


Introduction
When the early vertebrate Euphanerops longaevus from the Upper Devonian of Canada was first described it was compared to and later referred to the Anaspida (Woodward, Reference Woodward1900, Reference Woodward and von Zittel1902), a group recognised by Traquair (Reference Traquair1899) on the basis of Silurian material from Scotland. Remarkably, the beds that yielded these anaspids also contain a fairly common euphaneropid that has not been recorded previously. The elusive nature of this animal is attributed to the often poorly visible remains of soft tissue comprising the body and it closely recalls recent discoveries of euphaneropids from Middle Devonian deposits at Achanarras Quarry in Caithness, Scotland (Newman & Trewin, Reference Newman and Trewin2001; Newman, Reference Newman2002). The aim of the present article is to describe and name a new Silurian vertebrate that belongs to the family Euphaneropidae Woodward, Reference Woodward1900 (see Table 1 for the known representatives), and to shed some light on the anatomy of this enigmatic group.
Table 1. An overview of representatives of the family Euphaneropidae known to date.

a Endeiolepis is preserved in different types of sediment from Euphanerops, but is regarded as a possible junior synonym (see Janvier & Arsenault, Reference Woodward2007).
Methods and material
The present specimens originate from several outcrops of the type attributed to the Lower Silurian Fish Bed Formation and equivalent horizons (for stratigraphical details, see Märss & Ritchie, Reference Märss and Ritchie1998), as based on the faunal associations from this unit which include the anaspids Lasanius problematicus and Birkenia elegans, the thelodonts Lanarkia spp. and Shielia taiti, the osteostracan Ateleaspis tessellata, and the mixopterid Lanarkopterus dolichoschelus. In order to understand better the soft tissues of the new taxon, a detailed comparison is made with Euphanerops longaevus as described by Janvier & Arsenault (Reference Janvier and Arsenault2007). The smaller individuals of this species appear to represent a similar form. In view of the fact that the fossilised remains of this new form are almost entirely composed of soft tissue (which normally are poorly preserved at best), it turned out to be necessary to collect hundreds of specimens in order to demonstrate the more delicate and rarely preserved features.
Systematic palaeontology
Phylum Chordata Haekel, 1874
Subdivision Craniata Lankester, 1877
Family Euphaneropidae Woodward, 1900
Genus Ciderius nov.
Derivation of name: With reference to cider, because the best-preserved features resemble a bottle, a barrel and a pair of apples when flattened.
Type species: Ciderius cooperi sp. nov.
Diagnosis: As for type species (monotypy).
Ciderius cooperi sp. nov.
Derivation of name: In honour of David John Cooper, acknowledging his co-operation during excavations.
Holotype: Oertijdmuseum De Groene Poort, Boxtel (the Netherlands) collections, no. MAB 6000, a complete individual with a long, whip-like tail.
Paratypes: MAB 6001, a near-complete specimen with clear head stains and a large digestive organ; MAB 6002, a slab with an individual with well-preserved eyes; MAB 6003, near-complete individual showing clear mineralised parachordals; MAB 6004, two partial individuals, arranged in parallel, the larger of which shows a detailed section of the branchial apparatus.
Horizon and locality: Fish Bed Formation (and equivalent horizons), lower Wenlock (Lower Silurian), Midland Valley of Scotland.
Diagnosis: Long and slender body comparable in shape to other euphaneropids and ranging in length from 30 to 150 mm; cranial region displaying a median barrel-shaped object, paired eyes and mineralised parachordals; greatly elongated branchial apparatus present between approximately 15% and 65% of entire length of body; bottle-shaped digestive apparatus situated in region approaching anterior end of branchial apparatus, possibly between branchial apparatus itself; vertical rows of short longitudinal stripes on sagittal plane of posterior half of body; fins apparently absent.
