



1. Introduction
Located on the northeastern margin of the Qinghai-Xizang (Tibetan) Plateau, the Qaidam Basin is renowned for its extensively developed Mesozoic and Cenozoic terrestrial deposits with abundant hydrocarbon source rocks of Jurassic age (Zhang et al. Reference Zhang, Li, Xiong, Zhang, Wang, He, Lin and Sun1998; Ritts et al. Reference Ritts, Hanson, Zinniker and Moldowan1999). These deposits also represent the most complete Lower–Middle Jurassic plant-bearing sequences of Northern China (Li et al. Reference Li, He, Wu, Mei and Li1988; Zhou, 1995; Wang et al. Reference Wang, Mosbrugger and Zhang2005). Based on these plant fossils, a series of palaeobotanical studies have been carried out since the 1950s, demonstrating that highly diverse vegetation grew in this basin during the Early–Middle Jurassic, including bryophytes, ferns, cycads, bennettitaleans, ginkgos and conifers (e.g., Sze, Reference Sze1959; Li et al. Reference Li, He, Wu, Mei and Li1988; Wang et al. Reference Wang, Mosbrugger and Zhang2005; Zhang et al. Reference Zhang, Wang, Ruhl, Kovács, Xu, Zhu, Lu and Chen2024). Fossil wood is regarded as one of the most important types of plant fossils with informative significance for exploring the vegetation composition and palaeoclimate variations (Wang et al. Reference Wang, Tian, Jiang, Yang and Ding2017); however, detailed systematic studies of fossil wood material from the Qaidam Basin have been absent so far.
Since the beginning of the 20th Century, a number of fossil wood specimens have been described from Pennsylvanian–Cenozoic deposits in China (Zheng et al. Reference Zheng, Li, Zhang, Li, Wang, Yang, Yi, Yang and Fu2008), with the Jurassic and Lower Cretaceous fossil wood record being extraordinarily diversified (Zheng et al. Reference Zheng, Li, Zhang, Li, Wang, Yang, Yi, Yang and Fu2008; Wang et al. Reference Wang, Yang, Zhang, Zheng and Tian2009; Yang et al. Reference Yang, Wang and Zhang2013). Up to now, about 20 genera and 40 species of fossil wood have been described from the Jurassic of China (Zheng et al. Reference Zheng, Li, Zhang, Li, Wang, Yang, Yi, Yang and Fu2008; Wang et al. Reference Wang, Yang, Zhang, Zheng and Tian2009; Xie et al. Reference Xie, Wang, Tian and Uhl2023, Reference Xie, Wang, Tian, Xu, Jiang, Xie, Wang and Uhl2024). However, as mentioned above, so far little is known about fossil wood from the Qaidam Basin, although some sedimentological studies mentioned the occurrences of fossil wood (Ritts, Reference Ritts1998; Ritts et al. Reference Ritts, Hanson, Zinniker and Moldowan1999; Li et al. Reference Li, Shao, Liu, Lu, Spiro, Wen and Li2016; Lu et al. Reference Lu, Zhou, Yang and Shao2020), but without providing detailed systematic investigations.
During recent fieldwork in the Dachaidan area of Qinghai Province, a specimen of permineralized fossil wood was collected from the Lower Jurassic Huoshaoshan Formation. In the present study, this well-preserved fossil specimen is investigated in detail, to describe its anatomy and carry out taxonomic determination, to interpret the palaeoclimatic signals preserved in the tree growth rings of the wood, and to shed new light onto the palaeobiogeography of the Gondwana type of Metapodocarpoxylon wood. The fossil wood here is recognized as Metapodocarpoxylon libanoticum (Edwards) Dupéron-Laudoueneix et Pons, so far only known from the Jurassic and Cretaceous deposits in Northern Gondwana regions (e.g., Bamford et al. Reference Bamford, Roberts, Sissoko, Bouaré and O’leary2002; Philippe et al. Reference Philippe, Cuny, Bamford, Jaillard, Barale, Gomez, Ouaja, Thévenard, Thiébaut and Von Sengbusch2003; El-Noamani et al. Reference El-Noamani, El-Saadawi and Philippe2021), thus representing the first record of Metapodocarpoxylon in Laurasia.
2. Geological setting and stratigraphy
Located at the northeastern margin of the Qinghai-Xizang (Tibetan) Plateau, the Qaidam Basin covers an area of ∼120,000 km2 (36°–39° N, 91°–98° E). The basin is surrounded by the Altyn Mountains in the northwest, the Qilian Mountains in the northeast and the Kunlun Mountains in the south (Fang et al. Reference Fang, Zhang, Meng, Gao, Wang, King, Song, Dai and Miao2007; Fig. 1). During the Mesozoic, the Qaidam Basin was tectonically influenced by the evolution of the Meso-Tethys, Neo-Tethys and Mongol-Okhotsk Ocean, as well as the collisions of related blocks (Ritts & Biffi, Reference Ritts, Biffi, Hendrix and Davis2001; Kravchinsky et al. Reference Kravchinsky, Cogné, Harbert and Kuzmin2002; Kapp et al. Reference Kapp, Decelles, Gehrels, Heizler and Ding2007; Gehrels et al. Reference Gehrels, Kapp, Decelles, Pullen, Blakey, Weislogel, Ding, Guynn, Martin, Mcquarrie and Yin2011). The basement of the Qaidam Basin is pre-Paleozoic crystalline rocks and Paleozoic non-metamorphic or shallow metamorphic rocks (Wang et al. Reference Wang, Mosbrugger and Zhang2005; Gong et al. Reference Gong, Chen, Wang, Kusky, Wang, Zhang, Ba and Liao2012; Liu et al. Reference Liu, Zhang, Xue, Wu and Chen2014). After a series of tectonic evolutionary stages, the intermontane basin developed terrestrial deposition from the Jurassic onwards, consisting of Mesozoic-Cenozoic sediments in a fluvial-lacustrine depositional environment, with a thickness of 3–16 km (Ritts & Biffi, Reference Ritts, Biffi, Hendrix and Davis2001; Xia et al. Reference Xia, Zhang, Yuan, Fan and Zhang2001; Zhuang et al. Reference Zhuang, Hourigan, Ritts and Kent-Corson2011; Jian et al. Reference Jian, Guan, Zhang and Feng2013; Ren et al. Reference Ren, Chen, Kelsey, Gong and Liu2017; Zhou et al. Reference Zhou, Lu, Zhang, Yang, Gao, Shao and Hilton2022).
