of ten evenly-spaced neurovascular foramina occupy the lateral external surface of the bone. The row of foramina follows the curvature of the ventral margin of the dentary.
Medially, the mandibular symphysis fully occupies the anterior tip of the dentary. The symphysis is anteroposteriorly short in proportion to the dentary, ending at the anterior border of the 4th alveolus (D4). The symphyseal surface bears longitudinal striations for articulation, distinct from deep rugosities in many mesoeucrocodylians (Holliday & Nesbitt Reference Holliday and Nesbitt2013). The Meckelian canal enters the symphyseal plate anteriorly and continues along the entire length of the preserved dentary. It shows a distinct dorsoventral constriction at the 7th alveolus (D7), followed by a trumpet-shaped expansion that opens to the truncated posterior margin of the specimen. A large neurovascular foramen is located in the Meckelian canal posterior to the constricted area, beneath the 11th alveolus (D11). A small fragment of splenial is preserved at the posteroventral margin of the dentary, and the splenial does not appear to have contributed to the symphysis (Fig. 2).
The ventral profile of the dentary is labiolingually narrow. The dentary curves towards the midline of the skull from the 6th alveolus (D6) to the anterior tip (Fig. 2A–B, G–H). We digitally mirrored the specimen to reconstruct a composite full lower jaw for NMS G.2016.21.1. The reconstructed mandibles display a tapering tip at the symphysis (Fig. 3), compared with the more rounded or U-shape outline in most crocodyliforms (Romer Reference Romer1956).

Figure 3 Reconstructed CT models of the anterior mandibles of NMS G.2016.21.1: (A) dorsal view; (B) ventral view. Scale bar=10 mm.
2.2. Alveoli
The dentary preserves 14 alveoli, but no teeth are preserved fully or partially in situ, nor are any associated tooth crowns present within the surrounding matrix. The first five alveoli (D1–D5) are longitudinally elongated and labiolingually compressed, whereas the posterior alveoli (D6–D14) are round. No caniniform tooth is observed in the specimen. Most of the alveoli measure approximately 1.5 mm in diameter, with the 1st, 2nd and 6th alveoli being about half the size of adjacent ones (Fig. 2A–B). Therefore, we infer that the lower jaw dentition was mildly heterodont, with two tooth “waves”. Each alveolus is separated by an interalveolar septum; the first two septa and those posterior to D6 are approximately uniform in width, but those between D3–D5 are almost absent. The septa surrounding the 6th alveolus (D6) are marginally wider, as this alveolus is slightly raised from the tooth row. The preserved alveoli are shallow, extending no deeper than the dorsal margin of the Meckelian canal.
2.3. Developmental stage
With 14 alveoli, the preserved portion of the dentary measures 28 mm (D1–D14). A row of the same number of dentary alveoli measures 30 mm in Knoetschkesuchus (IPFUB Gui Croc 8109: Schwarz & Salisbury Reference Schwarz and Salisbury2005; Schwarz et al. Reference Schwarz, Raddatz and Wings2017), 40–45 mm in Pachycheilosuchus (SMU 75278: Rogers Reference Rogers2003) and Pietraroiasuchus (PC-1: Buscalioni et al. Reference Buscalioni, Piras, Vullo, Signore and Barbera2011), and about 65 mm in Goniopholis (NHMUK PV OR 48300: Salisbury Reference Salisbury2002), which are small-bodied neosuchians, all appearing to be represented by anatomically mature specimens. Therefore, NMS G.2016.21.1 is smaller than an adult, and generally matches the size of a sub-adult or juvenile, when compared with the above taxa. When compared with the extant crocodylian Alligator mississippiensis Daudin, Reference Daudin1802 (a medium-large bodied eusuchian), NMS G.2016.21.1 is equivalent to a hatchling in size (Fig. 4). Future discoveries of fossil material may help better assess the developmental stage of this new Skye specimen.