Preservation
With the exception of a pair of mineralisations, these fossils comprise remains of soft tissues of variable preservation, not only at different localities, but also between and on bedding planes. When deposited on the lake or lagoon floor, most specimens were articulated, but deformation also occurs, accounting for dart-like and bloated specimens. Nearly all specimens have a body that was arched to some degree, which may be the result of the dorso-ventrally flattened anterior and laterally flattened posterior body shape that pushed the carcass into such a position or, possibly, rigor mortis or some other post-mortem process could be responsible. The most frequently preserved parts of the anatomy are the three head stains and the bottle-shaped digestive apparatus. These are visible as black films, the latter often containing ingested sediment. Occasionally, these are the sole remains discerned; it is possible that such fossils represent the final, most strongly decayed state of the body. Objects here referred to as ‘black lines’ are rarely distinguishable as such because they either have disintegrated and are now visible only as patchy black impressions covering a broad area of their original position, or are absent altogether. Similarly, the ‘longitudinal stripes’ infrequently display much detail, but despite this are visible in about 50% of the material in the form of faint vertical bands. The paired mineralisations of the head appear to have been particularly resistant to decay, being visible in almost all articulated individuals.
Morphology
The absence of any body outline or a trace of fins leaves many questions on body morphology unanswered. However, it should be noted that some specimens seemingly possess skin impressions, although it is more likely that these are in fact the disintegrated remains of internal organs. The black lines in particular seem to be responsible for this image. This is evident anteriorly where there are no black lines and the head stains appear separated from the rest of the body (e.g. Fig. 1). The best idea of body shape is based on the sole specimen that is preserved head to tail (the holotype, MAB 6000; Fig. 1), as well as on Euphanerops. Janvier & Arsenault (Reference Janvier and Arsenault2007) showed Euphanerops with a fusiform body, probably slightly deeper than broad, the head having an overhanging snout, an oral region possibly containing an annular cartilage, an elongated gill apparatus located on the ventral to ventro-lateral flanks, an anal fin (although paired anal fins are also possible; see Sansom et al., Reference Sansom, Gabbott and Purnell2013a), and a relatively large epicercal lobe. Whether all of these features were already present in the stratigraphically older Ciderius gen. nov. is not clear. However, at least one point of divergence in comparison with Janvier & Arsenault's (2007) reconstruction of Euphanerops seems worth noting. It is possible that the branchial apparatus was positioned laterally to ventro-laterally, thus giving the anterior half of the body a more dorso-ventrally flattened appearance than seen in Euphanerops. This is indicated by the propensity for dorso-ventral preservation of this region. There are also quite a few specimens (e.g. Figs 3 and 4) that reveal remains of the branchial apparatus superimposed on the bottle-shaped digestive apparatus, which is unlike the position in Euphanerops in which the former has been suggested to be ventral to the latter.
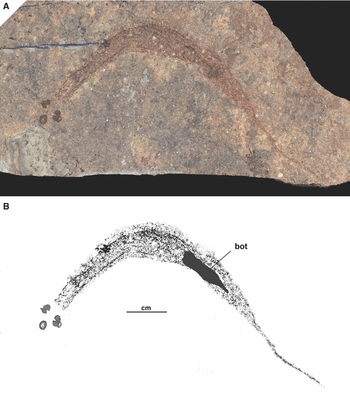
Fig. 1. Holotype (MAB 6000) of Ciderius cooperi gen. nov., sp. nov. A. Complete specimen composed of largely disarticulated remains of the internal anatomy; B. Drawing of A showing the position of the bottle-shaped object (bot). Lower Wenlock, Fish Bed Formation and equivalent horizons), Midland Valley of Scotland.
Barrel-shaped object
Description: The anterior head stain usually is preserved as an amorphous dark spot that is slightly larger than the adjacent paired head stains. A small number of specimens reveal it to be a three-dimensional structure that appears to consist of three distinct parts forming a barrel-shaped object. Anteriorly a small cylinder occurs, behind which a plate seems to be present that covers the posterior side (Fig. 2A and B). The anterior opening of the cylinder is often surrounded by what appears to be a dark rim (Fig. 2A–D). Owing to the dorso-ventral flattening of this object, the natural orientation, and in particular the direction of the opening, remains unclear. Attempts to discover this by examining laterally preserved specimens of other euphaneropids have not been conclusive.