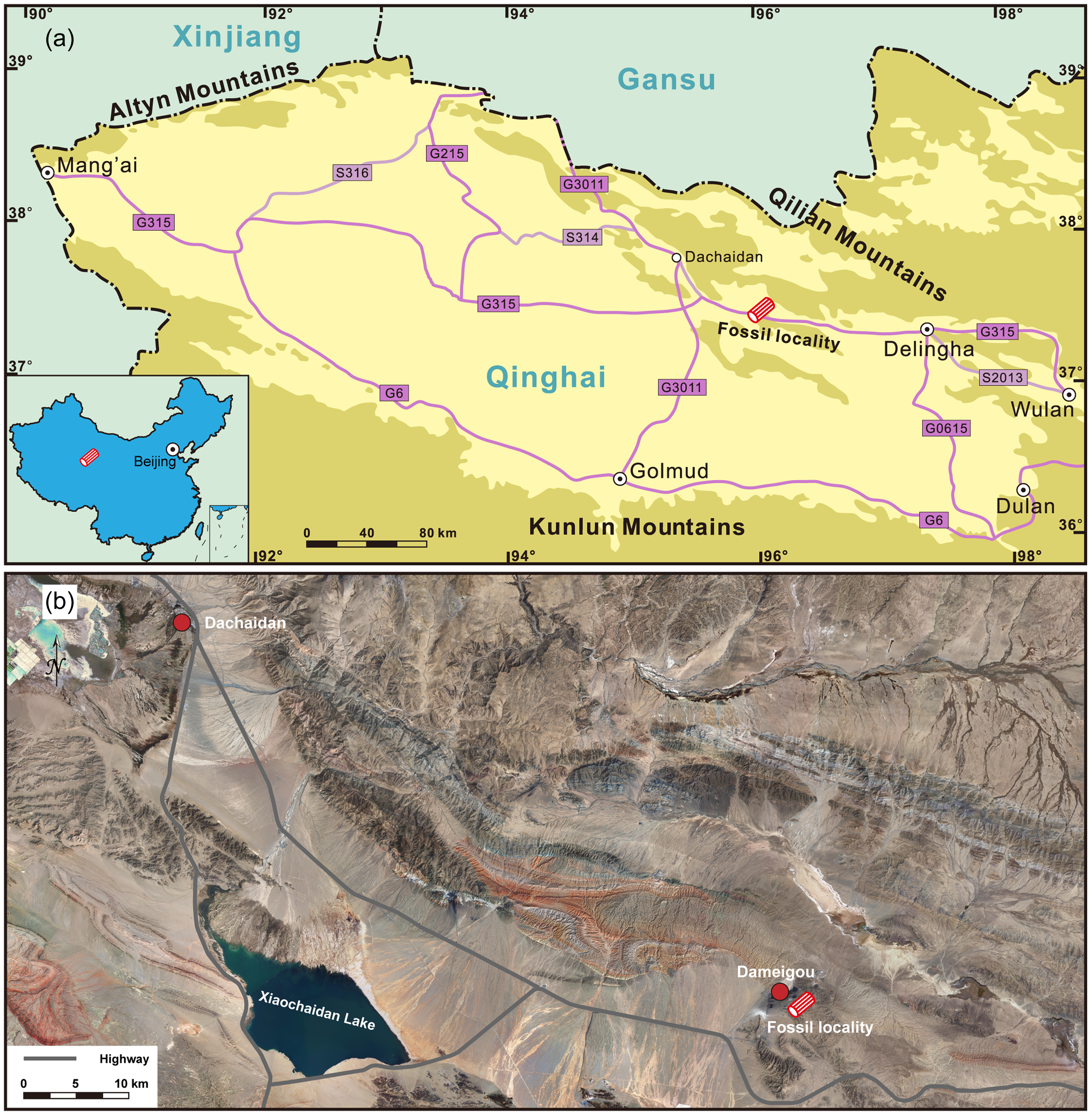
Figure 1. (a) Sketch map of fossil wood locality at Dameigou section in Dachaidan Town, Delingha City, Qinghai Province, China. (b) Close-up of (a), showing details of fossil wood locality; the base satellite image is according to the National Platform for Common GeoSpatial Information Services.
