



Introduction
A current challenge in agricultural systems is to balance increases in productivity and reductions in cultivation area while also preserving the environment and generating profits for farmers (Roesch-McNally et al., Reference Roesch-Mcnally, Arbuckle and Tyndall2018). Integrated systems are techniques that provide benefits for both agriculture and livestock production, leading to economic and especially environmental gains (Santos et al., Reference Santos, Albuquerque Filho, Vilela, Ferreira, Carvalho and Viana2014).
Integrated crop–livestock systems are characterized by the use of the same production area for agriculture and livestock production, either simultaneously or in succession, to improve the use of land and of environmental resources. When properly managed, these systems can efficiently store carbon (C) and help to maintain the carbon/nitrogen (C/N) ratio of the soil via the function of the root system of plants, increasing the amount of biomass of the plant material used for mulch, which in turn is used as soil cover in no-till systems (Carvalho et al., Reference Carvalho, Avanzi, Silva, Mello and Cerri2010a).
The biomass of perennial plants used as mulch promotes high levels of organic matter (OM) in the soil, which contributes to nutrient cycling, especially the replenishment of nitrogen (N) and C, and water maintenance and improves soil properties (Ryschawy et al., Reference Ryschawy, Martin, Moraine, Duru and Therond2017). Furthermore, this soil dynamic helps to reduce the use of fertilizer, thus reducing costs and the impacts on the environment (Laroca et al., Reference Laroca, Souza, Pires, Pires, Pacheco, Silva, Wruck, Carneiro, Silva and Souza2018).
Thus, the selection of a cover crop to provide the amount of mulch biomass needed to maintain an integrated system should account for the climatic conditions of the region. In the Brazilian Cerrado (savannah), the winters are characterized by drought events and low humidity, while the summers have high temperatures and rainfall, which hinder the longevity of mulch biomass on the soil, as high temperature and humidity accelerate the decomposition rate the plant materials (Castro et al., Reference Castro, Silva, Moreira and Resende2017).
Brachiaria species are considered the primary species for the production of mulch biomass, with the potential to accumulate and release nutrients into the soil for subsequent crops (Flávio Neto et al., Reference Flávio Neto, Severiano, Costa, Guimarães Junnyor, Gonçalves and Andrade2015; Costa et al., Reference Costa, Costa, Santos, Severiano, Epifânio, Silva, Teixeira and Silva2016; Costa et al., Reference Costa, Costa, Severiano, Santos, Rocha, Souza, Brandstetter and Castro2017). However, with the emergence of new Panicum maximum cultivars, the Mombaça guinea grass (Soratto et al., Reference Soratto, Costa, Crusciol, Neto Ferrari and Moro2019) and Tamani guinea grass (Machado et al., Reference Machado, Cecato, Comunello, Concenço and Ceccon2017) have been increasingly used in integrated systems, with positive results.
In contrast to information on Brachiaria ruziziensis, which has been the most frequently used grass species in integrated systems for many years because of its high efficiency, information on the introduction of these new cultivars is scarce. As such, it is necessary to determine the potential of these forage grasses in integrated production systems for promoting increased soybean yields via biomass production compared with that of maize in succession with soybean as the second crop.
Thus, the objective of this study was to evaluate the biomass production of and total nutrient in Brachiaria spp. and P. maximum forage grasses used as mulch and soybean yields in an integrated crop–livestock system and in soybean/maize (Zea mays) succession.
Materials and methods
Description of the area
This study was conducted at the Instituto de Ciência and Tecnologia Comigo – ITC (Institute of Science and Technology Comigo) in Rio Verde, state of Goiás (GO), Brazil, from October 2016 to February 2018, at the coordinates 17°45′48″S and 51°02′14″W and at an altitude of 832 m. The soil was classified as a typical Dystrophic Red Latosol according to the Brazilian classification system (Santos et al., Reference Santos, Jacomine, Anjos, Oliveira, Lumbreras, Coelho, Almeida, Araújo Filho, Oliveira and Cunha2018).
The experiment was established in 2016. Soybean was always grown during the summer season, with the management systems subsequently evaluated during the off-season. During the experiment, the monthly rainfall and average temperature data were recorded (Fig. 1).
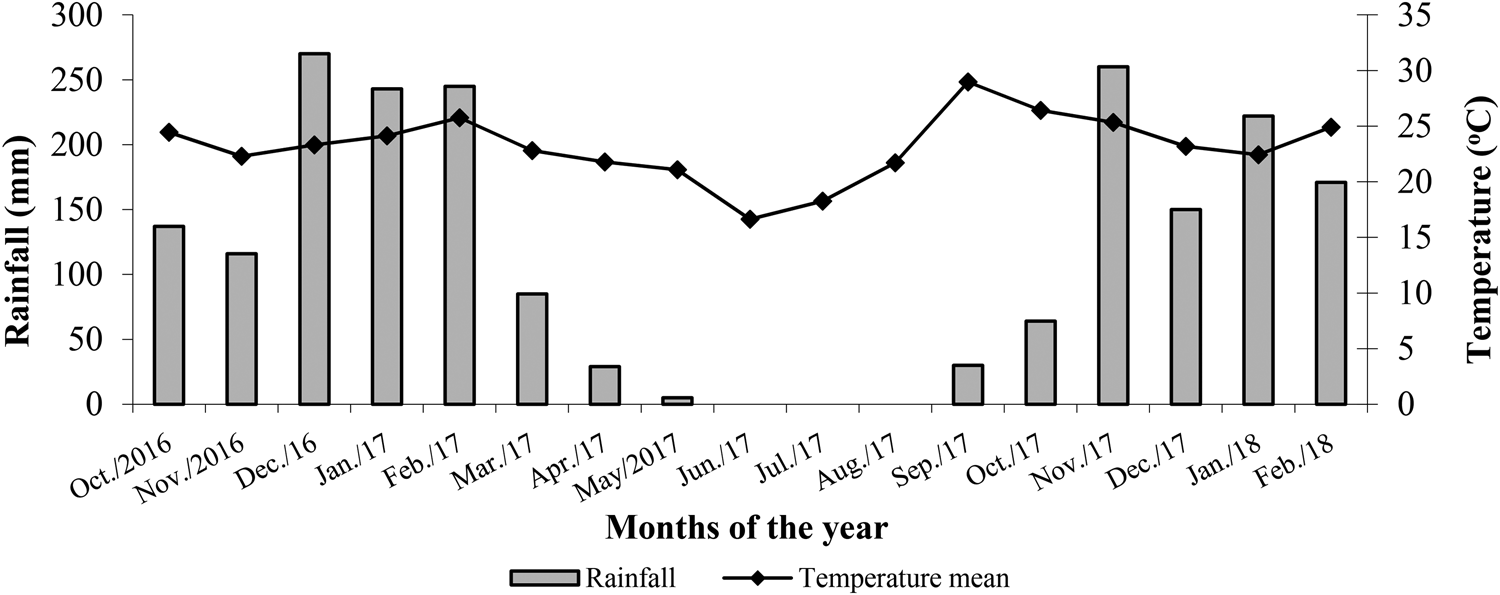
Fig. 1. Monthly rainfall and mean daily temperatures recorded from October 2016 to February 2018 in Rio Verde-GO, Brazil.