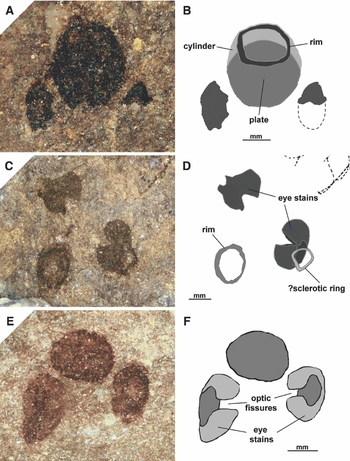
Fig. 2. Head region of several specimens of Ciderius cooperi gen. nov., sp. nov., showing the characteristic three head stains. A. Head region of paratype, MAB 6001; B. Drawing of A, showing the barrel-shaped object consisting of an anterior opening marked by a darkened rim, a cylinder and a plate. The plate is largely obscured by the cylinder element; C. Enlargement of the head region of the holotype, MAB 6000; D. Drawing of C, showing the position of the possible sclerotic ring in relation to the eye stains as well as the rim surrounding the anterior opening of the barrel-shaped object; E. Head stains of paratype, MAB 6002; F. Drawing of E, showing the lateral eye stains with optic fissures. Lower Wenlock, Fish Bed Formation (and equivalent horizons), Midland Valley of Scotland.
Interpretation: Both in relative position and size, this object corresponds to the anterior head stains of the genera Euphanerops and Achanarella. It may also be the same object as the anterior stain of Jamoytius reported by Ritchie (Reference Ritchie1968). Because previous authors did not recognise the barrel shape of this object, their interpretations should be looked into first. Existing ideas include a tectal cartilage plate that armed the snout, as in lampreys (e.g. in Euphanerops; Janvier & Arsenault, Reference Janvier and Arsenault2007), a mouth opening or annular cartilage, as in Jamoytius (Ritchie, Reference Ritchie1968; Sansom et al., Reference Sansom, Freedman, Gabbott, Aldridge and Purnell2010) and in Achanarella (Newman, Reference Newman2002), nasal opening (in Jamoytius; Sansom et al., Reference Sansom, Freedman, Gabbott, Aldridge and Purnell2010), and a median eye (in Euphanerops; see Loosemore, Reference Loosemore2009). While the position of this object corresponds to either of the tectal cartilage plates in lampreys, the shape does not. It appears to be three dimensional rather than a plate-shaped element such as the anterior and posterior tectal cartilage plates of lampreys.
An interpretation as an annular cartilage is not supported by the barrel shape. However, the anterior opening could be an oral aperture, of unknown configuration, because it has not been convincingly accounted for elsewhere (see Janvier & Arsenault, Reference Janvier and Arsenault2007). The remainder of the object could then be a small oral cavity. However, two issues conflict with such an idea. An indication that this object allowed for the passage of food particles has not been found; the plate component appears to block the posterior end (except, possibly, for Jamoytius, pers. obs.). Secondly, if food particles could travel through this object then it may be on the small side for the oral opening of an animal that was probably a filter-feeder, given the presence of such a large branchial apparatus.