The Jurassic strata mainly crop out at the northern margin of the Qaidam Basin in the Yuqia, Dameigou, Baishushan, and Wanggaxiu areas (Li et al. Reference Li, He, Wu, Mei and Li1988; Zhang et al. Reference Zhang, Li, Xiong, Zhang, Wang, He, Lin and Sun1998; Wang et al. Reference Wang, Mosbrugger and Zhang2005). In these areas, the Jurassic deposits have been divided into seven formations: the Lower Jurassic Xiaomeigou Formation, Huoshaoshan Formation, Tianshuigou Formation and Yinmagou Formation, as well as the Middle Jurassic Dameigou Formation, Shimengou Formation and Caishiling Formation in ascending chronostratigraphic order (Zhang et al. Reference Zhang, Li, Xiong, Zhang, Wang, He, Lin and Sun1998; Wang et al. Reference Wang, Mosbrugger and Zhang2005; Zhou et al. Reference Zhou, Lu, Zhang, Yang, Gao, Shao and Hilton2022). Specifically, the Lower Jurassic is only well-exposed in the Dameigou section, representing a relatively complete Jurassic succession in the Qaidam Basin (Zhang et al. Reference Zhang, Li, Xiong, Zhang, Wang, He, Lin and Sun1998; Wang et al. Reference Wang, Mosbrugger and Zhang2005; Zhou et al. Reference Zhou, Lu, Zhang, Yang, Gao, Shao and Hilton2022).
The fossil wood specimen studied here was collected from the Huoshaoshan Formation at Dameigou section (37°30′43″ N, 96°2′17″ E) in Dachaidan Town, Delingha City, Qinghai Province, China. This formation is composed of conglomerates, sandstones, mudstones and coal seams, representing a braided delta plain depositional environment (Fig. 2) (Jin et al. Reference Jin, Qi, Xue, Wang and Fan2006; Yang et al. Reference Yang, Yang, Ma, Dong and Yuan2007; Yu et al. Reference Yu, Xiao, Wu, Tian, Rittner, Lou and Pan2017; Lu et al. Reference Lu, Zhou, Yang and Shao2020; Zhao et al. Reference Zhao, Zeng, Tian, Hu, Wang, Yan, Wang and Zhao2020). In general, the age of the Huoshaoshan Formation is regarded as Early Jurassic (Pliensbachian), based on evidence from lithology, chronostratigraphy (detrital zircon U-Pb dating and carbon isotope chemostratigraphy), cyclostratigraphy (based on cyclostratigraphcial analysis of rock colour datasets) and biostratigraphy (palaeobotany and palynology) (Zhang et al. Reference Zhang, Li, Xiong, Zhang, Wang, He, Lin and Sun1998; Wang et al. Reference Wang, Mosbrugger and Zhang2005, Yu et al. Reference Yu, Xiao, Wu, Tian, Rittner, Lou and Pan2017; Lu et al. Reference Lu, Zhou, Yang and Shao2020; Zhou et al. Reference Zhou, Wang, Yang, Gao, Liu, Shao and Lu2023; Li et al. Reference Li, Li, Hesselbo, Li, Liu, Wu, Pan and Gao2024). The fossil macroflora of the Huoshaoshan Formation is composed of sphenophytes (e.g., Neocalamites Halle, Equisetites Sternberg), ferns (e.g., Cladophlebis Brongniart), cycads (e.g., Ctenis Lindley et Hutton, Anomozamites Schimper emend. Harris), ginkgophytes (e.g., Ginkgoites Seward emend. Florin, Sphenobaiera Florin) and conifers (e.g., Podozamites A. Braun, Elatocladus Halle emend. Harris) (Li et al. Reference Li, He, Wu, Mei and Li1988; Zhang et al. Reference Zhang, Li, Xiong, Zhang, Wang, He, Lin and Sun1998). The palynoflora is represented by a Protoconiferus funarius–Chasmatosporites hians assemblage (Wang et al. Reference Wang, Mosbrugger and Zhang2005).