Treatments and crop establishment
The experimental design was a randomized block design with four replicates. The treatments consisted of the following cropping systems: Xaraes palisadegrass (Brachiaria brizantha cv. Xaraés) intercropped with soybean, Congo grass (B. ruziziensis) intercropped with soybean, Mombaça guinea grass (P. maximum cv. Mombaça) intercropped with soybean, Tamani guinea grass (P. maximum cv. BRS Tamani) intercropped with soybean and a soybean/maize (Z. mays) succession. The treatments were randomly distributed throughout the total area. The area of each plot was 2000 m2, and each plot was divided by an electric fence, such that there were four plots, each with a separate forage grass, and four plots containing maize.
Figure 2 shows a diagram of the cropping systems comprising Brachiaria spp. and Panicum forage grasses in an integrated crop–livestock system and a system with maize in succession to soybean. This system was designed to diversify activities, maximizing land use throughout the year.
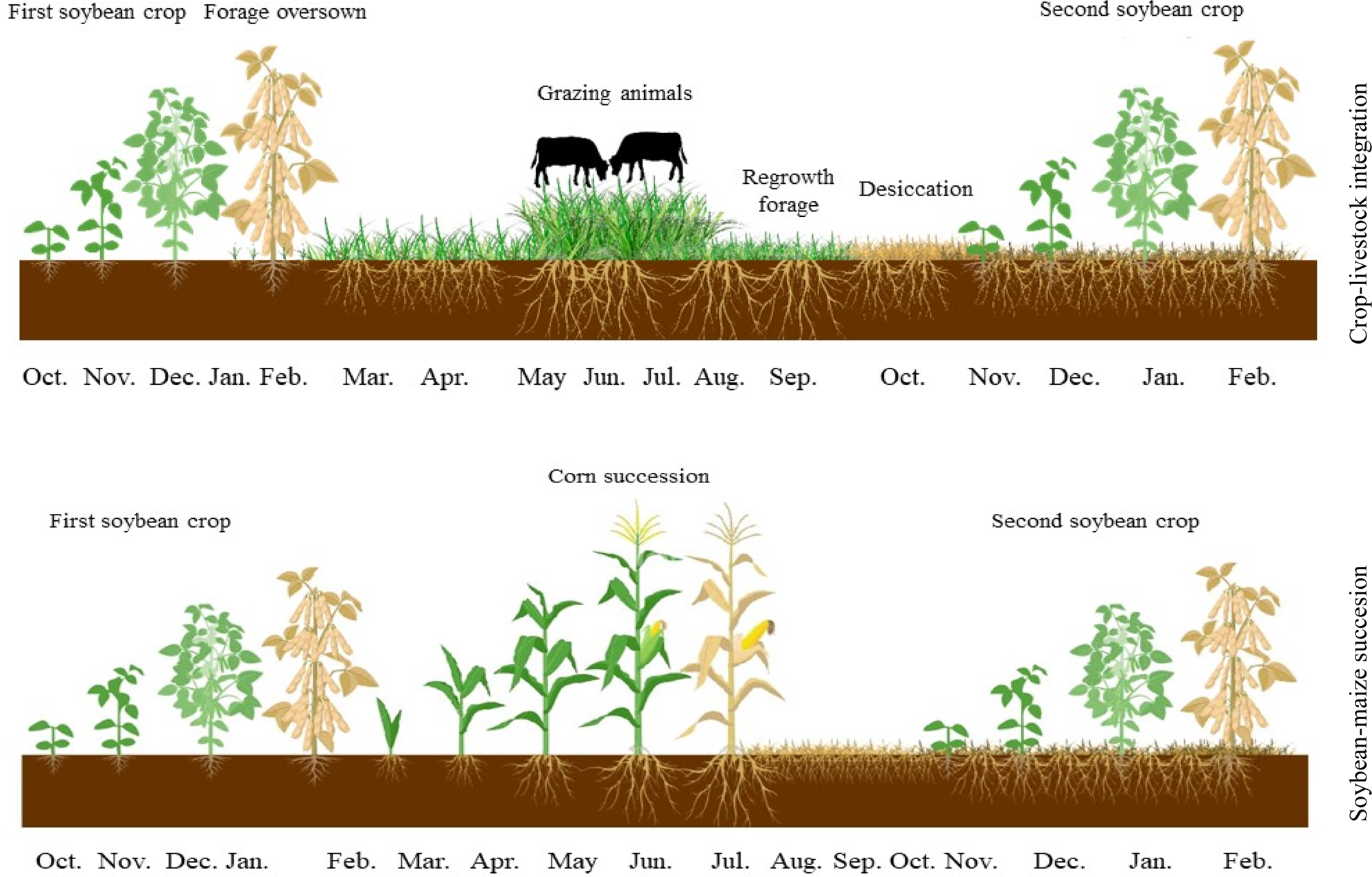
Fig. 2. (Colour online) Diagram of the cropping systems comprising Brachiaria spp. and Panicum forage species in an integrated crop–livestock system and a system with maize in succession to soybean, covering all evaluation stages.
In October 2016, the first soybean crop was established, and in February 2017, when the soybean plants were at the R6–R7 stage (Fehr and Caviness, Reference Fehr and Caviness1977) and the leaves began to become yellow and fall, the forage grass seed was sown. In total, 5.0 kg/ha of seed of each Brachiaria species and 3.5 kg/ha of seed of P. maximum were sown, which were rated as 60 and 40% of the seed germination and purity, respectively. The seed and monoammonium phosphate (MAP) were mixed together at a dose of 100 kg/ha, after which the mixtures were distributed over their respective areas via a Lancer spreader.
The soybean plants were harvested 34 days after the forage grass seed was sown, and the forage grasses continued to grow until the animals entered the system.
AG7098 maize was sown on 20 February 2017, and 300 kg/ha of 2-20-18 fertilizer was applied at sowing. At the V4–V6 phenological stage, topdressing urea fertilizer was applied at a dose of 90 kg/ha of nitrogen, according Spackman et al. (Reference Spackman, Fernandez, Coulter, Kaiser and Paiao2019). After weeds had emerged, atrazine, tembotrione and soybean oil methyl ester were applied at doses of 1500, 101 and 720 g of a.i./ha, respectively. At the same time, 0.3 litres/ha of Match (equivalent to 15 g/ha of lufenuron) and 1 litres/ha of Lanate (equivalent to 215 g/ha of methomyl) were also applied.
In April, animals were added to the system; 25 male Nellore cattle with an average weight of 232 kg were used. Each forage system received animals. The stocking rate varied and was adjusted as necessary throughout the experiment according to the forage availability shown in Table 1. The mean stocking rates were 2.66, 2.42, 3.20 and 2.52 animals/ha for Xaraes palisadegrass, Congo grass, Mombaça guinea grass and Tamani guinea grass, respectively. The animals remained in the plots for 141 days (off-season) as part of an intermittent grazing system, with the animals in the paddock for 7 days followed by a 28-day resting period for the forage, with a total of five grazing cycles.