Another possibility is that it represents a part of the olfactory organ. This could then be the nasal sac or some type of tissue that lined the wall of the nasal organ and that left such a black impression. The position, anterior to the eyes, is conventional for a vertebrate olfactory organ while the anteriorly directed opening and barrel shape appear suitable for a nasal sac. While this is the most likely interpretation at this point, it is difficult to confirm as there is no example, fossil or extant, to compare it with. Its shape is unlike the nasal sac of lampreys (capsula nasalis; Fig. 22, Marinelli & Strenger, Reference Marinelli and Strenger1954), or the nasal tube of hagfishes (tuba nasalis; Fig. 84, Marinelli & Strenger, Reference Marinelli and Strenger1956), two extant forms generally regarded as the closest living proxies. None of the fossil lampreys or hagfish species have been reported with preserved nasal sacs either. This appears to be echoed by Sansom et al.'s (2013b) decay experiments on extant hagfishes and lampreys, which do not indicate the nasal sacs to be particularly resistant to decay. It can also be said that anaspids, the group usually mentioned in one breath with euphaneropids, also do not show a trace of nasal sacs. This leaves the Cambrian forms Haikouichthys and Metaspriggina as the only naked early vertebrates that could contain nasal sacs. They have been suggested to possess closely grouped paired nasal sacs that lie anterior or between the eye stains (see Shu et al., Reference Shu, Conway Morris, Han, Zhang, Yasui, Janvier, Chen, Zhang, Liu, Li and Liu2003; Conway Morris & Caron, Reference Conway Morris and Caron2014). However, unlike the barrel-shaped object, these are paired and relatively much smaller.
The last option to be discussed is perhaps the most unusual. The new data provided on the other (paired) head stains indicate that they are the remains of the eye capsules. This makes it unlikely that the barrel-shaped object is a median eye of the cyclopean origin suggested by Loosemore (Reference Loosemore2009), which notion relied on the absence of paired eyes. However, there still remains the possibility that this is a photoreceptive organ of some kind, and two lines of thought could support this interpretation. The three head stains are similarly preserved in that they appear to be equally decay resistant and occur equally clearly even when the rest of the animal, except for the ‘bottle-shaped object’, is no longer visible. This might support the view that all three head stains constitute the remains of the same type of tissue. Since the other two head stains probably represent the remains of retinal pigments, the barrel-shaped object could also be composed of tissue associated with a photoreceptive organ. This could then be the pigment of a large-parietal or pineal eye. Furthermore, the barrel shape of this object may be suitable to have contained a light-gathering pit of a photoreceptive organ, such as a parietal or pineal eye. The problem with this interpretation, however, is that it would be rather unconventional. The relative size of this object, i.e. larger than the paired eyes, and its rostral position are quite unlike the pineal and parietal eyes of primitive fishes, which are commonly used for comparative purposes of presumed agnathan fossils (e.g. lampreys and hagfishes). Like the nasal sac, a parietal or pineal eye has not been recorded in Sansom et al.'s (2013b) decay experiments as particularly decay resistant.
In summary, none of the interpretations above are supported by conclusive evidence, although the nasal sac interpretation seems most probable at this point. Better-preserved specimens are needed to improve our understanding of the morphology and orientation of this structure.
Paired eyes
Description: Postero-lateral to the barrel-shaped object are a pair of stains which appear apple-shaped when flattened (see Fig. 2C–F). They seem to be impressions of rounded objects with laterally directed external openings, giving each object the appearance of a cup. This conforms to the description by Sansom et al. (Reference Sansom, Freedman, Gabbott, Aldridge and Purnell2010) of the paired head stains in Jamoytius. However, there is a small difference. There is a cleft present, in the same location on each stain (see Fig. 2E and F). This cleft occurs across the dorsal part of the cup, which would give it a lateral appearance slightly resembling a horseshoe. Its widespread occurrence in the newly collected material indicates that it is more than a mere crack or rip as a result of decay.
Interpretation: Paired anterior stains such as these have been recorded from numerous other vertebrates. In fact, they have long been considered eye remains, or to have been associated with such organs (e.g. Myxinikela, Bardack, Reference Bardack1991; Haikouichthys, Shu et al., Reference Shu, Conway Morris, Han, Zhang, Yasui, Janvier, Chen, Zhang, Liu, Li and Liu2003; Mesomyzon, Chang et al., Reference Chang, Zhang and Miao2006). This interpretation is not universally accepted, however, and some doubts have been expressed (e.g. Pridmore et al., Reference Pridmore, Barwick and Nicoll1997; Janvier & Arsenault, Reference Janvier and Arsenault2007; Turner et al., Reference Turner, Burrow, Schultze, Blieck, Reif, Rexroad, Bultynck and Nowlan2010). It is possible to settle this matter now by considering the cleft in detail. In all likelihood this cleft represents the optic fissure, a divide that extends across the part of the eye that forms when the optic stalk invaginates at an early stage of development to create the eye ball. As this shape is very specific to eye balls, it appears justified to interpret these as such. It may also be possible that the tissue responsible for these remains is the pigment layer, as Lindgren et al. (Reference Lindgren, Uvdal, Sjövall, Nilsson, Engdahl, Schultz and Thiel2012) have recently demonstrated for the eye stain in an Eocene gnathostome.
