



For pigs reared in modern swine facilities, weaning is a source of significant stress resulting from the exposure to environmental, psychological and nutritional stressors(Reference Pluske, Hampson and Williams1,Reference Lallès, Boudry and Favier2) . This stress results in an immediate reduction in feed intake, leading to disturbances in gastrointestinal physiology, microbiology and immunology(Reference Pluske3). This post-weaning intestinal dysfunction creates an opportunity for colonisation by pathogenic bacteria, which when combined with reduced gut barrier function and inflammation leads to post-weaning diarrhoea(Reference Pluske, Hampson and Williams1). The maturation of the gastrointestinal tract (GIT) is still underway during the early post-weaning period, and the optimal functioning of the GIT is a crucial factor affecting pig performance post-weaning(Reference Moeser, Pohl and Rajput4). A healthy GIT encompasses several factors such as efficient nutrient digestion and absorption, energy generation, a stable diverse microbiota, effective barrier function, effective mucosal immune system and the balanced interaction of these factors(Reference Pluske, Turpin and Kim5). In contrast, post-weaning intestinal dysfunction is characterised by atrophy of the small intestinal architecture, up-regulation of pro-inflammatory cytokines, proliferation of pathogenic bacteria and diarrhoea leading to reduced feed intake and performance(Reference O’Doherty, Bouwhuis and Sweeney6).
Antibiotic growth promoters were included in weaner pig diets to overcome post-weaning intestinal dysfunction. However, this management strategy was banned in the EU in 2006 (EC regulation no. 1831/2003). The inclusion of pharmacological doses of zinc oxide (ZnO) has successfully reduced diarrhoea and improved growth performance in the post-weaning period(Reference Kim, Hansen and Mullan7). However, concerns regarding Zn accumulation in soils and the relationship between ZnO usage and the rise in antibiotic resistance(Reference Long, Chen and Zhang8) have resulted in the planned phasing out of pharmacological doses of ZnO in the EU by 2022 (Commission Implementing Decision of 26 June 2017, C(2017) 4529 Final). The EU Commission has also agreed to impose further restrictions on the use of antibiotics in farm animals from 2022, including a ban on the preventative use of antibiotics in groups of animals(9). Thus, there is an urgent requirement to find alternative feed supplements to prevent the post-weaning growth check and intestinal dysfunction in the absence of in-feed medications. As feed intake is notoriously low in the immediate post-weaning period, the use of supplements which confer benefits to GIT health and prevent intestinal dysfunction may be one way to enhance growth performance in the post-weaning period.
Brown seaweeds are a source of natural bioactive polysaccharides such as laminarin(Reference Deville, Gharbi and Dandrifosse10). Laminarin is composed of 1,3-linked β-d-glucose residues with different degrees of branching at β-(1,6) with the extent of branching influencing its solubility(Reference Rioux, Turgeon and Beaulieu11). As laminarin has previously demonstrated anti-inflammatory and anti-bacterial activities(Reference Walsh, Sweeney and O’Shea12), it may be considered as a dietary intervention to prevent intestinal dysfunction and thus improve post-weaning performance. However, the polysaccharide content and composition of seaweed extracts can vary with season of harvest, macroalgal species and extraction procedure(Reference Garcia-Vaquero, Rajauria and O’Doherty13). Furthermore, the production of purified extracts is both costly and inefficient in terms of yields. Thus, the objective of the present study was to investigate the optimum inclusion level of laminarin from a 65 % purified laminarin extract on the growth performance of weaned pigs. A second objective was to identify the effects of the optimum inclusion level of laminarin on weaning associated intestinal dysfunction using the following parameters: small intestinal morphology, the expression of genes involved in nutrient digestion and absorption, inflammation, mucin production, pathogen recognition and tight junctions, populations of selected bacteria and volatile fatty acid (VFA) concentrations. It was hypothesised that laminarin would enhance growth performance in the first 14 d post-weaning, by enhancing intestinal morphology, reducing Enterobacteriaceae, increasing the expression of nutrient transporters and digestive enzymes and reducing the expression of pro-inflammatory cytokines.
Materials and methods
All experimental procedures described in this work were approved under University College Dublin Animal Research Ethics Committee (AREC-17-19-O’Doherty) and conducted in accordance with Irish legislation (SI no. 534/2012) and the EU directive 2010/63/EU for animal experimentation.