Figure 4 The mandibular symphysis of Alligator mississippiensis through ontogeny: (A–B) left mandibular rami of a hatchling (AMNH R7129): (A) medial view; (B) dorsal view; (C–D) a medium-sized individual (AMNH R7140): (C) medial view; (D) dorsal view; (E–F) an adult (AMNH R66645): (E) medial view; (F) dorsal view. Scale bars=10 mm.
3. Crocodyliform affinity of NMS G.2016.21.1
The Middle Jurassic of the Isle of Skye yields a diverse array of archosaurs, lepidosaurs, choristoderes, stem mammals and marine reptiles (e.g., Waldman & Savage Reference Waldman and Savage1972; Clark et al. Reference Clark, Boyd, Dixon and Ross1995; Evans & Waldman Reference Evans and Waldman1996; Evans et al. Reference Evans, Barrett, Hilton, Butler, Jones, Liang, Parish, Rayfield, Sigogneau-Russell, Underwood, Barrett and Evans2006; Wills et al. Reference Wills, Barrett and Walker2014; Brusatte et al. Reference Brusatte, Young, Challands, Clark, Fischer, Fraser, Liston, Macfadyen, Ross, Walsh and Wilkinson2015, Reference Brusatte, Challands, Ross and Wilkinson2016; Brusatte & Clark Reference Brusatte and Clark2015; Close et al. Reference Close, Davis, Walsh, Wolniewicz, Friedman and Benson2016; Young et al. Reference Young, Tennant, Brusatte, Challands, Fraser, Clark and Ross2016a). NMS G.2016.21.1 differs from lepidosaurs and choristoderes in having thecodont dentition with rounded alveoli and a single row of neurovascular foramina on the lateral surface (Romer Reference Romer1956; Gao & Fox Reference Gao and Fox1998). It also differs from ichthyosaurs in having shallow tooth sockets: some Triassic ichthyosaurian taxa have thecodont dentition, but their tooth roots attach deeply in the jaw bones (Motani Reference Motani, Callaway and Nicholls1997). The dentary of NMS G.2016.21.1 lacks tooth replacement pits, which contrasts with the multiple rows of dentary teeth in eosauropterygians (Sassoon et al. Reference Sassoon, Foffa and Marek2015). Amongst archosaurs, NMS G.2016.21.1 differs from dinosaurs in having the long Meckelian canal extending into the mandibular symphysis (Romer Reference Romer1956; Holliday & Nesbitt Reference Holliday and Nesbitt2013). A pterosaur affinity is unlikely, as they generally have elongate mandibular symphysis (Romer Reference Romer1956).
Crocodyliforms and some derived “sphenosuchians” have the Meckelian canal extending into the mandibular symphysis (Nesbitt Reference Nesbitt2011), which is the same as in NMS G.2016.21.1. However, “sphenosuchians” have a proportionally longer mandibular symphysis than the new specimen (Colbert & Mook Reference Colbert and Mook1951; Clark & Sues Reference Clark and Sues2002; Clark et al. Reference Clark, Xu, Forster and Wang2004; Pol et al. Reference Pol, Ji, Clark and Chiappe2004). In non-crocodyliform diapsids, the neurovascular foramina are limited to a single line near the dorsal margin of the dentary, whereas in crocodyliforms the neurovascular foramina are more numerous and form a “beehive” arrangement, which is associated with the presence of dome pressure receptors (Soares Reference Soares2002). The arrangement of dentary foramina in NMS G.2016.21.1 is consistent with those of crocodyliforms (Soares Reference Soares2002). Therefore, we exclude NMS G.2016.21.1 from these other groups, and confidently refer it to a small-bodied, juvenile to sub-adult crocodyliform.
4. Taxonomic comparisons within Crocodyliformes
As mentioned previously, there is a diverse range of Middle Jurassic crocodyliform clades either known from fossils or inferred from ghost lineages to which NMS G2016.21.1 could potentially be assigned. Here, we provide extensive comparisons to all clades as a process of elimination to deduce a likely candidate clade for the specimen.