Table 1. Dry matter production (kg/ha) of forage in different grazing cycles
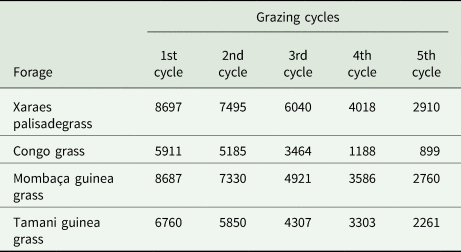
1st cycle (May), 2nd cycle (June), 3rd cycle (July), 4th cycle (August) and 5th cycle (September).
After the off-season (September 2017), the animals were removed from the area, and the forage grasses were allowed to recover and regrow before termination to form mulch for the no-till system. The forage grasses were killed 14 days before soybean planting by an application of glyphosate herbicide at a dose of 960 g of a.i./ha.
Assessment of mulch biomass and total nutrient
One day before soybean sowing, biomass samples of the mulch were collected from a 1-m2 area randomly selected within each plot. The plant material was cut at ground level. The cut material was weighed, the samples were placed in a forced-air oven at 65 °C until they reached a constant weight, and the amounts were converted to dry weight (kg/ha).
Mulch decomposition was evaluated for dead forage material collected in 25 × 30 cm litter bags made of a 2-mm mesh (Thomas and Asakawa, Reference Thomas and Asakawa1993). For this, four bags containing biomass samples of each species in amounts proportional to the dry mass produced per hectare were placed in direct contact with the soil.
At 30, 60, 90 and 120 days after the bags were placed on the soil, one litter bag was removed from each plot to evaluate the remaining amount of mulch biomass and to determine mulch decomposition throughout the 120-day period (which ended at soybean harvest). On the basis of the initial mass (kg/ha) in the systems, the loss percentage was calculated as the ratio of the mass of the litter bags (kg/ha) to the initial production.
At each evaluation time point, the material was subsequently sent to the laboratory. The soil debris was removed by washing the material with running water until all the debris was gone, after which the material was dried in an oven at 55 °C until it reached a constant weight, according to the methods of Costa et al. (Reference Costa, Costa, Severiano, Santos, Rocha, Souza, Brandstetter and Castro2017). The biomass samples were ground (2 mm) in a mill to determine the carbon (C), nitrogen (N), phosphorus (P), potassium (K), calcium (Ca), magnesium (Mg) and sulphur (S) contents according to the methods of Malavolta et al. (Reference Malavolta, Vitti and Oliveira1997). The C/N ratio of the material was subsequently calculated. To evaluate the nutrient contents, the macronutrient concentrations were multiplied by the dry mass production, and the results were expressed as kg/ha.
The N, P2O5 and K2O fertilizer equivalents of the mulch biomass of the forage and maize cropping systems were determined considering the atomic mass of the elements according to analytical chemistry conventions and the N, P and K concentrations of the biomass analysed (Santos et al., Reference Santos, Albuquerque Filho, Vilela, Ferreira, Carvalho and Viana2014).
Establishment of soybean in the 2017/18 cropping system and crop management practices
To determine the fertilizer recommendations for the second soybean crop (2017/18), soil samples were collected from the 0 to 20-cm layer, and their chemical properties were as follows: pH CaCl, 4.90; Ca, 2.58 cmolc/dm3; Mg, 0.74 cmolc/dm3; aluminium (Al), 0.11 cmolc/dm3; Al + hydrogen (H), 5.05 cmolc/dm3; K, 0.21 cmolc/dm3; cation exchange capacity, 8.59 cmolc/dm3; vanadium (V), 41.14%; P (Mehlich), 33.70 mg/dm3; copper (Cu), 1.23 mg/dm3; zinc (Zn), 6.33 mg/dm3; iron (Fe), 53.87 mg/dm3 and OM, 27.18 g/kg.
According to the soil test recommendations, before soybean was planted, 1 t/ha of limestone and gypsum was applied. In addition, MAP fertilizer was applied at planting at a rate of 250 kg/ha, which was equivalent to 25 and 125 kg/ha of nitrogen and P2O5 per row, respectively.
The soybean (variety M7110PRO) seed was sown on 11/07/2017 in rows spaced 0.50 m apart. As a seed treatment, 200 ml of the insecticide Standak Top (fipronil) was applied to each 100 kg of seed. The insecticide was applied by spraying the sowing furrow with inoculant containing the bacteria Bradyrhizobium elkanii SEMIA 587 and SEMIA 5019 (Gelfix, 5 × 109 CFU/ml) at a dose of 0.6 litres of commercial product/ha and applying the micronutrients cobalt (Co) and molybdenum (Mo) (Nodulus Gold, 12.9 and 139 g a.i./l) at a dose of 0.2 litres of commercial product/ha. A spray volume of 60 litres/ha was used for these applications.
At 15 days after sowing (DAS), 80 kg/ha of K was applied by broadcasting, with K2O used as the source. In addition, fungicides were applied at 40 DAS [0.3 litres/ha of the commercial product Orkestra (167 and 333 g of a.i./l pyraclostrobin and fluxapyroxad) and 0.5 litres/ha of Assist mineral oil], at 60 DAS [0.2 kg/ha of Elatus (300 and 150 g a.i./l azoxystrobin and benzovindiflupyr) and 0.6 litres/ha of Nimbus mineral oil), at 75 DAS [0.4 litres/ha of Fox (150 and 175 g of a.i./l fluoxastrobin and prothioconazole) and 0.3 litres/ha of Aureo vegetable oil) and at 85 DAS (0.3 litres/ha of Orkestra and 0.5 litres/ha of Assist).
Establishment of soybean stands
The sowing uniformity was evaluated 30 days after emergence with the aid of tape measures in nine central 2-m rows (spacing between the plants in according to recommended). During the evaluation, the following spacing patterns were used according to the Brazilian National Standards Organization (1994): double spacing, which is 0.5 times smaller than the average established spacing of reference (Xref); acceptable or normal spacing, which is 0.5–1.5 times the average of Xref; and flawed spacing, which 1.5 times greater than the average of Xref (Mantovani et al., Reference Mantovani, Cruz and Oliveira2015). The average spacing established for the population was 0.057 m between plants.
Evaluation of soybean agronomic traits
The soybean agronomic traits were evaluated at 102 DAS. The following traits were evaluated: the plant height and height of insertion of the first and last pods (measured as the distance between the soil surface and the apex and between the soil surface and the insertion of the first and last pod on the main stem, respectively); the number of pods per plant (the sum of all the pods with seeds of ten plants in the studied area of the plot, calculated as the mean number of pods per plant); the 1000-grain weight; and the yield (kg/ha).
Statistical design and analysis
To describe the decomposition of the mulch biomass and the nutrient contents, the data were fitted together with the standard error to an exponential mathematical model (y = aekx) and, for the C/N ratio, to a linear model (y = a + bx) via Sigma Plot software. The estimated equations were compared according to a procedure described by Snedecor and Cochran (Reference Snedecor and Cochran1989); this method tests the homogeneity of the data (F) and the significance of the slope (0.4343k) and intercept (log a) of a linear equation (log y = log a + 0.4343kx).
To calculate the half-life (t 1/2), i.e. the time required for 50% of the remaining biomass to be decomposed, the equation proposed by Paul and Clark (Reference Paul and Clark1989) was used, namely, t 1/2 = 0.693/k, where t 1/2 is the half-life of the dry biomass and k is a constant of the dry biomass decomposition.