Should there still remain doubts over the presence of optic fissures in these eye stains, it should be pointed out that this feature can be readily observed in photographs in works on other early vertebrates, although it has not yet been recognised as such (e.g. in conodonts, Aldridge et al., Reference Aldridge, Briggs, Smith, Clarkson and Clark1993, fig. 2, 4; Gabbott et al., Reference Gabbott, Aldridge and Theron1995, fig. 3; in Jamoytius, Ritchie, Reference Ritchie1968, pl. 5.1; Sansom et al., Reference Sansom, Freedman, Gabbott, Aldridge and Purnell2010, text-fig. 2e, f; possibly also Metaspriggina, Conway Morris & Caron, Reference Conway Morris and Caron2014, fig. 4d; Haikouichthys, Shu et al., Reference Shu, Conway Morris, Han, Zhang, Yasui, Janvier, Chen, Zhang, Liu, Li and Liu2003, fig. 1a,b). This is quite surprising as some of those specimens, and conodonts in particular (see Knell, Reference Knell2013), should be among the best-studied fossils in the history of palaeoichthyology.
?Sclerotic ring
Description: The remains of a small collapsed ring occur besides the opening of one of the eyes in the holotype (Fig. 2c,d). Many random shapes occur within these rocks, so by itself this is not particularly noteworthy. However, this object recalls the ‘doughnut-shaped structure’ recorded from the head region of several specimens of Euphanerops (see Janvier & Arsenault, Reference Janvier and Arsenault2007, fig. 38). Their relative position and size, in relation to the paired head stains, appear to correspond.
Interpretation: The dorsoventrally flattened nature of this specimen of Ciderius cooperi gen. nov., sp. nov. suggests that an interpretation as a sclerotic ring is the most likely. This is indicated by the position and size of this ring in comparison to the opening of the adjacent eye stain. In view of the fact that this feature is visible only in a single specimen of the new taxon, more material is needed to verify its presence.
Parachordals
Description: A pair of elongate and outwardly arched mineralisations are located near the sagittal plane at approximately 20% of the body length (Fig. 3; pc).

Fig. 3. Specimen of Ciderius cooperi gen. nov., sp. nov., with selected details. A. Near-complete body (MAB 6003, paratype); B. Drawing of A, with highlighted parachordals (pc), horizontal black lines (hbl, ?hbl), sagittal black line (sbl), bottle-shaped object (bot) and longitudinal stripes (lst). Lower Wenlock, Fish Bed Formation (and equivalent horizons), Midland Valley of Scotland.
Interpretation: Paired mineralisations have been recorded from within the head region of the lamprey Priscomyzon and the anaspid Lasanius, where they have been interpreted as remains of the otic capsules (Gess et al., Reference Gess, Coates and Rubidge2006; van der Brugghen, Reference van der Brugghen2011). However, in these instances the mineralisations are rounded rather than elongated and are not as closely grouped. It is more likely that in Ciderius gen. nov. they represent mineralised remains of the parachordals on account of their arched shape, position at the presumed posterior of the cranium and close proximity to the sagittal plane.
Branchial system
Description: There are several long horizontal black lines that seem to be connected at intervals by more numerous, thinner vertical ones (Fig. 4). The exact arrangement of these black lines is unclear because there are very few specimens revealing enough detail. However, the area in which they occur is evident as it is often marked by degraded remains. Each side of the body contains a grouping of black lines. They originate a short distance anterior to the paired mineralisations, extend across roughly half the length of the body and seem to terminate slightly posterior to the bottle-shaped object. The posterior extent is not obvious; here particularly poorly defined remains can be discerned (see Figs 3 and 4; ?hbl).