The new specimen (NMS G2016.21.1) has a short mandibular symphysis, shallow dentary alveoli and inconspicuous dentary ornamentation. It differs from large-bodied aquatic clades that evolved elongate symphyseal sutures and lower jaws, such as Thalattosuchia, the most commonly discovered and diverse Middle Jurassic crocodyliforms (e.g., Andrews Reference Andrews1913; Pierce & Benton Reference Pierce and Benton2006; Young et al. Reference Young, Brusatte, Ruta and Andrade2010). Among taxa considered to be “protosuchians”, species with a similarly short mandibular symphyseal suture have markedly different dentary morphology when compared with the new specimen. Zosuchus davidsoni Pol & Norell, Reference Pol and Norell2004a and Gobiosuchus kielanae Osmólska, Reference Osmólska1972 are labiolingually expanded at the anterior tip of the dentary (Osmólska et al. Reference Osmólska, Hua and Buffetaut1997; Pol & Norell Reference Pol and Norell2004a), whereas NMS G.2016.21.1 is labiolingually compressed along the symphyseal suture. Numerous notosuchian genera have anteriorly tapering mandibles with short symphyses, including Araripesuchus (Pol & Apesteguía Reference Pol and Apesteguía2005; Turner Reference Turner2006), Notosuchus (Fiorelli & Calvo Reference Fiorelli and Calvo2008), and Malawisuchus (Gomani Reference Gomani1997). However, the dentary of notosuchians is much more sigmoid than in NMS G.2016.21.1. Therefore, we consider the referral of NMS G.2016.21.1 to Notosuchia to be unlikely.
The new specimen (NMS G.2016.21.1) also differs from several small-bodied “atoposaurids” known from the UK and across Europe. Theriosuchus sp. from the Middle Jurassic of Skye (Tennant et al. Reference Tennant, Mannion and Upchurch2016c; Young et al. Reference Young, Tennant, Brusatte, Challands, Fraser, Clark and Ross2016a) has deeper rugosities in the symphyseal plate, more prominent ornamentation on the lateral surface of the dentary and a different alveolar morphology. Compared with NMS G.2016.21.1, Alligatorium has a curved dorsal margin of the dentary, and Atoposaurus and Alligatorellus have the splenial contributing to the mandibular symphysis; although the latter two most likely represent skeletally immature specimens (Tennant & Mannion Reference Tennant and Mannion2014; Tennant et al. Reference Tennant, Mannion and Upchurch2016c). Montsecosuchus, a small-bodied, phylogenetically uncertain neosuchian from Spain (Buscalioni & Sanz Reference Buscalioni and Sanz1990; Tennant et al. Reference Tennant, Mannion and Upchurch2016c), has more strongly developed dentary ornamentation than NMS G.2016.21.1.
The Skye jaw (NMS G.2016.21.1) differs from genera consistently found to be hylaeochampsids. Recent analyses that gave conflicting evolutionary positions for the putative hylaeochampsid Pachycheilosuchus do not do so for Acynodon or Iharkutosuchus, instead consistently recovering them within Hylaeochampsidae (e.g., Turner Reference Turner2015; Narváez et al. Reference Narváez, Brochu, Escaso, Pérez-García and Ortega2015; Schwarz et al. Reference Schwarz, Raddatz and Wings2017). Acynodon, although originally described as being more closely related to crocodylians, is known from the Late Cretaceous of Europe, and includes preserved dentary material for multiple specimens (Delfino et al. Reference Delfino, Martin and Buffetaut2008a; Martin Reference Martin2007). However, Acynodon differs from NMS G.2016.21.1 in the vast stratigraphic range disparity, as well as in multiple dentary characteristics: the symphysis extends more than four alveoli, the dentary alveoli form a chain and are small and laterally placed relative to the dorsal surface and the ventral surface is greatly mediolaterally expanded and flattened (Delfino et al. Reference Delfino, Martin