The data concerning the total nutrient, fertilizer equivalents, soybean agronomic traits and grain yield were analysed by analysis of variance by the use of R software version 3.1.1 (2014); specifically, the ExpDes package was used (Ferreira et al., Reference Ferreira, Cavalcanti and Nogueira2014). The means were compared according to Tukey's test at a significance level of 5%.
Results
Aspects of Mulch biomass and nutrient cycling
The biomass production was affected (P < 0.05) by cropping system (Table 2). The greatest production at time zero (soybean sowing) occurred for Xaraes palisadegrass, followed by the Tamani guinea grass, Mombaça guinea grass and Congo grass; these results contrast with those of maize, which produced the least amount of biomass. This same trend was observed for the remaining biomass until the final cycle of soybean development (Fig. 3a), with the results of all the systems evaluated differing from each other. Tamani guinea grass and Xaraes palisadegrass had the shortest half-life, at 72 days and 92 days, respectively.

Fig. 3. Remaining mulch biomass (a) and C/N ratio (b) of forages for the Brachiaria and Panicum genera and maize during soybean cultivation (0–120 days).
Table 2. Biomass production and total nutrients in biomass of different cropping systems
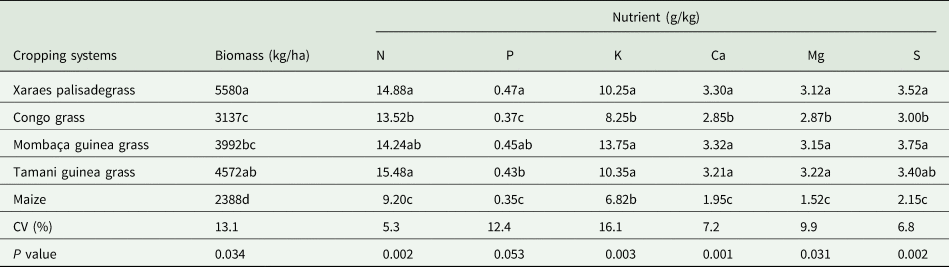
Averages followed by different letters do not differ from each other by Tukey's test at 5% probability.
There was a linear decrease in the C/N ratio for all the cropping systems over time, and the results of each system differed from each other according to comparisons of regressions following linearization (Snedecor and Cochran, Reference Snedecor and Cochran1989). Compared with the grasses, maize had a greater C/N ratio during all the evaluated seasons, and the C/N ratio of Congo grass was the second greatest (Fig. 3b). Tamani guinea grass had the lowest C/N ratio, with an initial value of 36.69 and a final value of 28.66.
The forage grasses, especially Xaraes palisadegrass, Mombaça guinea grass and Tamani guinea grass, had the greatest total nutrients. Maize had the lowest nutrients, which differed significantly (P < 0.05) from those of the forage grasses (Table 2).
There was a significant effect (P < 0.05) of cropping system on nutrient contents (Fig. 4), with an exponential reduction in the biomass content. Comparisons between nutrient-release equations indicated homogeneity only between Mombaça guinea grass and Tamani guinea grass for phosphorus and sulphur, according to the methods of Snedecor and Cochran (Reference Snedecor and Cochran1989). For this reason, these results were combined, and a new regression equation was generated.
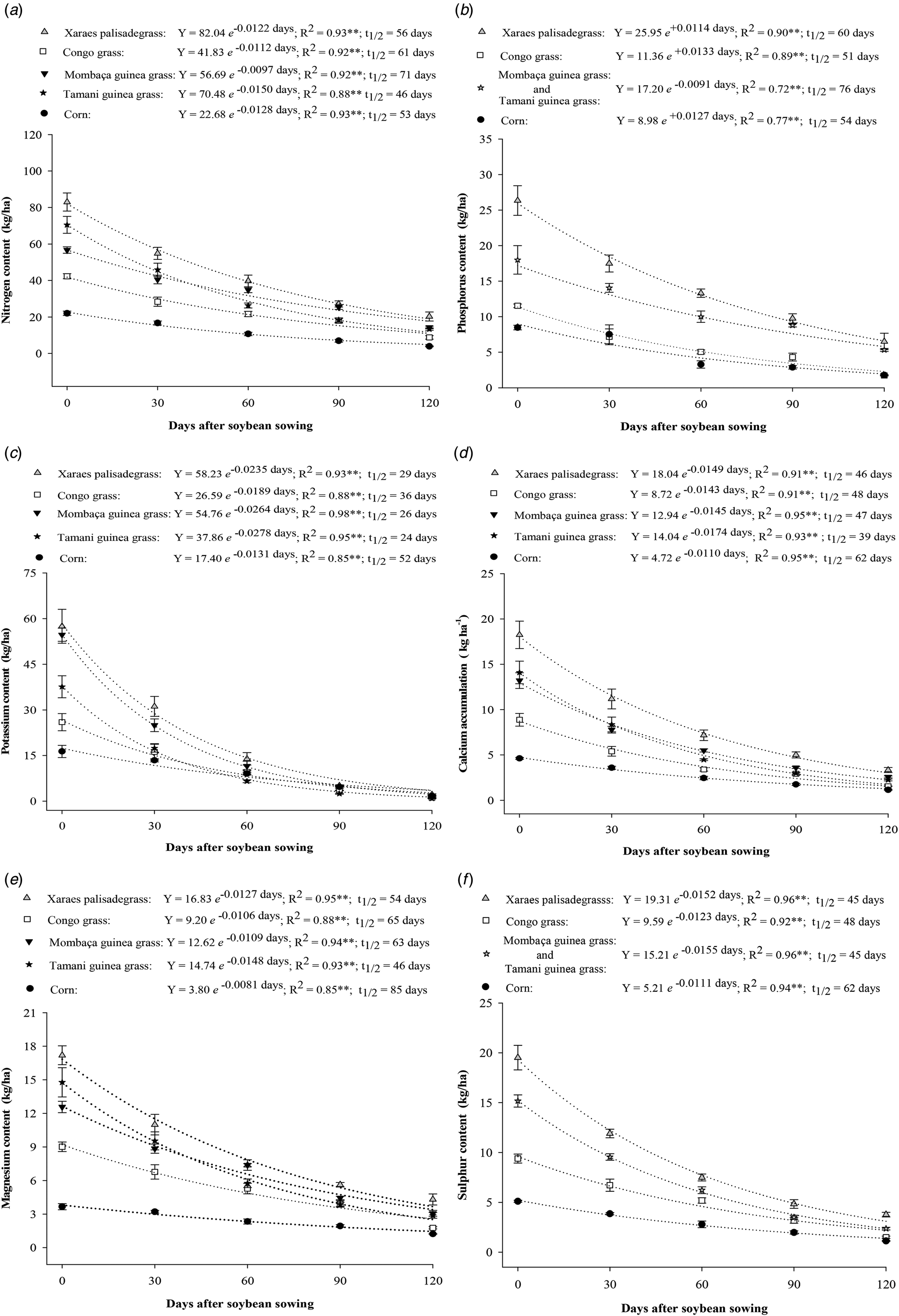
Fig. 4. Nitrogen (a), phosphorus (b), potassium (c), calcium (d), magnesium (e) and sulphur (f) content in the biomass of forages in the Brachiaria and Panicum genera and maize during soybean cultivation (0–120 days).