Experimental design and diets
The experiment comprised four dietary treatments: (T1) basal diet; (T2) basal diet + 100 parts per million (ppm) laminarin; (T3) basal diet + 200 ppm laminarin and (T4) basal diet + 300 ppm laminarin. The inclusion levels of laminarin were determined based on the previous work by Smith et al.(Reference Smith, O’Doherty and Reilly14) where no further benefit was observed when the dietary laminarin inclusion level was increased from 300 to 600 ppm. Ninety-six healthy piglets (progeny of meatline boars × (large white × landrace sows)) with an average weight of 8·4 (sd 1·09) kg were sourced from a commercial pig farm at weaning (28 d of age) and housed in pens of three. The pigs were blocked based on weaning weight, litter of origin and sex and within each block assigned to one of the four dietary treatments (eight replicates/treatment). The basal diet contained 14·95 MJ/kg digestible energy, 190 g/kg crude protein and 13·5 g/kg total lysine. All amino acid requirements were met relative to lysine(15). The ingredient composition and chemical analysis of the dietary treatments are presented in Table 1. The laminarin-rich extract was sourced from BioAtlantis Ltd. The extract was prepared by using water as an extraction solvent under optimum heating conditions (undisclosed method). A single extraction was performed from Laminaria spp. to produce the extract which contained 650 g of laminarin/kg DM, 181 g of fucoidan/kg DM, 4 g phlorotannins/kg DM, 60 g mannitol/kg DM, 28 g alginates/kg DM and 77 g ash/kg DM. The appropriate quantity of the laminarin-rich extract was added to the basal diet to achieve 100, 200 or 300 ppm laminarin inclusion levels.
Table 1. Ingredient and chemical composition of basal diet*

ppm, parts per million.
* Treatments: (1) basal diet; (2) basal diet + 100 ppm laminarin; (3) basal diet + 200 ppm laminarin; (4) basal diet + 300 ppm laminarin.
† Calculated for tabulated nutritional composition(Reference Sauvant, Perez and Tran43). Provided (mg/kg complete diet): Cu, 100; Fe, 140; Mn, 47; Zn, 120; iodine, 0·6; Se, 0·3; retinol, 1·8; cholecalciferol, 0·025; α-tocopherol, 67; phytylmenaquinone, 4; cyanocobalamin, 0·01; riboflavin, 2; nicotinic acid, 12; pantothenic acid, 10; choline chloride, 250; thiamine, 2; pyridoxine, 0·015. Celite included at 300 mg/kg complete diet.
Housing and animal management
The pigs were housed in fully slatted pens (1·7 × 1·2 m). They were weighed at the beginning of the experiment (d0; day of weaning) and on days 7 and 14. The ambient environmental temperature within the house was thermostatically controlled at 30°C for the first 7 d and then reduced by 2°C for the remainder of the 2nd week, and the humidity was maintained at 65 %. Feed in meal form and water were available ad libitum from four-space feeders and nipple drinkers. Aluminium trays were placed under feeders to catch any displaced feed and avoid feed wastage. Every day throughout the experiment, faecal scores (FS) were recorded in the individual pens by the same operator on a scale ranging from 1 to 5. The following scoring system was used: 1 = hard, firm faeces; 2 = slightly soft faeces; 3 = soft, partially formed faeces; 4 = loose, semi-liquid faeces and 5 = watery, mucous-like faeces(Reference Walsh, Sweeney and O’Shea16).