and Buffetaut2008a; Martin Reference Martin2007). The dentary of Iharkutosuchus is distinct from NMS G.2016.21.1 in that the symphysis extends posteriorly to the D8 alveolus, and in that the dental arcade forms an arcuate shape, shifts posteriorly from a lateral position to a more medial position on the occlusal surface, and has a mediolaterally expanded anterior margin similar to other eusuchians (Ősi Reference Ősi2008). Unfortunately, the type species Hylaeochampsa vectiana Owen, 1874 does not preserve the dentary (Clark & Norell Reference Clark and Norell1992). Similarly, comparison with Allodaposuchus, a possible Late Cretaceous allodaposuchid or putative hylaeochampsid, depending on analysis used (see Buscalioni et al. Reference Buscalioni, Piras, Vullo, Signore and Barbera2011; Delfino et al. Reference Delfino, Codrea, Folie, Dica, Godefroit and Smith2008b; Puértolas-Pascual et al. Reference Puértolas-Pascual, Canudo and Moreno-Azanza2014; Narváez et al. Reference Narváez, Brochu, Escaso, Pérez-García and Ortega2015), is not possible, as no known specimen preserves the dentary. A recent study extending the Turner (Reference Turner2015) analysis found the susisuchids Susisuchus and Isisfordia to be paraphyletic within an unnamed clade comprised of all potential hylaeochampsids (Schwarz et al. Reference Schwarz, Raddatz and Wings2017). The symphyseal region of the mandible of Susisuchus is labiolingually expanded, which differs from NMS G.2016.21.1 (Salisbury et al. Reference Salisbury, Frey, Martill and Buchy2003). The dentary of Isisfordia cannot be compared with NMS G.2016.21.1, due to the holotype lacking the anterior region (Salisbury et al. Reference Salisbury, Molnar, Frey and Willis2006; Fortier & Schultz Reference Fortier and Schultz2009).
Curiously, NMS G.2016.21.1 is most similar to two species of putative hylaeochampsids (Turner Reference Turner2015): Pachycheilosuchus trinquei from the Early Cretaceous of Texas, USA (Rogers Reference Rogers2003) and Pietraroiasuchus ormezzanoi from the Early Cretaceous of Italy (specimen PC-1 in Buscalioni et al. Reference Buscalioni, Piras, Vullo, Signore and Barbera2011). The Skye jaw (NMS G.2016.21.1) shares with Pachycheilosuchus and Pietraroiasuchus a short mandibular symphysis, extending to the anterior border of the 4th alveolus (Figs 5, 6C–F). All three of the taxa have mild heterodont dentition and lack caniniform tooth (inferred for NMS G.2016.21.1 by the absence of any greatly enlarged alveoli). The Skye jaw is similar to Pietraroiasuchus in the gently arcuate form of the arcade in dorsal view, and in that the splenial does not contribute to the symphyseal area (Buscalioni et al. Reference Buscalioni, Piras, Vullo, Signore and Barbera2011). In particular, the Skye jaw shares with Pachycheilosuchus a row of evenly spaced and ventrally positioned neurovascular foramina on the lateral surface of the dentary, which is a unique feature among hylaeochampsids (Fig. 6A–B). However, NMS G.2016.21.1 cannot be assigned to either of the two above genera. It is different from Pietraroiasuchus in the medially expanded anterior occlusal shelf in the latter (Buscalioni et al. Reference Buscalioni, Piras, Vullo, Signore and Barbera2011). In comparison with NMS G.2016.21.1, most small-bodied neosuchians display varying degrees of labiolingual expansion at the anterior tip of the dentary (Young et al. Reference Young, Steel, Foffa, Price, Naish and Tennant2014, fig. 8; Young et al. Reference Young, Tennant, Brusatte, Challands, Fraser, Clark and Ross2016a, fig. 3; Fig. 5C). The dentary of Pachycheilosuchus has a small lingual projection along the symphyseal suture (Fig. 6D), but the anterior tip of the dentary is less labiolingually compressed than that in the Skye jaw (Fig. 5A–B).