At time point zero and after 30 days of decomposition, the Xaraes palisadegrass and Tamani guinea grass had the greatest values of N content, and the maize presented the lowest N content (Fig. 4a). When the initial N content was compared with the values attained during the last evaluation at 120 days, 75.5, 79.3, 80.8, 74.6 and 82.4% N release occurred for the Xaraes palisadegrass, Congo grass, Tamani guinea grass, Mombaça guinea grass and maize, respectively.
With respect to the phosphorus content (Fig. 4b), maize had the lowest content, and Xaraes palisadegrass had the greatest content. When the initial phosphorus values were compared with the values attained at 120 days, 73.7, 84.9, 67.35 and 78.7% phosphorus release levels occurred for the Xaraes palisadegrass, Congo grass, Mombaça guinea grass, Tamani guinea grass and maize, respectively, indicating that, of these grasses, Tamani guinea grass released the most phosphorus.
Compared with the other grasses, Xaraes palisadegrass and Mombaça guinea grass had greater initial potassium contents (Fig. 4c). At 90 and 120 days, the values were similar between the cropping systems. The potassium release rates were 96.1, 95.5, 91.2, 97.6 and 88.7% for Xaraes palisadegrass, Congo grass, Mombaça guinea grass, Tamani guinea grass and maize, respectively.
For calcium (Fig. 4d), magnesium (Fig. 4e) and sulphur (Fig. 4f), the greatest initial contents were detected in Xaraes palisadegrass, followed by Tamani guinea grass and Mombaça guinea grass, whereas Congo grass and maize displayed the lowest potential to accumulate these nutrients in the soil. For Xaraes palisadegrass, Congo grass, Mombaça guinea grass, Tamani guinea grass and maize, the respective calcium release rates at 120 days were 81.2, 82.1, 80.1, 83.4 and 75%; for magnesium 74.4, 80.2, 75.7, 79.6 and 65.8% and for sulphur 80.4, 84.1, 83.7, 84.2% and 77.8%.
Potassium, calcium, magnesium and sulphur showed longer half-life within the maize biomass, while nitrogen and phosphorus had longer half-life within the Mombaça guinea grass biomass (Figs 4). However, the Tamani guinea grass had the shortest half-life in terms of the release of potassium, calcium and magnesium the half-life of sulphur was similar to that of Mombaça guinea grass; however, the shortest half-life for phosphorus occurred in Congo grass.
With respect to the evaluation of the fertilizer equivalents, Table 3 shows that, compared with the other grasses, Xaraes palisadegrass and Tamani guinea grass provided greater amounts of nitrogen equivalents. The same occurred for phosphorus, in which Xaraes palisadegrass provided the greatest amounts of phosphorus equivalents. With respect to potassium, Xaraes palisadegrass and Mombaça guinea grass showed the best results.
Table 3. N, P2O5 and K2O equivalent contents in the biomass of different cropping systems
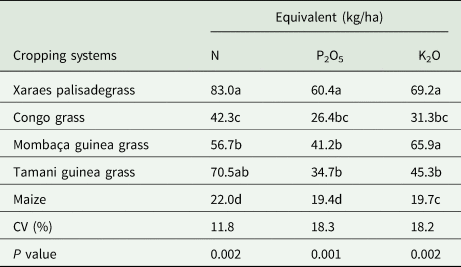
Averages followed by different letters do not differ from each other by Tukey's test at 5% probability.
Soybean population arrangement, agronomic traits and yield
Regarding sowing uniformity, there was no effect of cropping system on plant distribution or population (Fig. 5), which exhibited similar patterns.
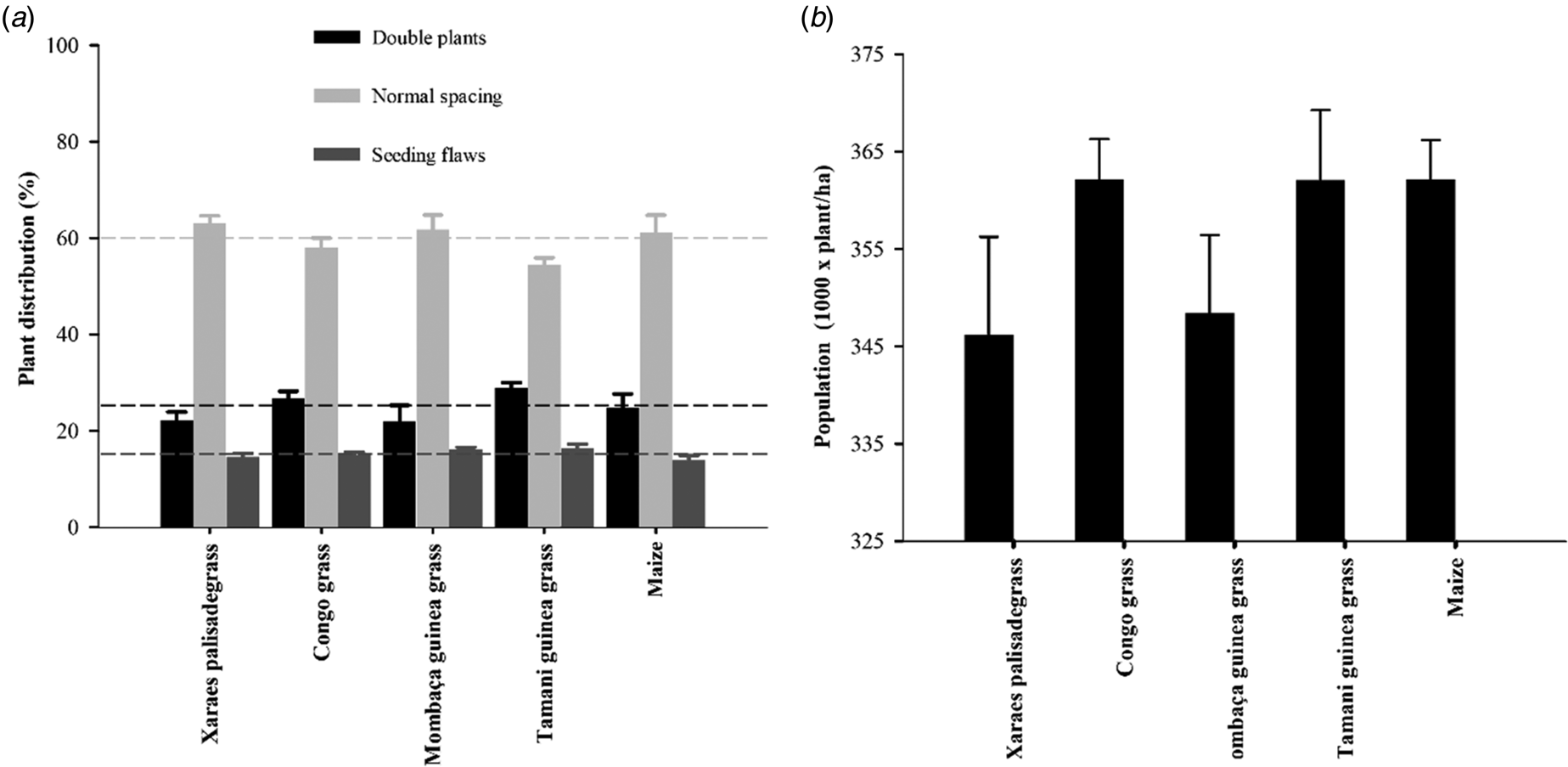
Fig. 5. Soybean sowing uniformity at the time of stand establishment in the different cropping systems plant distribution (a) and plant population (b) on a typical Dystrophic Red Latosol. The dashed lines indicate the mean, and the vertical bars represent the standard error of the mean.