Fig. 4. A relatively detailed section of the branchial region of Ciderius cooperi gen. nov., sp. nov. A. Two partial individuals (MAB 6004, paratypes), the larger of which misses the head and tail region; B. Drawing of A, with highlighted horizontal black lines (hbl, ?hbl) and vertical black lines (vbl) of the presumed gill apparatus, the sagittal black line (sbl), and longitudinal stripes (lst). Lower Wenlock, Fish Bed Formation (and equivalent horizons), Midland Valley of Scotland.
Interpretation: Black lines have been reported from within the branchial region of Euphanerops, their arrangement suggesting that they could represent the efferent and afferent branchial arteries or the efferent branchial veins (see Janvier & Arsenault, Reference Janvier and Arsenault2007; ‘black lines’). This interpretation could be applied to Ciderius gen. nov. as the black lines occur at what appears to be the same position. Furthermore, there are other black lines within the body which seem to indicate that blood vessels have an exceptional preservation potential (see below, ‘Other black lines’).
Digestive apparatus
Description: A tarry bottle-shaped object is located slightly beyond half the body's length (Figs 1 and 3). It is often filled, in part or completely, with sediment composed of grains that are finer and more homogeneous than the surrounding matrix.
Interpretation: This object correlates with the ‘visceral cavity’ recorded in Euphanerops by Janvier & Arsenault (Reference Janvier and Arsenault2007), which they presumed housed the stomach, intestine, liver, kidneys and gonads. Instead, it is suggested that only the stomach and/or intestine is represented as the sediment, presumably ingested content, occasionally fills the bottle-shaped contour of this object entirely. What remains to be seen is which kind of digestive apparatus this represents. The leaf-shaped sedimentary infill of Endeiolepis, which can be regarded as the equivalent structure, has been reported to contain a spiral section in one specimen (see Arsenault & Janvier, Reference Arsenault and Janvier2010). It has been suggested by Arsenault & Janvier that this coiled section may represent a part of a spiral intestine. Examination of the Ciderius gen. nov. material did not reveal such a coiled region. However, there is some other, although rather indirect, evidence to put forward which could support this idea. An unusual example of the Middle Devonian lungfish Dipterus has been found which also shows a bottle-shaped object (see Fig. 5). Although no coils are visible in this specimen, the tapering tube terminates around the level of the anal fin. This specimen is of interest because extant lungfishes are known to possess a spiral valve intestine instead of a stomach (e.g. see Hassanpour & Joss, Reference Hassanpour and Joss2009, Icardo et al., Reference Icardo, Wong, Colvee, Loong and Ip2010). If this is also true for Dipterus, which seems reasonable to assume, then its spiral intestine is preserved as a bottle-shaped impression that is flattened in the same way as Ciderius gen. nov., and also does not display a coiled structure.

Fig. 5. Specimen of Dipterus valenciennesi showing a bottle-shaped object. The white area may be the flattened impression of a spiral valve intestine, an interpretation which could also apply to the bottle-shaped object in Ciderius gen. nov., sp. nov. (van der Brugghen Collection, no. VDB4DV0114), Eifelian, Achanarras Limestone Member, Lower Caithness Flagstone Group; Achanarras Quarry, Caithness, Scotland.
Other ‘black lines’
Description: Apart from the branchial region there are two other areas in which black lines have been found. The most commonly preserved is a long black line located on the sagittal plane (see Figs 3 and 4). It appears to originate near the level of the parachordals, extends across the trunk and terminates in the tail region. The features of Euphanerops termed ‘black line 1’ and ‘black line 2’ by Janvier & Arsenault (Reference Janvier and Arsenault2007) seem to match the position and thickness of the ‘sagittal black line’ of Ciderius gen. nov. This may confirm their suggestion that these two black lines in Euphanerops could be part of a single continuous structure. The other black lines of Ciderius gen. nov. are known from a single example visible on a detached sedimentary infill of the digestive apparatus (see Fig. 6). Much of the infill is covered by minute branching black lines. These lines are composed of small black dots that are similar to the degraded preservation of the sagittal black line and branchial black lines, but also to some of the ‘black lines’ recorded in Euphanerops by Janvier & Arsenault (Reference Janvier and Arsenault2007).