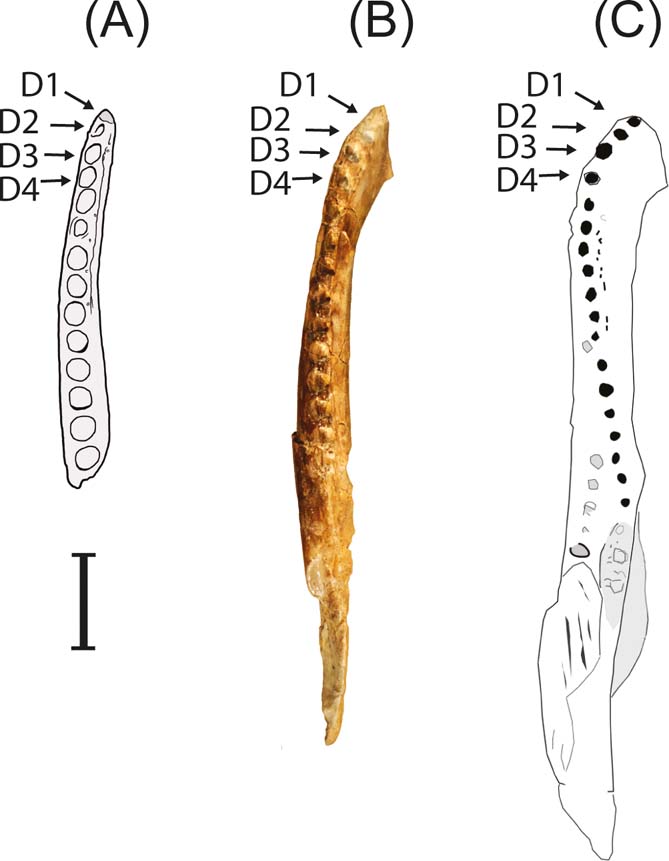
Figure 5 Lower jaw comparisons between NMS G.2016.21.1 and Early Cretaceous hylaeochampsids: (A) the Duntulm specimen (NMS G.2016.21.1); (B) Pachycheilosuchus trinquei (SMU75279). Photograph by D. Winkler; (C) Pietraroiasuchus ormezzanoi (PC-1, modified from Buscalioni et al. Reference Buscalioni, Piras, Vullo, Signore and Barbera2011). D1–D4 denotes first four alveoli.

Figure 6 Morphological comparisons between NMS G.2016.21.1 (A, C, E) and Pachycheilosuchus trinquei (SMU75279: B, D, F): (A–B) dentary in lateral view. A row of neurovascular foramina evenly distributes along the lateroventral margin in both taxa; (C–D) dentary in dorsal view. D1–D4 denotes first four alveoli; (E–F) the Meckelian canal expands dorsoventrally in the new specimen, but constricts anteriorly in P. trinquei. Dashed lines indicate the posterior limit of the symphyseal plate. Not to scale.
4.1. Impact of ontogeny on assessment of NMS G.2016.21.1
The uncertainty of the ontogenetic stage of NMS G.2016.21.1 makes assessing its phylogenetic position problematic. Characters that unite NMS G.2016.21.1 with Pachycheilosuchus, the lack of dentary ornamentation and the shallow articulation rugosities in the symphyseal plate, are known to vary during ontogeny. In extant crocodylians, young individuals have shallow dermatocranial ornamentations that develop into prominent pits and rugosities in adults (Mook Reference Mook1921; Joffe Reference Joffe1967). But lack of ornamentation may also imply a phylogenetic signal. Several fossil crocodyliforms represented by mature specimens have a smooth or weakly pitted external surface of the dentary. These include several “protosuchian” species (Yang Reference Yang1973; Wu et al. Reference Wu, Sues and Dong1997; Pol & Norell Reference Pol and Norell2004a, Reference Pol and Norellb), various metriorhynchid genera (e.g., Young et al. Reference Young, Andrade, Etches and Beatty2013), the atoposaurid Alligatorium (Tennant & Mannion Reference Tennant and Mannion2014), and Pachycheilosuchus (Rogers Reference Rogers2003).