Sample collection
On day 15, eight pigs (1 pig/pen) from the basal group and best-performing laminarin treatment (300 ppm laminarin) group (based on ADG and ADFI) received a lethal injection with pentobarbitone sodium (Euthatal Solution, 200 mg/ml; Merial Animal Health) at a rate of 0·71 ml/kg body weight to the cranial vena cava to humanely kill the animals. Euthanasia was completed by a competent person in a separate room away from sight and sound of the other pigs. The entire intestinal tract was immediately removed. Sections from the duodenum (10 cm from the stomach), the jejunum (60 cm from the stomach) and the ileum (15 cm from the caecum) were cut out and fixed in 10 % neutral-buffered formalin. Digesta from the caecum and colon was collected into sterile containers (Sarstedt) and immediately frozen for subsequent quantification of total bacteria, Enterobacteriaceae, Bifidobacterium spp. and Lactobacillus spp. using quantitative PCR. In addition, tissue samples were taken from the duodenum, jejunum, ileum and colon to measure the gene expression of cytokines, digestive enzymes, nutrient transporters, mucins, tight junction components, pathogen recognition receptors, transcription regulators, appetite regulators, growth factors, kinases, ligand-dependent nuclear receptors, suppressors of cytokine signalling, peptidases, transmembrane receptors and viral defence genes using the Nanostring nCounter, which detects and counts target mRNA molecules using target-specific, colour-coded probe pairs(Reference Geiss, Bumgarner and Birditt17). Tissue sections of 1 cm2 from the duodenum, jejunum, ileum and colon were cut out, emptied by dissecting them along the mesentery and rinsed using sterile PBS (Oxoid). The tissue sections were stripped of the overlying smooth muscle before storage in 5 ml RNAlater® solution (Applied Biosystems) overnight at 4°C. The RNAlater® was removed before storing the samples at −80°C.
Gut morphological analysis
Preserved duodenal, jejunal and ileal tissue samples were prepared using standard paraffin-embedding techniques. The samples were sectioned at a thickness of 5 μm and stained with haematoxylin–eosin. Villus height (VH) and crypt depth (CD) were measured in the stained sections (4× objective) using a light microscope fitted with an image analyser (Image-Pro Plus; Media Cybernetics). Measurements of fifteen well orientated and intact villi and crypts were taken for each segment. The VH was measured from the crypt–villus junction to the tip of the villus, and CD was measured from the crypt–villus junction to the base. Results are expressed as mean VH or CD in μm.
Gene expression
RNA extraction
Total RNA was extracted from duodenal, jejunal, ileal and colonic tissue using TRIreagent (Sigma-Aldrich) according to the manufacturer’s instructions. The crude RNA extract was further purified using the GenElute Mammalian Total RNA Miniprep Kit (Sigma-Aldrich) according to the manufacturer’s instructions. A DNase removal step was included using an on-Column DNase 1 Digestion Set (Sigma-Aldrich). The total RNA was quantified using a Nanodrop-ND1000 Spectrophotometer (Thermo Scientific), and the purity was assessed by determining the ratio of the absorbance at 260 and 280 nm. All total RNA samples had 260/280 nm ratios above 2·0.
Nanostring nCounter analysis
The small intestinal (duodenal, jejunal and ileal) tissues and colonic tissue were analysed using the Nanostring nCounter Analysis System (Nanostring Technologies). Two custom nCounter panels one for the small intestine and one for the colon were designed by our group and manufactured by Nanostring Technologies. The genes measured in the small intestine and large intestine are presented in the Supplementary material (online Supplementary Tables S1 and S2, respectively). The small intestinal codeset contained thirty-two target genes and five reference genes and the large intestinal codeset contained fifty-two target genes and eight reference genes. Both panels contained six positive and eight negative system controls.
The expression profile of all target genes was performed for each sample in a single multiplexed hybridisation reaction, as originally described by Geiss et al.(Reference Geiss, Bumgarner and Birditt17). Briefly, prior to analysis, all samples were calibrated to 20 ng/μl using a Qubit fluorometer (Thermo Fisher Scientific). For the hybridisation reaction, a master mix was created by adding 70 μl of hybridisation buffer to the reporter codeset, as per manufacturer instructions. Master mix (8 μl), sample (5 μl) (total RNA concentration 100 ng) and capture probeset (2 μl) were added to each reaction tube and inverted to mix and spun down before incubation at 65°C for 20 h in a Bio-rad thermocycler. Post-hybridisation processing was performed within the Nanostring nCounter prep station liquid handling robot which involves the removal of excess unbound probes and immobilisation of samples onto the internal surface of the sample cartridge. Following this imaging is completed in the digital analyser, which collects data by taking images of the immobilised fluorescent reporters in the sample cartridge with a CCD camera through a microscope objective lens. The analysis and normalisation of the raw Nanostring data were performed using nSolver Analysis Software v4.0 (Nanostring Technologies). Background was corrected for using background thresholding in which a threshold value is set and all counts which fall below that value are adjusted to match it. The background threshold value was estimated using the average count of the negative control probes in every reaction plus 2 standard deviations(Reference Vigors, JV and Ryan18). Target genes with raw counts below the threshold in more than two thirds of samples were excluded from the analysis. Raw counts were normalised using a combination of positive control normalisation and codeset content normalisation. The former accounts for errors such as pipetting errors, lot-to-lot variation in nCounter preparation plates and nCounter cartridges, while the latter uses reference/housekeeping genes to account for variability in the quantity and quality of sample RNA. Nanostring results (raw and normalised counts) were produced from RCC files using nSolver software V4.0.