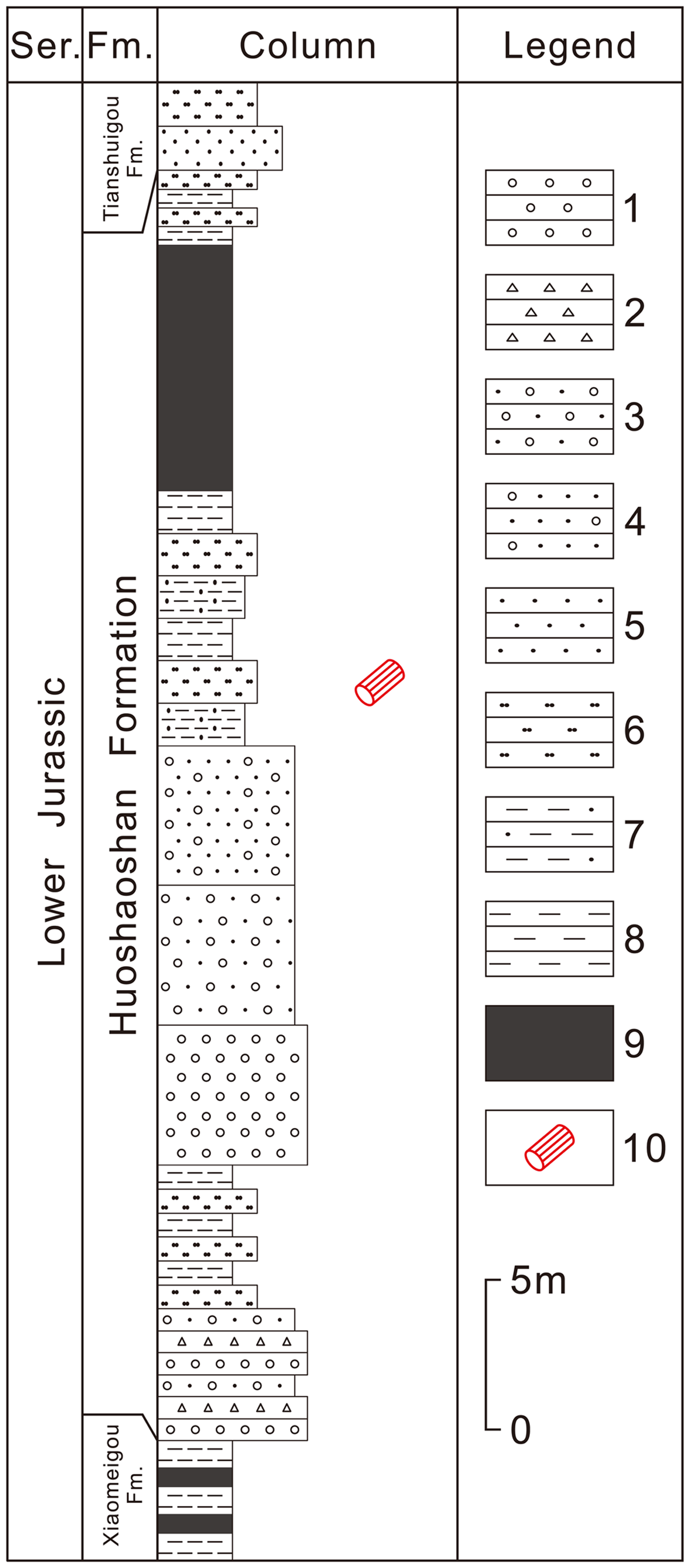
Figure 2. Stratigraphic column and fossil wood horizon of the Lower Jurassic Huoshaoshan Formation at Dameigou section in the Qaidam Basin, China (according to lithological descriptions of Zhang et al. Reference Zhang, Li, Xiong, Zhang, Wang, He, Lin and Sun1998). 1. Conglomerate; 2. Breccia; 3. Glutenite; 4. Conglomeratic sandstone; 5. Sandstone; 6. Siltstone; 7. Sandy mudstone; 8. Mudstone; 9. Coal seam; 10. Fossil wood.
3. Material and methods
For obtaining standard petrographic thin sections for wood identification, the fossil wood specimen was cut transversely, radially and tangentially in the three planes of section. Thin sections were investigated and observed under a Zeiss Imager M2 microscope; images were photographed with an Axiocam 506 colour digital camera and software ZEN 2 pro adapted to the microscope. The fossil specimen and 16 thin sections are deposited at the Nanjing Institute of Geology and Paleontology, Chinese Academy of Sciences (NIGPAS) with Catalog Number PB204109. For the description of fossil wood, we followed the terminology for wood anatomical structure defined by the International Association of Wood Anatomists (IAWA) list of microscopic features for softwood identification (IAWA Committee, 2004), supplemented by Philippe & Bamford (Reference Philippe and Bamford2008), and Boura et al. (Reference Boura, Bamford and Philippe2021).
4. Results
4.a. Systematic palaeobotany
Class: Pinopsida Burnett 1833
Order: Pinales (=Coniferales) Gorozhankin 1904
Family: Unknown
Genus: Metapodocarpoxylon Dupéron-Laudoueneix et Pons 1985
Species: M. libanoticum (Edwards) Dupéron-Laudoueneix et Pons 1985 (Figs. 3, 4)

Figure 3. Metapodocarpoxylon libanoticum from the Lower Jurassic Huoshaoshan Formation at Dameigou section in the Qaidam Basin, China; all photographs were taken of transverse (PB204109-a–b) and radial (PB204109-c, f, o) sections of specimen PB204109. (a) Transverse section, showing distinct growth rings (red arrows); PB204109-a; (b) Close-up of (a), transverse section, showing details of a distinct growth ring (red arrow); PB204109-a; (c) Transverse section, showing shapes of tracheids, and axial parenchyma cells (red arrows); PB204109-b; (d–f) Radial section, showing uniseriate contiguous and compressed (black arrows), as well as biseriate alternate radial tracheid pits (red arrows); PB204109-c; (g) Radial section, showing rarely uniseriate distant radial tracheid pits; PB204109-f; (h) Radial section, showing cross-field bearing one podocarpoid pit (white arrow), or one oopore (red arrow); PB204109-o.