Similarly, shallow rugosities in the symphyseal plate of NMS G.2016.21.1 may be phylogenetically informative, but these rugosities become deeper through ontogeny in extant crocodylians (Fig. 4). The dominant articulation type in Mesoeucrocodylia is Class III symphysis with interdigitated rugosities (Holliday & Nesbitt Reference Holliday and Nesbitt2013), which is lacking in Pachycheilosuchus and NMS G.2016.21.1 (Fig. 6F). The latter two taxa have Class II articulation with shallow rugosities. The uncertain developmental stage of NMS G.2016.21.1 considered, these observations add information to “typical” symphyseal morphology in Neosuchia and potentially more inclusive clades in Crocodyliformes.
The lingual projection of the symphyseal suture remains reasonably stable throughout the ontogeny of extant crocodylians, which we observed in the lower jaw of Alligator mississippiensis (Fig. 4). It retains a labiolingual expansion near the anterior tip of the symphyseal suture, from hatchling-sized juveniles (AMNH R7129; Fig. 4A–B) to medium-sized young individuals (AMNH R7140; Fig. 4C–D), and into skeletal maturity (AMNH R66645; Fig. 4E–F). If Alligator is a good model for the ontogeny of the taxon NMS G.2016.21.1 belongs to, this indicates that the mediolateral constriction of the symphyseal suture observed in NMS G.2016.21.1 may not be an ephemeral juvenile feature, but may also be present in adult individuals; and, therefore, could be potentially diagnostic of a new taxon.
Based on the above comparisons, NMS G.2016.21.1 shares multiple mandibular characteristics with Pachycheilosuchus and Pietraroiasuchus, but its unique symphyseal region differs from all other neosuchians compared herein. We do not name a new taxon, given the incomplete nature of the specimen and its likely juvenile status. Additional skull material and further understanding of ontogenetic changes in crocodyliform mandibles will help assess the phylogenetic position of NMS G.2016.21.1.
5. Discussion
Neosuchia cf. Hylaeochampsidae (NMS G.2016.21.1) is the first known definitive crocodyliform from the Duntulm Formation, which extends our knowledge of the Middle Jurassic archosaur assemblage of the Isle of Skye (Clark et al. Reference Clark, Boyd, Dixon and Ross1995; Evans et al. Reference Evans, Barrett, Hilton, Butler, Jones, Liang, Parish, Rayfield, Sigogneau-Russell, Underwood, Barrett and Evans2006; Evans & Waldman Reference Evans and Waldman1996; Wills et al. Reference Wills, Barrett and Walker2014; Brusatte & Clark Reference Brusatte and Clark2015; Brusatte et al. Reference Brusatte, Challands, Ross and Wilkinson2016; Young et al. Reference Young, Tennant, Brusatte, Challands, Fraser, Clark and Ross2016a). This specimen differs from other crocodyliforms currently known from Skye, including Theriosuchus sp. (Young et al. Reference Young, Tennant, Brusatte, Challands, Fraser, Clark and Ross2016a) from the Valtos Formation (Fig. 1A). Theriosuchus sp. has deep dentary ornamentation, strongly heterodont alveoli and a unique “starburst” morphology in the mandibular symphysis (NMS G.2014.52.1: Young et al. Reference Young, Tennant, Brusatte, Challands, Fraser, Clark and Ross2016a; Fig. 2D), none of which are present in NMS G.2016.21.1. Isolated neosuchian remains are also known from the Kilmaluag Formation that is stratigraphically immediately above the Duntulm Formation (Evans & Waldman Reference Evans and Waldman1996; Evans et al. Reference Evans, Barrett, Hilton, Butler, Jones, Liang, Parish, Rayfield, Sigogneau-Russell, Underwood, Barrett and Evans2006; Wills et al. Reference Wills, Barrett and Walker2014). The Kilmaluag material – comprising teeth, osteoderms and incomplete postcranial skeletons – is not directly comparable with NMS G.2016.21.1 due to lack of overlapping skeletal elements. It is possible that these specimens belong to the same taxon as NMS G.2016.21.1 and, if so, one or more of them would make more suitable holotypes, which is one reason we do not name NMS G.2016.21.1 here. These Kilmaluag specimens are currently under study, so their systematics and relationships with NMS G.2016.21.1 will become clearer in the future. These discoveries show that there were various small-sized neosuchians in the nearshore ecosystems of the Isle of Skye during the Middle Jurassic.