There was a significant effect (P < 0.05) of cropping system biomass on soybean agronomic traits and grain yield (Table 4). The greatest plant height, height of the insertion of the first pod and number of pods per soybean plant occurred for cropping systems containing Xaraes palisadegrass, Mombaça guinea grass and Tamani guinea grass, while those with Congo grass and maize presented the lowest values.
Table 4. Plant and insertion of the first pod height, number of pods per plant and number of grains per pods, 1000-grain weight and soybean yield on the biomass of different cropping systems
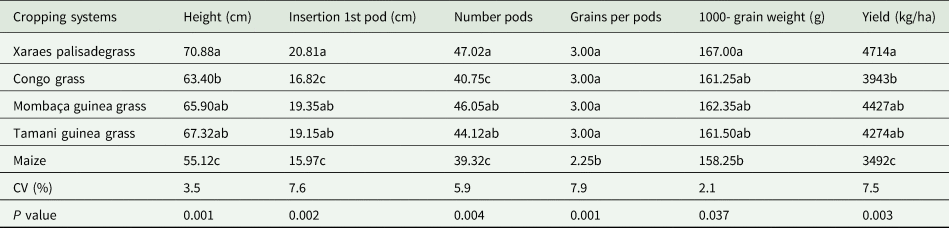
Averages followed by different letters do not differ from each other by Tukey's test at 5% probability.
With respect to the number of seeds per pod (Table 3), only the results of the maize cropping system significantly differed (P < 0.05) from among all the tested grasses; maize presented the lowest value, which equated to a 33% reduction in the number of seeds per pod compared with that of the forage grasses. The results of the 1000-grain weight were similar between the cropping systems, with Xaraes grass presenting the greatest value and differing (P < 0.05) only from maize. The lowest grain yield occurred in the maize cropping system. However, among the forage cropping systems, there was a significant difference only between Xaraes grass and Congo grass, the latter of which presented the lowest soybean yield (Table 3).
Discussion
Aspects of Mulch biomass and nutrient cycling
The greatest biomass production obtained by Xaraes palisadegrass followed by Tamani guinea grass is due to the morphology of these grasses; compared with the other grasses studied, these grasses regrow faster after animal grazing and can grow even under low-rainfall conditions (Fig. 3a). Xaraes palisadegrass is tall, and its leaves are wide; this grass displays vigorous and rapid regrowth (Costa et al., Reference Costa, Severiano, Simon, Epifanio, Silva, Costa, Santos and Rodrigues2014). However, Tamani guinea grass is small in size, but it has short, thin stalks that facilitate rapid growth, resulting in high yields (Machado et al., Reference Machado, Cecato, Comunello, Concenço and Ceccon2017).
Notably, compared with those of the other grasses, the stocking rates of Xaraes palisadegrass and Tamani guinea grass varied less at 141 days of grazing, maintaining greater stability during the grazing periods because of the improved development of these forage grasses (Table 1), even under low-rainfall conditions (Fig. 1). After the removal of the animals, these forage grasses displayed quick regrowth at the beginning of the rainy season, favouring the production of biomass at time zero (Table 2) for the subsequent crop in the soybean no-till systems.
These results are important for selecting the correct forage grass for an integrated crop–livestock system, with the potential to improve animal performance during the off-season and subsequently improve the biomass production of the subsequent crop. For many years, B. ruziziensis was considered one of the primary forage grass species for use in integrated crop–livestock systems for biomass production; however, new cultivars have emerged that have shown better potential, as reported in studies by Flávio Neto et al. (Reference Flávio Neto, Severiano, Costa, Guimarães Junnyor, Gonçalves and Andrade2015) on Xaraes palisadegrass and by Machado et al. (Reference Machado, Cecato, Comunello, Concenço and Ceccon2017) on Tamani guinea grass.
The relatively short half-life of Tamani guinea grass and Xaraes palisadegrass is related to the relatively high amount of leaves; this is due to the better regrowth of these grasses following grazing in September, consequently accelerating the biomass decomposition rate.
One of the key requirements for the success of no-till systems is the amount of biomass on the soil surface. Soil OM improves soil properties and is a source of both plant nutrients and energy for microorganisms (Malhi et al., Reference Malhi, Légère, Vanasse and Parent2018). The quantity and quality of soil organic C and N are affected by soil and crop management practices (Liu et al., Reference Liu, Hebert, Hashemi, Zhang and Ding2006). Thus, the correct choice of plant species is extremely important because the climatic factors that are characteristic of each region and soil type must be considered (Costa et al., Reference Costa, Andreotti, Ulian, Costa, Pariz and Teixeira Filho2015).
Given this information, dry spells are notably frequent in the Brazilian Cerrado, even during the rainy season, hindering crop development. Additionally, the amount of biomass produced by Xaraes palisadegrass, Tamani guinea grass and Mombaça guinea grass (2375, 1619 and 1858 kg/ha, respectively) was advantageous; compared with Congo grass and maize, whose biomass amounts were 1233 and 1070 kg/ha at the end of the soybean development cycle, these grasses displayed greater productivity and were distributed in greater amounts in the soil.
The relatively low amount of maize biomass (Fig. 4a) is due to the early decomposition of maize during the off-season; compared with the forage grasses, which were killed 14 days before soybean was sown, maize remained in the soil for a longer duration. Moreover, maize biomass does not thoroughly cover the soil despite the large number of stalks; rather, maize biomass leaves gaps in the cover and thus leaves the soil unprotected.
The decomposition rate of the remaining biomass determines the residence time of the mulch on the soil, which is influenced by the lignin contents and can be reflected by the C/N ratio. Forage grasses generally produce relatively high amounts of biomass, which is also characterized by a high C/N ratio, contributing in turn to increased longevity of the soil cover (Oliveira et al., Reference Oliveira, Almeida, Junior, Reis, Souza and Favarin2019). C/N ratios lower of 20 contribute to mineralization, and greater values contribute to the immobilization of nutrients in the soil; the ideal is a balance between the two processes – C/N ratios between 20 and 30 (Truong and Marschner, Reference Truong and Marschner2018).
In this context, until the final cycle of soybean development, the Brachiaria spp. and Panicum forage grasses notably maintained a C/N ratio greater than 30, which was closer to the ratio associated with immobilization than to that associated with mineralization. Therefore, the C/N ratios of the forage grasses were important for determining the immobilization and the decomposition rate of the mulch biomass. These results are extremely important in terms of the central region of Brazil, which is usually characterized by high temperatures, favouring the decomposition of mulch biomass.