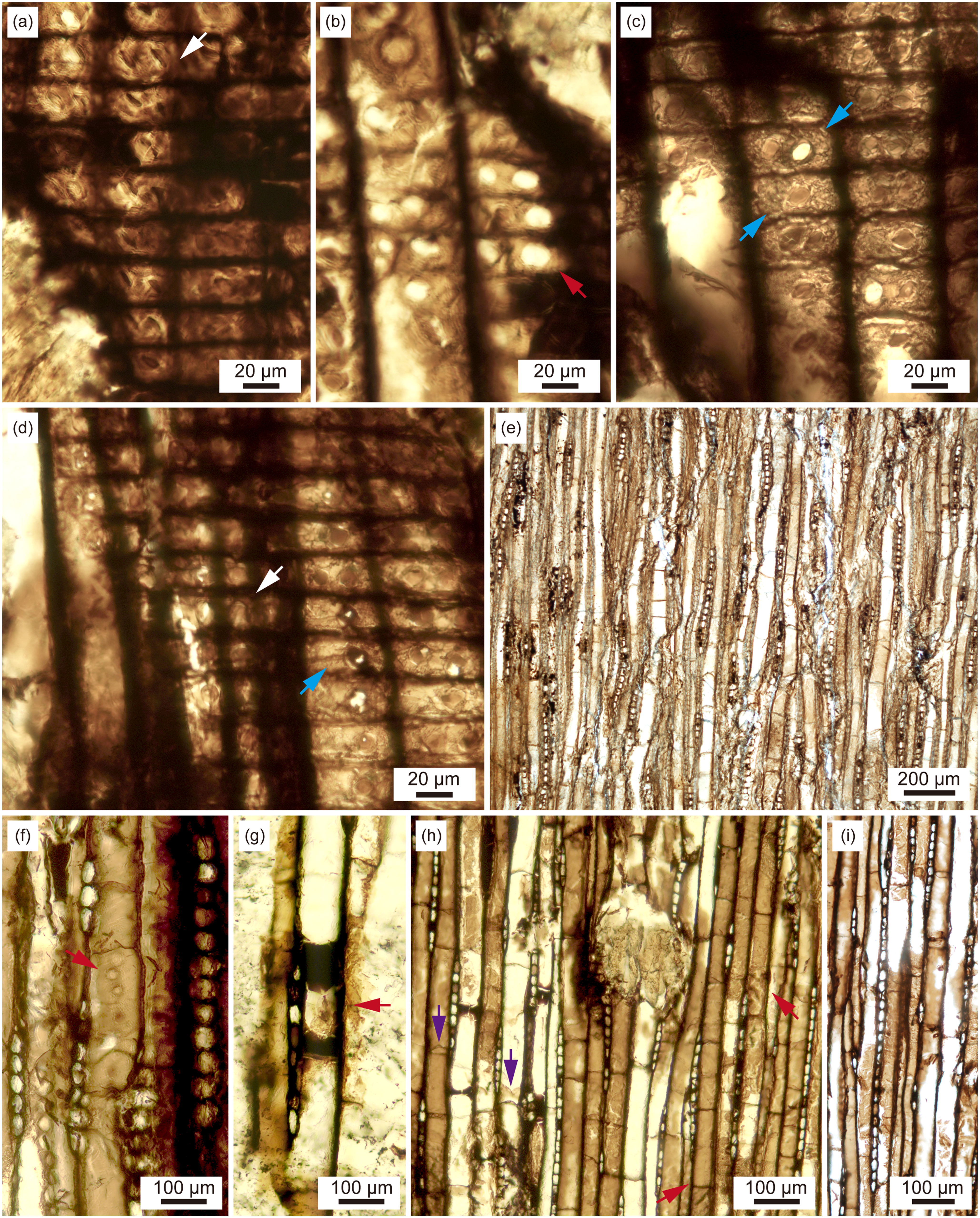
Figure 4. Metapodocarpoxylon libanoticum from the Lower Jurassic Huoshaoshan Formation at Dameigou section in the Qaidam Basin, China; all photographs were taken of radial (PB204109-c, e, o) and tangential (PB204109-p) sections of specimen PB204109. (a) Radial section, showing cross-field bearing two podocarpoid pits (white arrow); PB204109-o; (b) Radial section, showing cross-field bearing one oopore (red arrow); PB204109-e; (c) Radial section, showing cross-field bearing two taxodioid pits (blue arrows); PB204109-c; (d) Radial section, showing cross-field bearing one taxodioid pit (blue arrow), or one podocarpoid pit (white arrow); PB204109-c; (e) Tangential section, showing uniseriate rays; PB204109-p; (f) Tangential section, showing uniseriate tangential tracheid pits (red arrow); PB204109-p; (g) Tangential section, showing an axial parenchyma cell (red arrow); PB204109-p; (h) Tangential section, showing axial parenchyma cells (red arrows) and septa in tracheids (purple arrows); PB204109-p; (i) Tangential section, showing a high ray; PB204109-p.
Locality: Dachaidan Town, Delingha City, Qinghai Province, China.
Material studied: One specimen PB204109 with 16 thin sections (PB204109-a to PB204109-p).
Horizon and age: Huoshaoshan Formation, Early Jurassic (Pliensbachian).
Repository: The specimens and slides are housed in the Paleobotanical Collection, NIGPAS, Nanjing, China.
Description: This description is based on secondary xylem. Bark, pith and primary xylem are not preserved. In transverse section, growth rings are distinct (Fig. 3a, b, red arrows). The earlywood band has a mean width of 2.51 mm (2.09–2.97 mm; median = 2.50 mm; n = 5). Earlywood tracheids are large, thin-walled, with an oval to polygonal shape in cross section (Fig. 3b, c). Earlywood cells measure on average 67 µm (31–109 µm; median = 65 µm; n = 163) in radial diameter (height, the growth direction of tree growth rings), and on average 64 µm (29–101 µm; median = 63 µm; n = 163) in tangential diameter (width). The latewood is composed of narrow bands of tracheids, usually 3 rows wide; the latewood tracheids are somewhat thicker-walled than those of the earlywood tracheids (Fig. 3b). The latewood band has a mean width of 87 µm (70–120 µm; median = 84 µm; n = 5). Latewood cells measure on average 22 µm (15–38 µm; median = 20 µm; n = 28) in radial diameter (height, the growth direction of tree growth rings), and on average 50 µm (20–72 µm; median = 52 µm; n = 28) in tangential diameter (width). The cross-sectional shape of tracheids in the latewood is narrow, from narrowly elliptical to narrowly polygonal (Fig. 3b). The abrupt transition between earlywood and latewood is characterized by size differences of the tracheids in radial diameter and wall thickness (Fig. 3a, b, red arrows). Axial parenchyma cells are present (Fig. 3c, red arrows), whereas the resin canals are absent.