5.1. Implications of NMS G.2016.21.1 as a potential hylaeochampsid
The morphological resemblance of NMS G.2016.21.1 to Pachycheilosuchus and Pietraroiasuchus could potentially have implications for the evolution of Hylaeochampsidae that currently is only known from the Cretaceous (Norell & Clark Reference Norell and Clark1990; Pol et al. Reference Pol, Turner and Norell2009; Turner Reference Turner2015). Should Pachycheilosuchus and Pietraroiasuchus be referred to Hylaeochampsidae, then NMS G.2016.21.1 would suggest this clade has a much longer evolutionary history than previously known. The type species Hylaeochampsa vectiana is known from the Early Cretaceous of the Isle of Wight (southern coast of the United Kingdom), whilst all other species consistently referred to Hylaeochampsidae are known from the Late Cretaceous of Europe (Clark & Norell Reference Clark and Norell1992; Martin Reference Martin2007; Delfino et al. Reference Delfino, Martin and Buffetaut2008a, Reference Delfino, Codrea, Folie, Dica, Godefroit and Smithb; Puértolas-Pascual et al. Reference Puértolas-Pascual, Canudo and Moreno-Azanza2014). Therefore, NMS G.2016.21.1 potentially could extend the temporal range of Hylaeochampsidae, or a slightly more inclusive clade, into the Middle Jurassic, and geographically into the north of the United Kingdom.
However, it is possible that NMS G.2016.21.1, Pachycheilosuchus and Pietraroiasuchus form a clade of derived neosuchians that are basal to Eusuchia. The variable phylogenetic position of Pachycheilosuchus is one of the biggest issues in determining where NMS G.2016.21.1 belongs on the crocodyliform tree, second only to the specimen's general incompleteness. Pachycheilosuchus was originally described as a possible atoposaurid (Rogers Reference Rogers2003), but Turner (Reference Turner2015) recovered it at the base of Eusuchia. Other studies have recovered this species in radically different positions, such as: in a more basal position within Neosuchia, closer to the split with Tethysuchia (Adams Reference Adams2014); as a derived neosuchian but basal to Eusuchia (Narváez et al. Reference Narváez, Brochu, Escaso, Pérez-García and Ortega2015); as a basal neosuchian, even more basal than the Tethysuchia–Eusuchia split (Tennant et al. Reference Tennant, Mannion and Upchurch2016b); or in a clade with an assortment of hylaeochampsids and susisuchids (Schwarz et al. Reference Schwarz, Raddatz and Wings2017). The phylogeny of Turner (Reference Turner2015) is consistent with Buscalioni et al. (Reference Buscalioni, Piras, Vullo, Signore and Barbera2011) in the monophyly of Hylaeochampsidae, despite the fact that the latter study found Pietraroiasuchus as the sister group of Pachycheilosuchus (although they did not sample Acynodon). Young et al. (Reference Young, Hastings, Allain and Smith2016b) recovered Pachycheilosuchus and Pietraroiasuchus as sister taxa, and that clade was the sister taxon to Crocodylia. However, Young et al. (Reference Young, Hastings, Allain and Smith2016b) did not sample any other hylaeochampsids.
The incompleteness of NMS G.2016.21.1 hinders its inclusion into a rigorous phylogenetic analysis of Hylaeochampsidae, but new dentary characters are observed in multiple putative members of the clade and will be important in future systematic evaluations. The mandibular symphysis extends less than four alveoli in NMS G.2016.21.1, Pachycheilosuchus (Rogers Reference Rogers2003) and Pietraroiasuchus (Buscalioni et al. Reference Buscalioni, Piras, Vullo, Signore and Barbera2011), and it is shorter than five alveoli in Acynodon iberoccitanus (Martin Reference Martin2007). In all these taxa, the dentary alveoli extend no deeper than the dorsal margin of the Meckelian canal. These characters could potentially be diagnostic of Hylaeochampsidae.