Microbial DNA extraction and quantitative PCR assay
Microbial genomic DNA was extracted from the caecal and colonic digesta samples using a QIAamp DNA stool kit (Qiagen) in accordance with the manufacturer’s instructions. The quantity and quality of DNA were assessed using a Nanodrop spectrophotometer (Thermo Scientific). For the quantitative PCR, standard curves were prepared with pooled aliquots of caecal and colonic digesta DNA as described previously by O’Shea et al.(Reference O’Shea, Sweeney and Bahar19). Domain, genus and family-specific primers were used to amplify the 16s rRNA gene. Primers for total bacteria, Lactobacillus spp., Bifidobacterium spp. and Enterobacteriaceae are presented in online Supplementary Table S3. The selected bacterial groups were estimated based on gene copy number in the digesta using quantitative PCR on the 7500 Fast Real-Time PCR System (Applied Biosystems). Quantitative PCR was carried out in a final reaction volume of 20 μl containing 3 μl template DNA, 1 μl of forward and reverse primers (100 pm), 10 μl SYBR Green PCR Master Mix (Applied Biosystems) and 5 μl nuclease-free water. The thermal cycling conditions involved an initial denaturation step at 95°C for 10 min followed by forty cycles of 95°C for 15 s and 65°C for 1 min. Dissociation curves confirmed the specificity of the final PCR products. All samples were prepared in duplicate, and the mean threshold cycle (Ct) value was used for calculations. The estimates of gene copy number for selected bacteria were log-transformed and are presented as log gene copy number per g of digesta.
Volatile fatty acids
The VFA concentrations in the caecal and colonic digesta were determined using GLC according to the method described by Pierce et al.(Reference Pierce, Sweeney and Callan20). A 1 g sample was diluted with distilled water (2·5 × weight of sample) and centrifuged at 1400 g for 10 min (Sorvall GLC–2B laboratory centrifuge, DuPont). The subsequent supernatant (1 ml) and internal standard (1 ml; 0·05 % 3-methyl-n-valeric acid in 0·15 m oxalic acid dihydrate) were mixed with 3 ml of distilled water. The reaction mixture was centrifuged at 500 g for 10 min, and the supernatant was filtered through 0·45 polytetrafluoroethylene syringe filter into a chromatographic sample vial. An injection volume of 1 μl was injected into a Varian 3800 GC equipped with an EC™ 1000 Grace column (15 m × 0·53 mm I.D) with 1·20 μm film thickness. The temperature programme set was 75–95°C increasing by 3°C/min and 95–200°C increasing by 20°C/min, which was held for 0·50 min. The detector and injector temperature were 280 and 240°C, respectively, while the total analysis time was 12·42 min.
Feed analysis
The feed samples were milled through a 1 mm screen (Christy and Norris hammer mill). The DM of the feed was determined after drying overnight at 104°C. Crude ash content was determined after ignition of a known weight of concentrate in a muffle furnace (Nabertherm) at 550°C for 6 h. The crude protein content was determined as Kjeldahl N × 6·25 using the LECO FP 528 instrument. The neutral-detergent fibre content was determined according to Van Soest et al.(Reference Van Soest, Robertson and Lewis21).