Fig. 6. Detached sedimentary infill of the ‘stomach’ of Ciderius cooperi gen. nov., sp. nov., showing possible blood vessels. A. Sedimentary infill covered by black lines; B. Drawing of A highlighting a network of larger and smaller branching black lines, the likely remains of blood vessels. Lower Wenlock, Fish Bed Formation (and equivalent horizons), Midland Valley of Scotland.
Interpretation: Most black lines of Euphanerops are assumed to represent major blood vessels (Janvier & Arsenault, Reference Janvier and Arsenault2007). This interpretation could also be applied to Ciderius gen. nov. because the sagittal black line and the above-mentioned branchial black lines occur within areas where larger blood vessels could be expected. The sagittal black line would then correspond to the dorsal aorta. An alternative explanation, that the sagittal black line represents the notochord or notochordal sheath, is less favoured. In Euphanerops, of which the smaller individuals seem to be taphonomically similar, the notochord is reportedly unpreserved. Its absence is possibly marked by an empty region between the arcualia (see Janvier & Arsenault, Reference Janvier and Arsenault2007). The minute branching black lines could be interpreted as blood vessels that lined the walls of the stomach and/or intestine. As to the matter why these delicate blood vessels could be so well preserved, when larger ones are not, it is likely that the fine-grained sediment on which it is imprinted has enabled a localised improvement of visible detail.
Longitudinal stripes
Description: A series of vertical rows of short longitudinal stripes extend across the sagittal plane of the posterior half of the body (see Figs 3 and 4). There are approximately 20–30 rows. The number of stripes per row differs, with one or two at the anterior, gradually increasing posteriorly to around 10 and then decreasing in number approaching the tail. The ‘sagittal black line’ seems to travel through the rows, with roughly one half of the stripes occurring ventral and the other half dorsal to this structure. It is worth noting that while the term ‘stripes’ is used to describe the individual components making up the rows, some of these objects show what could be a more complex structure. The exact shape remains unclear because it cannot yet be correlated in any two stripes.
Interpretation: Interpreting these objects is problematic. While they are frequently visible in Ciderius gen. nov., they have not been observed in the taphonomically similar Devonian euphaneropids. There would also appear to be nothing comparable in extant jawless fishes or in the early vertebrate fossil record. They are probably located at nearly the same position as the dorsal and ventral arcualia of Euphanerops (Janvier & Arsenault, Reference Janvier and Arsenault2007), yet their longitudinal direction and vertical repetition do not correspond. Perhaps their possible coincidence with the myomeres suggests an association. Again, more material is needed to clarify the precise arrangement of these objects and to reveal the structure of the individual stripes.
Reconstruction
Even though hundreds of specimens have been collected, a reconstruction of the body is hampered by the absence of individuals that show skin impressions, traces of fins and the head region in lateral aspect. This does not allow a three-dimensional reconstruction to be made with any confidence. At the moment the best rendition of the anatomy is the one shown in Figs 1B and 3B.
Further discussion
Comparison
Characters in common between Ciderius gen. nov. and other euphaneropids are the paired eyes, a barrel-shaped object, well-preserved parachordals and a greatly elongated branchial apparatus. Additionally, Euphanerops also displays a bottle-shaped digestive apparatus (see Janvier & Arsenault, Reference Janvier and Arsenault2007, fig. 13a). Ciderius gen. nov. differs mainly from other euphaneropids by the presence of longitudinal stripes and the apparent absence of anal and caudal fins. Whether these differences can be ascribed to taphonomy or phylogeny remains to be seen. A lack of fins may, after all, seem surprising when the soft-bodied anaspid Lasanius, from the same horizon as Ciderius gen. nov., quite often displays a delicate caudal fin membrane. However, it is possible that skin pigments are responsible for revealing this feature in Lasanius because the rest of its body outline usually is preserved as well. This is unlike Ciderius gen. nov., in which the absence of a body outline suggests that it did not possess skin pigments. Any additional differences between Ciderius gen. nov. and other euphaneropids are not evident in the current material due to the lack of detail shown by these soft-tissue remains. The relationships of euphaneropids to other groups also remain unclear. These animals, with their three head stains and greatly elongated branchial apparatus, do not appear to resemble any other major early vertebrate group closely. The single Jamoytius kerwoodi is the only other fossil that could have a similar arrangement.