Several clades of small-to-medium-sized Mesozoic neosuchians have been suggested to be closely positioned to the origin of Eusuchia, including Hylaeochampsidae, Susisuchidae, Bernissartiidae and Paralligatoridae, although the taxonomic compositions of these clades vary among crocodyliform phylogenies (e.g., Turner & Sertich Reference Turner and Sertich2010; Adams Reference Adams2014; Turner Reference Turner2015; Young et al. Reference Young, Hastings, Allain and Smith2016b; Schwarz et al. Reference Schwarz, Raddatz and Wings2017). However, the various phylogenetic analyses appear to converge on a divergence between the large-bodied and primarily aquatic neosuchian clades (Pholidosauridae+Dyrosauridae) and the relatively smaller-bodied, terrestrial or semi-aquatic clades (although the position of Goniopholididae is variable; Reference AdamsAdams 2014; Turner Reference Turner2015; Young et al. Reference Young, Hastings, Allain and Smith2016b; Schwarz et al. Reference Schwarz, Raddatz and Wings2017). Yet little is known about neosuchians before the Late Jurassic due to a patchy non-marine fossil record (Mannion et al. Reference Mannion, Benson, Carrano, Tennant, Judd and Butler2015; Tennant et al. Reference Tennant, Mannion and Upchurch2016b). The earliest definitive record of pholidosaurid material comes from the Late Jurassic of Uruguay and France (Fortier et al. Reference Fortier, Perea and Schultz2011). The Bathonian age for the putative pholidosaurid Anglosuchus was questioned by Watson (Reference Watson1911), and the specimens referred to this taxon cannot be located.
Discovery of a specimen morphologically similar to putative hylaeochampsids from the Middle Jurassic suggests that small body size, which is widespread among basal eusuchians, emerged early in Neosuchia. We remain cautious in this conclusion, however, because an accurate estimate of adult body size of NMS G.2016.21.1 cannot be determined due to its incomplete nature and likely juvenile status. Further investigation is required to elucidate the distribution, mode and magnitude of body size evolution among advanced neosuchians, and we suggest that the discovery of new Middle Jurassic fossils from places like the Isle of Skye may hold the key.
6. Conclusion
The new lower jaw from the Isle of Skye helps shed light on the evolution of crocodyliforms during the mysterious Middle Jurassic interval. Whilst it has several distinctive dentary characters, particularly in relation to the Meckelian canal and mandibular symphysis, we do not refer it to a new taxon due to its incompleteness. Based on similarities in the mandibular symphyseal suture and dentary alveoli, this specimen appears to have an affinity with the Early Cretaceous putative hylaeochampsids Pachycheilosuchus trinquei and Pietraroiasuchus ormezzanoi. Furthermore, the anterior dentary and alveolar morphological variation described herein will hopefully contribute to future phylogenetic analyses, especially those including taxa referred to Hylaeochampsidae. Along with other discoveries from Skye, this new specimen shows that small-bodied neosuchians were diversifying in the Middle Jurassic, during a time when their fossil record is extremely poor.
7. Acknowledgements
Ian Butler (University of Edinburgh) performed the CT scanning in November 2015. For access to modern crocodilian specimens, we thank David Kizirian, David Dicky, Margret Arnold and Lauren Vonnahme at the American Museum of Natural History. We thank Alan Turner and another reviewer for insightful comments that improved the quality of the manuscript. HY was funded by the Royal Society Sino-British Incoming Postdoctoral Fellowship and the University of Edinburgh. JPT is funded by the National Environmental Research Council. Fieldwork by the University of Edinburgh and the PalAlba Group on the Isle of Skye was funded by Richard Deveira, the Edinburgh Zoo and Royal Zoological Society of Scotland, the Edinburgh Geological Society, the Systematics Research Fund (administered by the Linnean Society of London and the Systematics Association) and the University of Edinburgh. SLB's lab is funded by a Marie Curie Career Integration Grant (CIG 630652) and a Royal Society Research Grant (RG130018).







 Open access
Open access