Statistical analysis
For sample size determination, the following power analysis formula was used: replication per treatment = 15·7 × (CV/% difference)2. The coefficient 15·7 is derived from statistical tables with a fixed experimental power of 80 % and a significance value of P < 0·05. The highest CV observed in comparable work was 14 % for ADG(Reference Heim, Walsh and Sweeney22). In the present study, D = 20 % is the difference that would be important to detect in selected variables including body weight change, gene expression and FS. Therefore, substituting the values into the above-mentioned formula:
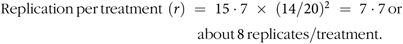 $${\rm{Replication}}{\mkern 1mu} \ {\rm{per}}{\mkern 1mu} \ {\rm{treatment}}{\mkern 1mu} {\mkern 1mu} \left( r \right)\; = \;15\cdot7\; \times \;{(14/20)^2}\; = \;7\cdot7{\mkern 1mu} \ {\rm{or}} \ {\mkern 1mu} 117 - 7 \ {\rm{about}}\ \ {\mkern 1mu} 8\ {\mkern 1mu} {\rm{replicates}}/ \ {\rm{treatment}}.$$
$${\rm{Replication}}{\mkern 1mu} \ {\rm{per}}{\mkern 1mu} \ {\rm{treatment}}{\mkern 1mu} {\mkern 1mu} \left( r \right)\; = \;15\cdot7\; \times \;{(14/20)^2}\; = \;7\cdot7{\mkern 1mu} \ {\rm{or}} \ {\mkern 1mu} 117 - 7 \ {\rm{about}}\ \ {\mkern 1mu} 8\ {\mkern 1mu} {\rm{replicates}}/ \ {\rm{treatment}}.$$
The resulting data were initially checked for normality using the univariate procedure in SAS. The performance data and FS data were analysed by repeated measures using the mixed procedure of SAS and the model included fixed effects of treatment, time and their associated interactions. The initial weight was used as a covariate for the performance data. The data on intestinal morphology, microbial populations, gene expression and VFA were analysed using the GLM procedure of SAS, using weight at slaughter as a covariate. The model assessed the effect of treatment, followed by Bonferroni’s test with the pig being the experimental unit. The probability level that denoted significance was P < 0·05. Data are presented as least-square means with their standard errors of the mean.
Results
Performance and faecal scores
The effects of laminarin supplementation on ADG, ADFI, G:F ratio and FS are presented in Table 2. From d0–7, there was no significant effect of laminarin on ADG or ADFI. From d7–14 to d0–14, the 300 ppm laminarin supplemented pigs had a higher ADG (P < 0·05) than the basal group and higher ADFI (P < 0·05) than all other groups. The G:F ratio was unaffected by treatment. FS remained within the healthy range, and there was no difference in FS between the basal and the laminarin supplemented diets (P > 0·05) during the 14-d experimental period.
Table 2. Effect of increasing inclusion levels of laminarin on pig growth performance and faecal scores
(Least-square mean values with their standard errors)

ppm, parts per million; ADG, average daily gain; ADFI, average daily feed intake; G:F, gain to feed ratio; FS, faecal score.
a,b Mean values within a row with unlike superscript letters were significantly different (P < 0·05).
* A total of eight replicates were used per treatment group (replicate = pen, 3 pigs/pen).
Small intestinal morphology
The effects of laminarin supplementation on villous height and CD in the small intestine are presented in Table 3. Pigs offered the 300 ppm laminarin diet had increased villous height in the duodenum and jejunum (P < 0·05) and increased CD in the duodenum (P < 0·01) compared with pigs offered the basal diet. No effect on morphology was observed in the ileum (P > 0·05).
Table 3. Effect of laminarin on villus height (VH) and crypt depth (CD) in the small intestine
(Least-square mean values with their standard errors)

ppm, parts per million.
* A total of eight replicates were used per treatment.
Microbiology and volatile fatty acids
The effects of laminarin supplementation on selected microbial populations in the caecum and colon are presented in Table 4. There was a reduction in the Enterobacteriaceae population in the caecum (P < 0·05) and an increase in Lactobacillus spp. in the colon (P < 0·05) of laminarin supplemented pigs compared with the basal group. There was no effect of laminarin on the numbers of total bacteria, Lactobacillus spp. or Bifidobacterium spp. in the caecum (P > 0·05) or the numbers of total bacteria, Enterobacteriaceae or Bifidobacterium spp. in the colon (P > 0·05).
Table 4. Effect of laminarin on selected microbial populations in the caecal and colonic digesta
(Least-square mean values with their standard errors)

ppm, parts per million; GCN, gene copy number.
* A total of eight replicates were used per treatment.
The effects of laminarin supplementation on the concentrations and molar proportions of VFA are presented in Table 5. In the colon, pigs offered laminarin had increased concentrations of total VFA (P < 0·05) and acetate (P < 0·05) and an increase in the molar proportion of butyrate (P < 0·01).
Table 5. Effect of laminarin inclusion on total and molar proportions of volatile fatty acids (VFA) in colonic digesta
(Least-square mean values with their standard errors)

ppm, parts per million.