The relatively high C/N ratio obtained for the maize biomass (Fig. 3b) is due to the relatively high amount of lignified stalks of maize crops; this lignified tissue is relatively resistant to the action and penetration of decomposing microorganisms, which hinders its decomposition. The greatest concentrations of carbon in maize plants are usually found in the ears and stems, followed by the leaves, tassels and husks; in practical terms, the high C/N ratio of the biomass ensures the persistence of the soil cover for an extended period of time (Barbosa et al., Reference Barbosa, Ferreira, Santos, Motta, Prior and Gabardo2016).
The greater nitrogen content in Xaraes palisadegrass and Tamani guinea grass compared with the other forage grasses (Fig. 4a) may be attributed to their greater biomass. Furthermore, these results may be related to the increased production of leaves by these grasses, which are the organs in which excess nutrients are concentrated, thus favouring N cycling. Another factor that may have an effect was the relatively stable stocking rate during the 141-day grazing period, which resulted in an improved nutrient return through manure and urine excretion, increasing nutrient recycling and thus resulting in increased concentrations (Table 2).
A study by Costa et al. (Reference Costa, Andreotti, Ulian, Costa, Pariz and Teixeira Filho2015) on the nutrient contents and decomposition rate of the biomass of forage plants showed that Xaraes palisadegrass accumulated 81.3 kg/ha of N; which this amount was similar to that obtained in the current study, which was 83.0 kg/ha at time point zero. Soratto et al. (Reference Soratto, Costa, Crusciol, Neto Ferrari and Moro2019) evaluated the nitrogen content of Mombaça guinea grass and reported amounts that surpassed 95.3 kg/ha.
Compared with other nutrients, nitrogen and potassium are extracted in greater amounts by forage plants used for biomass production; thus, these nutrients are presents in relatively a greater proportion within the biomass. These two nutrients are also the most easily leached, especially in deep soils, thus hindering access to them by the root system of many crops; this is an advantage for grasses with deep and aggressive root systems and is a benefit to soybean as a subsequent crop species, given that there is a relatively high concentration of these nutrients in the soil (Table 2).
The greatest content of phosphorus occurring in Xaraes palisadegrass is also due to the relatively high biomass production and, consequently, increased nutrient cycling. According to Jouany et al. (Reference Jouany, Cruz, Daufresne, Duru, Bunemann, Oberson and Frossard2011), grazing can be considered a regulator of labile phosphorus, and the effects of this process depend on pasture consumption and urine and manure recycling.
In general, tropical forage species are extremely important in production systems because they recycle nutrients from the subsoil, replenish OM and promote soil decompaction because of the abundant volume and aggressiveness of their root systems and the resulting biological activity. This phenomenon was demonstrated in a study by Flávio Neto et al. (Reference Flávio Neto, Severiano, Costa, Guimarães Junnyor, Gonçalves and Andrade2015), who reported that Xaraes palisadegrass had a high potential for soil decompaction, which is essential in areas that have been under no tillage for many years because it increases the water availability of crops in succession and promotes soil conservation in the Cerrado region.
The greatest potassium content occurring in Xaraes palisadegrass and Mombaça guinea grass is also due to their relatively high biomass production, as these forage grasses are tall and produce both a large amount of leaves and an abundance of roots. Most of the potassium accumulated within the biomass remains in the shoots before being released into the soil and used by the subsequent crop (Miguel et al., Reference Miguel, Pacheco, Carvalho, Souza, Feitosa and Petter2018).
The release of potassium is very interesting because the potassium suddenly decreased immediately after the grasses were killed; the lowest half-life were 29, 26 and 24 days for Xaraes palisadegrass, Mombaça guinea grass and Tamani guinea grass, respectively. However, Congo grass and maize had relatively long half-life – 36 and 52 days. Thus, notably, the nutrient release rate during decomposition depends on the material and the form of these nutrients in the plant tissues.
In the current study, the amount of potassium released at 30 days reached 45.1, 38.5, 53.9, 52.1 and 53.9% for Xaraes palisadegrass, Congo grass, Mombaça guinea grass, Tamani guinea grass and maize, respectively. The amount that remained within the biomass during the final cycle of soybean development was very small; the average was 6.18% of the total unreleased potassium from all the cropping systems. Taken together, these results show that the biomass presented release rates that exceeded 90% and demonstrate the representativeness of this the return of this nutrient to the soil via the biomass.
When evaluating the release rate of nitrogen, phosphorus and potassium within the mulch biomass of Marandu palisadegrass, Congo grass, Mombaça guinea grass, Tanzania guinea grass and Aries guinea grass, Pereira et al. (Reference Pereira, Mello, Pariz, Mendonça, Yano, Miranda and Crusciol2016) reported that the maximum release of these nutrients occurred during the first 30 days. In general, at 120 days after planting, more than 60% of the nutrients had already been released from the mulch, and these results are similar to those found in the current study.
The presence of the relatively low calcium and magnesium contents in maize is due to the early decomposition of the maize biomass until the soybean crop was sown, which is a disadvantage of this cropping system compared with integrated crop–livestock systems because of the relatively low soil cover at the time of soybean sowing. According to Crusciol et al. (Reference Crusciol, Cottica, Lima, Andreotti, Moro and Marcon2005), calcium and magnesium are easily released because they are components of ionic compounds and soluble molecules in plants and are thus quickly released into the soil by the decomposition process.
Relatively long half-life of nutrients (in terms of their release) are due to relatively high lignin contents and C/N ratios of the plants. In the current study, compared with the forage grasses, maize had a longer half-life for potassium, calcium, magnesium and sulphur. The stalk formation process of this species also contributes to the relatively high lignification of tissues, hindering nutrient cycling (Barbosa et al., Reference Barbosa, Ferreira, Santos, Motta, Prior and Gabardo2016).
The shorter half-life of the release of most nutrients and the lower C/N ratio of the Tamani forage grass compared with the other grasses is due to the high proportion of leaves and to its very fine and less lignified stems. Its biomass production, which reached 1619 kg/ha, allows protection of the soil until the end of the soybean development cycle and rapid nutrient cycling.
The lowest nutrient content values were generally recorded in the Congo grass and maize. Maize in succession with soybean is not the best option for biomass formation and nutrient cycling because its soil cover is inefficient and because its stalks are highly lignified, necessitating more time for nutrient release, with long-term benefits.
Moreover, Congo grass, which is one of the most frequently used forage grasses for this technique, did not perform satisfactorily in this study, indicating that new cultivars such as B. brizantha cv. Xaraes and Panicum cultivars (Mombaça and Tamani) might be better options for obtaining mutual benefits in integrated crop–livestock systems and no-till systems, as these new cultivars produce more biomass and, consequently, greater nutrient contents, increasing the efficiency of these systems.
The order of nutrient contents in the remaining biomass in all the cropping systems studied was as follows: N > K > P > Ca > S > Mg. These results highlight the importance of biomass production for soybean no-till systems because these systems involve nutrient cycling. In addition, this production maintains the C stocks in the biomass and in the soil and is considered a greenhouse-mitigating and environmentally friendly soil management practice (Costa et al., Reference Costa, Costa, Severiano, Santos, Rocha, Souza, Brandstetter and Castro2017).