In radial section, tracheidal pits are uniseriate and biseriate; when uniseriate, they are usually contiguous and compressed (Fig. 3d–f, black arrows), and rarely distant (Fig. 3g); when biseriate, the arrangement of radial tracheid pits is alternating (Fig. 3d–f, red arrows). These pits are generally oblate in outline. Crassulae were not observed. Cross-field pits vary in size and borders (oculipores and oopores). Each cross-field usually bears one to two pits (Figs. 3h, 4a–d). These pits are oculipores with an oblique slit-like aperture (podocarpoid) (Figs. 3h, 4a, d, white arrows), or oculipores with a broad elliptic aperture (taxodioid) (Fig. 4c, d, blue arrows), or oopores (Figs. 3h, 4b, red arrows). The rays consist of parenchymatous cells with thin, smooth horizontal and end walls (Figs. 3h, 4a–d). Helical thickenings are not present.
In tangential section, rays are homogenous, parenchymatous and uniseriate (Fig. 4e–i). The rays are variable in height, ranging from very low to very high, 3–35 cells high, mostly low, 3–13 cells tall (n = 291) (Fig. 5). Tangential tracheid pits are occasionally present; they are uniseriate, contiguous and circular (Fig. 4f, red arrow). Axial parenchyma cells are present, with smooth end walls (Fig. 4g, h, red arrows). Abundant septa are preserved in tracheids (Fig. 4e, h, purple arrows). Resin canals are not observed.

Figure 5. Ray cell heights in 291 rays as observed in a tangential section (PB204109-p) of Metapodocarpoxylon libanoticum from the Lower Jurassic Huoshaoshan Formation at Dameigou section in the Qaidam Basin, China; specimen PB204109.
4.b. Growth ring characteristics of Metapodocarpoxylon libanoticum
As mentioned above, distinct growth rings (GR1 to GR5) are observed in our fossil specimen PB204109, M. libanoticum (Fig. 6). On average, the earlywood band has a width of 2.51 mm on average (2.09–2.97 mm; median = 2.50 mm; n = 5). Earlywood cells measure on average 67 µm (31–109 µm; median = 65 µm; n = 163) in radial diameter (height), and on average 64 µm (29–101 µm; median = 63 µm; n = 163) in tangential diameter (width). The latewood consists of narrow bands of tracheids, usually 3 rows wide. The latewood band has a mean width of 87 µm (70–120 µm; median = 84 µm; n = 5). Latewood cells measure on average 22 µm (15–38 µm; median = 20 µm; n = 28) in radial diameter (height), and on average 50 µm (20–72 µm; median = 52 µm; n = 28) in tangential diameter (width). The transition between earlywood and latewood is abrupt.

Figure 6. Growth rings (GR1–GR5) in Metapodocarpoxylon libanoticum from the Lower Jurassic Huoshaoshan Formation at Dameigou section in the Qaidam Basin, China; specimen PB204109 with thin-section PB204109-a. All growth rings are distinct with narrow latewood, usually 3 rows wide.
5. Discussion
5.a. Taxonomic assignment and comparisons
In general, five different arrangements of radial tracheid pits are known from Mesozoic conifer-like wood: araucarian, abietinean, brachyoxylean (mixed), shimakurean (japonicum-type) and xenoxylean radial tracheid pitting (Philippe & Bamford, Reference Philippe and Bamford2008; Philippe et al. Reference Philippe, Boura, Oh and Pons2014; Boura et al. Reference Boura, Bamford and Philippe2021). In our study, the bordered pits on the radial tracheid walls in fossil specimen PB204109 from the Huoshaoshan Formation in Dachaidan are usually araucarian radial tracheid pits. The characters of cross-field pits, tracheids and ray cells in our specimen are represented by these aspects, such as each cross-field usually bearing 1–2 podocarpoid pits, taxodioid pits or oopores, abundant septa in tracheids, and all ray cell walls thin and unpitted. These anatomical characters demonstrate that the fossil wood PB204109 can be systematically assigned to the genus Metapodocarpoxylon Dupéron-Laudoueneix et Pons (Dupéron-Laudoueneix & Pons, Reference Dupéron-Laudoueneix and Pons1985; Bamford et al. Reference Bamford, Roberts, Sissoko, Bouaré and O’leary2002; Philippe & Bamford, Reference Philippe and Bamford2008).
Metapodocarpoxylon was first established by Dupéron-Laudoueneix and Pons in Reference Dupéron-Laudoueneix and Pons1985 when reviewing an old fossil wood specimen (type slides and duplicated thin-sections) from the Lower Cretaceous of Lebanon, which was originally described as Mesembrioxylon libanoticum Edwards (Edwards, Reference Edwards1929). Dupéron-Laudoueneix and Pons formally defined Metapodocarpoxylon as “homoxylous wood without resin canals; radial tracheid pitting usually araucarian type; each cross-field with one oopore, or 1–2 podocarpoid oculipores, or many transition forms; homogenous rays with smooth walls; axial parenchyma present; thyllosis septa in tracheids” (Dupéron-Laudoueneix & Pons, Reference Dupéron-Laudoueneix and Pons1985; Philippe & Bamford, Reference Philippe and Bamford2008). Following this definition, a bunch of fossil wood taxa were subsequently reassigned to its type species M. libanoticum (Edwards) Dupéron-Laudoueneix et Pons, such as M. libanoticum Edwards, Protophyllocladoxylon chudeaui Batton, P. curitiense Pons, P. diphtericum Batton et Boureau, P. leuchsii Kräusel, P. libanoticum (Edwards) Kräusel, P. maurianum Gazeau, P. rosablancaense Pons, P. subdiphtericum Duperon-Laudoueneix and Xenoxylon saadawi Youssef (Philippe et al. Reference Philippe, Cuny, Bamford, Jaillard, Barale, Gomez, Ouaja, Thévenard, Thiébaut and Von Sengbusch2003; Philippe & Wilde, Reference Philippe and Wilde2020). Most of these reassignments refer to species previously assigned into the genus Protophyllocladoxylon Kräusel. The reason is that the difference between Metapodocarpoxylon and Protophyllocladoxylon is very small; that is, the cross-field pits of Metapodocarpoxylon exhibit both oopores and oculipores, whereas those of Protophyllocladoxylon are only oopores (Kräusel, Reference Kräusel1939; Dupéron-Laudoueneix & Pons, Reference Dupéron-Laudoueneix and Pons1985; Philippe & Bamford, Reference Philippe and Bamford2008; Philippe & Wilde, Reference Philippe and Wilde2020). Concerning the anatomical character of oopores and oculipores in the cross-fields in our fossil wood specimen, its assignment to the genus Metapodocarpoxylon mentioned above is reliable, instead of Protophyllocladoxylon.