* A total of eight replicates were used per treatment.
Gene expression
While the Nanostring nCounter panels used in the present study assessed the expression of a broad range of genes, only the genes with relevance to gastrointestinal functionality in the post-weaning period are included in the results. The genes that were differentially expressed are presented in Table 6 with all expression data for relevant genes presented in the Supplementary material (online Supplementary Tables S4–S7).
Table 6. Effect of laminarin inclusion on the expression of nutrient transporters in the duodenum, jejunum and ileum; the expression of immune markers in the duodenum, ileum and colon; and appetite regulators in the ileum
(Least square means with their standard errors)

ppm, parts per million; SLC2A8, GLUT 8; CLDN5, claudin 5; TJP1, tight junction protein 1; TGFB1, transforming growth factor beta 1; SLC2A2, GLUT 2; SLC2A7, GLUT 7; SLC6A19, neutral amino acid transporter; SLC15A1, peptide transporter 1; FABP2, fatty acid binding protein 2; SLC5A8, sodium monocarboxylate transporter; CCK, cholecystokinin; MUC1, mucin 1; MUC2, mucin 2; CLDN3, claudin 3; TJP1, tight junction protein 1; TLR2, toll-like receptor 2; SLC16A1, monocarboxylate transporter 1; CLEC7A, c-type lectin domain containing 7a; PPARG, PPAR receptor.
* A total of eight replicates were used per treatment.
Nutrient and volatile fatty acids transporter gene expression
In the duodenum, laminarin supplementation increased the expression of the GLUT 8 (SLC2A8; P < 0·05). In the jejunum, laminarin supplementation increased the expression of GLUT 2 (SLC2A2; P < 0·05), GLUT 7 (SLC2A7; P < 0·05), peptide transporter 1 (SLC15A1; P < 0·05), neutral amino acid transporter 19 (SLC6A19; P = 0·05) and fatty acid binding protein 2 (FABP2; P < 0·05). In the ileum, laminarin supplementation reduced the expression of neutral amino acid transporter 19 (SLC6A19) and sodium monocarboxylate transporter (SLC5A8/SMCT1) (P < 0·05).
In the colon, laminarin supplementation increased the expression of monocarboxylate transporter 1 (SLC16A1; P < 0·05).
Appetite regulator gene expression
In the ileum, laminarin supplementation increased the expression of cholecystokinin (CCK; P < 0·05). There were no differences in the duodenum or jejunum.
Tight junction and immune marker gene expression
In the duodenum, laminarin supplementation reduced the expression of claudin-5 (CLDN5; P < 0·05), tight junction protein 1 (zona occludens 1) (TJP1; P < 0·01), NF-κB1 (P < 0·05) and transforming growth factor beta 1 (TGFB1; P < 0·05). In the ileum, laminarin supplementation reduced the expression of claudin-3 (CLDN3; P < 0·05), TJP1 (P < 0·05), toll-like receptor 2 (TLR2; P < 0·05), mucin 1 (MUC1; P < 0·05) and mucin 2 (MUC2; P < 0·01).
In the colon, laminarin supplementation reduced the expression of Dectin-1/C-type lectin domain containing 7A (CLEC7A; P < 0·05), while it increased the expression of PPAR gamma (PPARG; P < 0·05).
Discussion
The hypothesis of the present study was that laminarin supplementation would improve performance and reduce intestinal dysfunction during the first 14 d post-weaning, by enhancing intestinal morphology, reducing Enterobacteriaceae, increasing the expression of nutrient transporters and digestive enzymes and reducing the expression of pro-inflammatory cytokines. The positive response observed with laminarin supplementation, including increased VH in the duodenum and jejunum, reduced caecal Enterobacteriaceae, increased colonic lactobacilli and butyrate concentration and increased expression of VFA, glucose, fatty acid and protein transporters, supports the hypothesis that laminarin supplementation reduces post-weaning intestinal dysfunction. The present study also confirms that 300 ppm is the optimum of the tested inclusion levels of laminarin, to improve post-weaning pig performance.