Is Jamoytius a euphaneropid?
Jamoytius occurs within the same Silurian inlier as Ciderius gen. nov., although reportedly in a slightly older unit (see Märss & Ritchie, Reference Märss and Ritchie1998). There is a chance that Jamoytius is a euphaneropid, or is closely related to this group, even though much of the anatomy of both forms remains unknown. Jamoytius also possesses at least three anterior head stains which may be of the same nature as the ones described here. The fourth head stain reported in Jamoytius by Sansom et al. (Reference Sansom, Freedman, Gabbott, Aldridge and Purnell2010) is intriguing, but it requires further testing while taking into account the barrel-shaped object revealed here. Other than the head stains, a further comparison is problematic because all work published on Jamoytius is based on relatively few incomplete specimens. Was the caudal fin of the same euphaneropid type with a large epichordal lobe? Ritchie (Reference Ritchie1968) showed only a small part of it and the full extent of the tail remains unclear. There is also the question of how many branchial openings Jamoytius possessed. Were there just 15–17, as reported by Ritchie (Reference Ritchie1984), or were there more, as in euphaneropids? These missing data beg for new material, especially since the fossil bed containing Jamoytius has excellent preservation potential. This can be seen in the exceptionally preserved phosphatised muscle tissue that has been recorded in at least one member of the Jamoytius Horizon (see van der Brugghen et al., Reference van der Brugghen, Schram and Martill1997). Unfortunately, the only locality that yielded Jamoytius has been closed since 1995 and therefore its affinity will remain unclear.
Conclusions
As the earliest known euphaneropid, Ciderius gen. nov. demonstrates that this group went largely unchanged between the Lower Silurian and Upper Devonian, a time when jawed fishes became dominant and many previously successful groups were driven into isolated niches or perished. The success of the Euphaneropidae may be based on an efficient feeding strategy which utilised their greatly elongated branchial apparatus to filter large amounts of water for minute food particles.
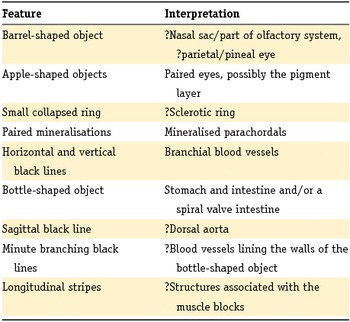
Much of the anatomy remains unclear for now (see Table 2). The barrel-shaped object is particularly enigmatic, as is the position and configuration of the oral opening. Where hard-bodied fossils will usually demonstrate most of the morphology with just a few complete specimens, these soft-tissue fossils are less revealing. More material of Ciderius gen. nov. is needed to clarify its anatomy but, with several hundred specimens collected during this study, it is becoming increasingly difficult to find specimens that display new details. Having said that, given their exquisite preservation potential, new material of Euphanerops and Jamoytius is most likely to provide answers to the vexing questions surrounding the family Euphaneropidae.
Table 2. A summary of the structures observed in Ciderius cooperi gen. nov., sp. nov. and their interpretations.
Acknowledgements
I thank René H.B Fraaije (Boxtel) for stimulating discussions and for suggesting improvement to an earlier typescript, John W.M. Jagt (Maastricht) for reviewing the typescript and providing useful comments, and Philippe Janvier (Paris) for useful discussions, review comments and for sharing with me the correct identification of the parachordal cartilages.