Metapodocarpoxylon is monospecific, and only one species, M. libanoticum has been described so far, although about 40 occurrences of Metapodocarpoxylon have so far been reported from Gondwana (Dupéron-Laudoueneix & Pons, Reference Dupéron-Laudoueneix and Pons1985; Philippe et al. Reference Philippe, Cuny, Bamford, Jaillard, Barale, Gomez, Ouaja, Thévenard, Thiébaut and Von Sengbusch2003; El-Noamani et al. Reference El-Noamani, El-Saadawi and Philippe2021). In general, the previously reported specimens of M. libanoticum obtain low rays, which is similar to the rays in our specimen (uniseriate, mostly 3–13 cells high). For example, the type specimen M. libanoticum from Lebanon is characterized by having uniseriate rays (1–14 cells high) (Dupéron-Laudoueneix & Pons, Reference Dupéron-Laudoueneix and Pons1985). The fossil specimen of M. libanoticum from Egypt obtains slightly shorter rays, i.e. mostly 1–7 cells high, very rarely 15 cells high (El-Noamani et al. Reference El-Noamani, El-Saadawi and Philippe2021). The fossil wood M. libanoticum from Mali exhibits uniseriate rays with mostly seven cells high (1–10 cells high) (Bamford et al. Reference Bamford, Roberts, Sissoko, Bouaré and O’leary2002). In addition, tangential tracheid pits are observed in the type specimen from Lebanon and our specimen from China. Combined with other generic characters mentioned above, our fossil wood specimen is thus recognized as M. libanoticum (Edwards) Dupéron-Laudoueneix et Pons.
5.b. Palaeobiogeography of Metapodocarpoxylon
With the discovery of our new fossil wood material, the occurrence of M. libanoticum in the Early Jurassic offers new insights into the palaeobiogeographic distribution of Metapodocarpoxylon. During the Mesozoic, Metapodocarpoxylon was widespread in Northern Gondwana with a clear latitudinal belt extending from Lebanon westward to Peru (Dupéron-Laudoueneix & Pons, Reference Dupéron-Laudoueneix and Pons1985; Bamford et al. Reference Bamford, Roberts, Sissoko, Bouaré and O’leary2002; Philippe et al. Reference Philippe, Cuny, Bamford, Jaillard, Barale, Gomez, Ouaja, Thévenard, Thiébaut and Von Sengbusch2003; El-Noamani et al. Reference El-Noamani, El-Saadawi and Philippe2021) (Fig. 7). The fossil wood Metapodocarpoxylon described here from the Huoshaoshan Formation represents the first report of this fossil wood genus in the Qaidam Basin in China.
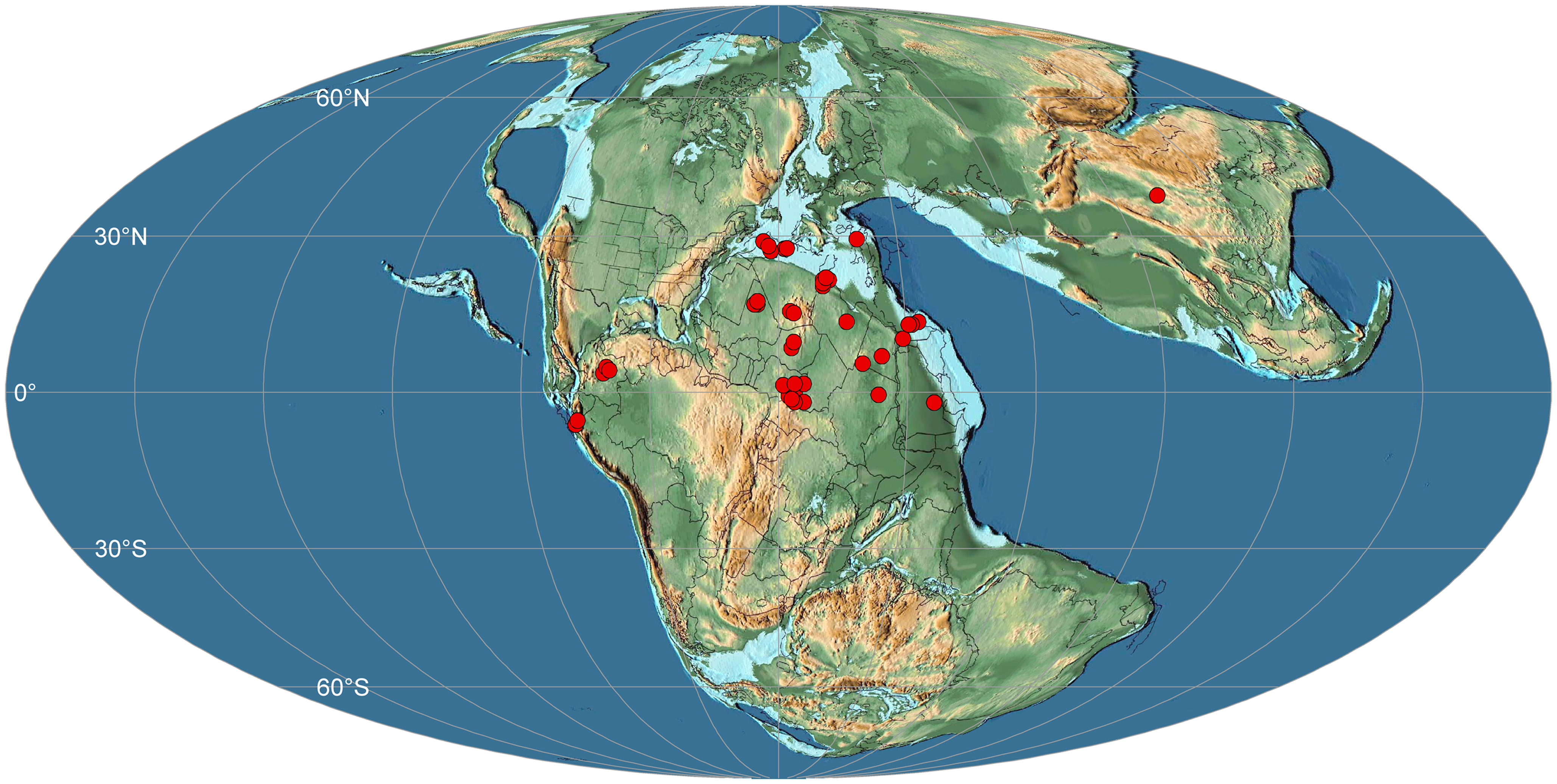
Figure 7. Palaeobiogeographic map showing the spatio–temporal distribution of Metapodocarpoxylon worldwide. The red dots indicate the occurrence of Metapodocarpoxylon (data from Philippe et al. Reference Philippe, Cuny, Bamford, Jaillard, Barale, Gomez, Ouaja, Thévenard, Thiébaut and Von Sengbusch2003, Reference Philippe, Bamford, Mcloughlin, Alves, Falcon-Lang, Gnaedinger, Ottone, Pole, Rajanikanth, Shoemaker, Torres and Zamuner2004, with updates). The base palaeogeographic map is modified after Scotese (Reference Scotese2014): Toarcian (∼180 Ma), Early Jurassic.