The greater amounts of nitrogen, phosphorus and potassium equivalents obtained via Xaraes palisadegrass compared with the other grasses (Table 3) are due to the greater biomass production and subsequent release of nutrients. Knowledge of fertilizer equivalents is highly important with respect to fertilizer programmes because the amount released must be considered for fertilizer recommendations for the subsequent crop (Santos et al., Reference Santos, Albuquerque Filho, Vilela, Ferreira, Carvalho and Viana2014; Assmann et al., Reference Assmann, Martins, Anghinoni, Denardin, Nichel, Costa, Silva, Balerini, Carvalho and Franzluebbers2017).
Thus, when considering the high cost of fertilizer, it is possible that the continuous use of cover crops in soybean agricultural systems enables the most efficient use of the nutrients available in the soil, with favourable effects on the cost of production, increasing production sustainability.
Soybean population arrangement, agronomic traits and yields
The sowing uniformity revealed positive characteristics for soybean in the mulch biomass in all the cropping systems, with less than 20% of flawed spacing in all the systems. When evaluating soybean, Zardo and Casimiro (Reference Zardo and Casimiro2016) reported 25.4% of flawed and 23.3% of double spacing, and these percentages were similar to those in the current study. Some authors have noted that the arrangement of soybean plants affects their productivity and is related to the adaptability of plants to the available space (Carmo et al., Reference Carmo, Braz, Simon, Silva and Rocha2018).
Notably, the period during which crops in integrated systems are killed may affect the sowing uniformity and agronomic performance of soybean in succession (Franchini et al., Reference Franchini, Balbinot Junior, Debiasi and Conte2015) because of the moisture of the biomass present on the soil at the time of planting, which may increase the risk of wheel slip by machinery during sowing and contribute to clogging of the machine with uncut biomass that in turn accumulates in the path of the seeder (Aratani et al., Reference Aratani, Freddi, Centurion and Andrioli2009).
Nevertheless, soybean farmers are resistant to cultivating tall grasses that display cespitose growth, such as Xaraes palisadegrass and Mombaça guinea grass, for the reasons presented here. On the basis of the results, the period between crop kill and soybean sowing was sufficient for the generation of biomass of all the evaluated forage grasses, highlighting the importance of animal management during the off-season for the control of large numbers of clumps of tall forage grasses.
The greater plant height, height of insertion of the first pod and number of pods per plant obtained for the Xaraes palisadegrass, Mombaça guinea grass and Tamani guinea grass than for Congo grass and maize are associated with the greater biomass of those forage grasses, which may have contributed to their greater release of nutrients, given that these forage grasses had greater total nutrient in their biomass and given that nutrient return in those systems favoured improved mineral nutrient contents for the subsequent soybean crop (Table 2).
Mulch biomass also provides physical benefits such as increased aeration and uncompacted soil (Flávio Neto et al., Reference Flávio Neto, Severiano, Costa, Guimarães Junnyor, Gonçalves and Andrade2015). Mulch biomass also provides biological benefits such as increased microbial decomposition activity and excretion of animal faeces and decreased amounts of weeds and pesticide applications, improving soybean plant establishment (Balbinot Junior et al., Reference Balbinot Junior, Franchini, Werner and Ferreira2016).
Compared with those cultivated on the other evaluated grasses, soybean plants cultivated on the Xaraes palisadegrass biomass presented greater insertion heights of the first pod because of their greater biomass production, leading to better crop development. Thus, the biomass in the integrated crop–livestock systems was important for increasing the height of pod insertion, which is directly related to grain yield (Carkner and Entz, Reference Carkner and Entz2017).
The lower values of the number of pods per plant found in the maize and Congo grass cropping systems compared with the other cropping systems can probably be attributed to the lower amount of soil cover and less nutrient cycling.
There is a positive relationship between the number of pods per plant and the other traits, in which increasing plant height and insertion height of the first and last pods results in an increasing number of pods per plant. It has also been reported that relatively large stems result in relatively large tillering areas (Lima et al., Reference Lima, Crusciol, Cavariani and Nakagawa2009). In the current study, the greatest values observed for all these traits were related to the relatively high forage biomass production and nutrient content and release, indicating the efficiency of this system for the production of adequate amounts of biomass for sequential crops in no-till systems.
Compared with the 33% reduction in the number of seeds per pod in the maize cropping system, the biomass produced by the forage grasses shows the importance of using new Brachiaria and Panicum cultivars in integrated crop–livestock systems to generate biomass for subsequent crops.
Similar to that which occurred for the other agronomic traits, seed production was greater in forage grass cropping systems than in the maize cropping system; the forage grass cropping systems produced more biomass because of the greater soil protection they provided for the soybean crop and because of the resulting nutrient cycling. It is important to emphasize that in the livestock phase, animal excreta are used, providing a pathway for the nutrients to return to the soil (He et al., Reference He, Pagliari and Waldrip2016), and grazing promotes the vigorous regrowth of forage grasses, favouring growth for the production of mulch biomass.
Several studies have shown that grazing livestock in an integrated crop–livestock system can alter nutrient and mulch cycling rates, changing the biochemical form of nutrients, the spatial distribution within the pasture and the time of release or availability (Assmann et al., Reference Assmann, Martins, Anghinoni, Denardin, Nichel, Costa, Silva, Balerini, Carvalho and Franzluebbers2017).
Thus, appropriate management of animals in these systems is important for maximizing the benefits of nutrient cycling, with increased soil OM (Assmann et al., Reference Assmann, Anghinoni, Martins, Costa, Kunrath, Bayer and Carvalho2014) occurring because of nutrient return through the excretion of manure and urine. The animals act as nutrient recycling agents (Carvalho et al., Reference Carvalho, Anghinoni, Moraes, Souza, Sulc, Lang, Flores, Lopes, Silva, Conte, Wesp, Levien, Fontaneli and Bayer2010b), which may result in a reduced need for fertilizer (Russelle et al., Reference Russelle, Entz and Franzluebbers2007) for the subsequent crop and reduce maintenance of production sustainability over time (Costa et al., Reference Costa, Costa, Santos, Severiano, Epifânio, Silva, Teixeira and Silva2016).
Thus, the main advantage of the use of animals in the off-season in integrated systems compared with maize cropping systems is due to the benefits provided by forage grasses and animals. The sowing of soybean onto forage biomass is an option for the diversification of activities (Fig. 2) and the maximization of land use throughout the year, which increases revenue per area (Crusciol et al., Reference Crusciol, Nascente, Mateus, Pariz, Martins and Borghi2014).
Conclusion
Compared with Congo grass and maize, Xaraes palisadegrass, Mombaça guinea grass and Tamani guinea grass produced greater biomass and fertilizer equivalents returned to the soil and caused greater nutrient cycling, showing the benefits of these integrated production systems.
Maize had a greater C/N ratio than did the forage grasses, but the latter also displayed high potential, protecting the soil until the end of the soybean development cycle. The use of a crop–livestock integration system with a forage crop provided greater soil nutrient cycling compared with that provided by the maize cropping system, which resulted in greater soybean yield, thus contributing to the agricultural sustainability.
Financial support
This work was supported by the Goias Federal Institute (Instituto Federal Goiano).
Conflict of interest
The authors declare there are no conflicts of interest.
Ethical standards
Not applicable.