The increased feed intake in the 300 ppm laminarin supplemented group is a major factor driving the improved growth performance in these animals. The oral consumption of food and its physical presence in the GIT is necessary for structural and functional maintenance of the intestinal mucosa(Reference Pluske, Hampson and Williams1), as villous atrophy and crypt hyperplasia are the consequences of low feed intake post-weaning(Reference Pluske, Hampson and Williams1,Reference Lallès, Boudry and Favier2) . In the present study, laminarin supplementation also increased villous height in the duodenum and jejunum. This increase was independent of live weight. Longer villi are associated with increased digestive and absorptive function(Reference Jayaraman and Nyachoti23). In the present study, laminarin supplementation increased the expression of the glucose transporters SLC2A8/GLUT8 (duodenum), SLC2A2/GLUT2 and SLC2A7/GLUT7 (jejunum), the peptide transporter SLC15A1/PEPT1 (jejunum), the amino acid transporter SLC6A19 (duodenum and ileum) and the long-chain fatty acid transporter FABP2 (jejunum). Nutrient transporters are responsible for the transport of nutrients into the bloodstream following digestion by digestive enzymes. It is possible that the increase in sugar, fatty acid, peptide and amino acid transporters improved energy and protein utilisation in pigs supplemented with laminarin. Unfortunately, the digestibility of these nutrients was not measured in the present study, due to the low quantity of digesta in the ileum at the time of sampling. It also suggests that laminarin supplemented pigs may have adapted more quickly to their post-weaning diet which is primarily comprised of carbohydrates (wheat and maize) and protein (soya bean meal and soya protein concentrate) by up-regulating the expression of the appropriate nutrient transporters. It is commonly considered that the small intestine responds to dietary changes by altering the expression of digestive enzymes and nutrient transporters, for example, an increase in dietary carbohydrates may cause an up-regulation of hexose transporters to promote an increase in sugar absorption(Reference Drozdowski and Thomson24).
Feed intake is regulated through the release of food intake-regulatory peptides from the GIT. One such peptide is CCK, which induces satiety and reduces food intake(Reference Wang, Chandra and Samsa25,Reference Prinz and Stengel26) . The up-regulation of CCK in the ileum of laminarin supplemented pigs suggests that feed intake requirements were satisfied in this group. CCK is mainly stimulated by fats and proteins(Reference Wang, Chandra and Samsa25,Reference Prinz and Stengel26) , and the increase in FABP2 and PEPT1 in the jejunum of laminarin supplemented pigs may indicate that there was sufficient absorption of these nutrients prior to reaching the ileum.
The relevance of measuring Enterobacteriaceae populations as an indicator of pathogenic bacteria is widely debated; however, an increase in Enterobacteriaceae has been reported in diarrhoeic pigs post-weaning(Reference Dou, Gadonna-Widehem and Rome27). The Enterobacteriaceae family includes opportunistic pathogens such as Salmonella enterica serovar Typhimurium and enterotoxigenic Escherichia coli which can induce intestinal inflammation in pigs(Reference Guevarra, Lee and Lee28) and enterotoxigenic Escherichia coli is reported to be one of the main causative factors of post-weaning diarrhoea in piglets(Reference Gresse, Chaucheyras-Durand and Fleury29,Reference Rhouma, Fairbrother and Beaudry30) . The ability of β-glucans (laminarin) to agglutinate certain bacterial species and prevent their attachment to the intestinal epithelium was previously reviewed by Sweeney & O’Doherty(Reference Sweeney and O’Doherty31). Although many members of the Enterobacteriaceae family are commensals, the potential to reduce pathogenic Enterobacteriaceae without affecting beneficial bacterial populations such as the bifidobacteria and lactobacilli may have contributed to the improved growth performance in the laminarin treatment group. However, future studies with laminarin will investigate its effects on specific pathogens using 16S rRNA sequencing to fully elucidate its prebiotic and anti-microbial potential.
Of significance was the increase in lactobacilli numbers and increased concentration of total VFA, acetate and butyrate, as well as the increased expression of the butyrate transporter monocarboxylate transporter 1 (MCT1/SLC16A1)) with laminarin supplementation in the colon. Lactobacilli were enumerated as a reflection of changes in the population structure of beneficial bacteria as they can enhance host GIT health through the competitive exclusion of pathogenic bacteria, antioxidant activities and aiding the regulation of the immune system(Reference Valeriano, Balolong and Kang32). The production of VFA allows for greater energy utilisation by recovering energy from carbohydrates which were not digested in the small intestine(Reference Bedford and Gong33). VFA are essential for the growth and proliferation of colonocytes(Reference Liu34), and butyrate is the preferential energy source of the colonocytes(Reference Williams, Verstegen and Tamminga35). Butyrate has also other intestinal health-enhancing properties such as the facilitation of the absorption of water, Na and K(Reference Bedford and Gong33), regulation of gene expression by inhibiting the expression of histone deacetylases, enhancing the expression of host defence peptides, and antibacterial activity(Reference Bedford and Gong33,Reference Xiong, Tan and Song36) . As butyrate plays an important role in the maintenance of colonic health, the expression of SLC16A1 and absorption of butyrate can have an important impact on colonic homeostasis(Reference Cuff, Lambert and Shirazi-Beechey37).
Laminarin inclusion induced a number of positive immunomodulatory effects in the small intestine of the post-weaning pig. Laminarin inclusion was associated with a down-regulation of the transcription factor NF-κB1 and the growth factor TGFB1 in the duodenum. NF-κB1 induces the expression of pro-inflammatory cytokines(Reference Liu, Zhang and Joo38), and TGFB1 is involved in mucosal cell growth, differentiation, migration and epithelial restitution(Reference Xiao, Song and Jiao39). TGFB1 has been shown to be up-regulated in the 1st week post-weaning and then returns to pre-weaning levels(Reference Xiao, Song and Jiao39). The expression of genes involved in the formation of tight junctions CLDN5 (duodenum), TJP1 (duodenum and ileum) and CLDN3 (ileum) was also reduced in laminarin supplemented pigs. An increase in the expression of these genes likely functions to enhance the epithelial barrier during periods of challenge; previously, the expression of TJP1, CLDN3 and OCLN was transiently increased in heat stressed pigs and associated with increased LPS permeability and reduced villous height(Reference Pearce, Mani and Weber40). The TGFB1, CLDN3, CLDN5 and TJP1 expression data in association with the villous architecture data suggest that laminarin supplemented pigs may have achieved epithelial maturity in the small intestine faster than the basal fed animals. However, these changes in gene expression were minor, and while the reductions were statistically significant, the biological impact of their reduction is probably limited. The expression of TLR2, MUC1 and MUC2 was down-regulated in the ileum of laminarin supplemented pigs. TLR2 recognises microbial-associated molecular patterns of yeast and bacteria, and it has been previously shown to be up-regulated in the ileum immediately post-weaning and subsequently declines(Reference Tao, Xu and Wan41). Mucin secretion can be initiated by a number of factors, including microbes, microbial products, cytokines and reactive oxygen species(Reference Kim and Ho42), indicating it is up-regulated in response to challenge. Together, the reduction in mucins and TLR2 suggests reduced exposure to pathogens in laminarin supplemented animals. Considering FS remained within a healthy range throughout the experiment, it is unlikely the immune system of either group was challenged. Nonetheless, these results suggest laminarin supplementation may have beneficial immunomodulatory effects under more challenging conditions.
Conclusion
In conclusion, improved ADG and ADFI suggest 300 ppm is the optimum of the tested inclusion levels of laminarin from a laminarin-rich extract (65 % laminarin) to enhance post-weaning performance. The increased VH, expression of nutrient transporters, populations of lactobacilli and butyrate concentrations, in conjunction with reduced populations of Enterobacteriaceae and reduced expression of immune markers and pathogen recognition receptors, suggest that laminarin supplementation reduced intestinal dysfunction in the post-weaning period. Thus, laminarin has potential as an alternative to in-feed medications for piglets in the post-weaning period. Furthermore, laminarin has potential as a dietary supplement for other monogastric animals and humans for the treatment or prevention of diseases related to intestinal dysfunction.
Acknowledgements
This work was funded by the Science Foundation Ireland (SFI) (grant number: 14/IA/2548).
The authors’ contributions were as follows: R. R. performed the experiment, collected the samples, carried out the laboratory analyses and wrote the manuscript; J. V. O. D. designed the experiment, supervised data collection and statistical analyses and corrected the manuscript; T. S. designed the experiment and corrected the manuscript; S. M., K. T. and G. R. contributed to sample collection and laboratory analyses. All authors approved the final version of the manuscript.
None of the authors had a financial or personal conflict of interest in relation to the present study.
Supplementary material
For supplementary material referred to in this article, please visit https://doi.org/10.1017/S0007114519002678