So far, only a single species of Metapodocarpoxylon is recognized worldwide; however, about 40 occurrences of this genus have been documented since the early twentieth century (Fig. 7) (Philippe et al. Reference Philippe, Cuny, Bamford, Jaillard, Barale, Gomez, Ouaja, Thévenard, Thiébaut and Von Sengbusch2003, Reference Philippe, Bamford, Mcloughlin, Alves, Falcon-Lang, Gnaedinger, Ottone, Pole, Rajanikanth, Shoemaker, Torres and Zamuner2004; El-Noamani et al. Reference El-Noamani, El-Saadawi and Philippe2021). So far there are no pre-Jurassic records of Metapodocarpoxylon. When the current study of Metapodocarpoxylon is considered, it implies that this genus would have originated in the Early Jurassic. In the Middle Jurassic, there are seven occurrences of Metapodocarpoxylon from Morocco (Gazeau, Reference Gazeau1970; Dupéron-Laudoueneix & Pons, Reference Dupéron-Laudoueneix and Pons1985) and Israel (Philippe et al. Reference Philippe, Cuny, Bamford, Jaillard, Barale, Gomez, Ouaja, Thévenard, Thiébaut and Von Sengbusch2003). In the Late Jurassic, five occurrences of this genus have been reported from Colombia (Pons, Reference Pons1982), Algeria (Benest et al. Reference Benest, Bensalah, Bouabdellah and Ouardas1999; Philippe et al. Reference Philippe, Cuny, Bamford, Jaillard, Barale, Gomez, Ouaja, Thévenard, Thiébaut and Von Sengbusch2003), Tunisia (Barale et al. Reference Barale, Zarbout and Philippe1998, Reference Barale, Ouaja and Philippe2000) and Lebanon (Edwards, Reference Edwards1929).
During the Early Cretaceous, Metapodocarpoxylon reached a significant period of prosperity in geographic distribution, as 28 occurrences have been documented from Italy (Biondi, Reference Biondi1980), Colombia (Pons, 1978; Reference Pons1988), Ecuador (Philippe et al. Reference Philippe, Cuny, Bamford, Jaillard, Barale, Gomez, Ouaja, Thévenard, Thiébaut and Von Sengbusch2003), Peru (Philippe et al. Reference Philippe, Cuny, Bamford, Jaillard, Barale, Gomez, Ouaja, Thévenard, Thiébaut and Von Sengbusch2003), Morocco (Attims & Crémier, Reference Attims and Crémier1969), Tunisia (Barale et al. Reference Barale, Zarbout and Philippe1998; Philippe et al. Reference Philippe, Barale, Gomez, Guignard and Thévenard1999; Philippe et al. Reference Philippe, Cuny, Bamford, Jaillard, Barale, Gomez, Ouaja, Thévenard, Thiébaut and Von Sengbusch2003), Algeria (Philippe et al. Reference Philippe, Cuny, Bamford, Jaillard, Barale, Gomez, Ouaja, Thévenard, Thiébaut and Von Sengbusch2003), Mali (Dupéron-Laudoueneix & Pons, Reference Dupéron-Laudoueneix and Pons1985; Bellion et al. Reference Bellion, Lefranc and Micahrd1992; Bamford et al. Reference Bamford, Roberts, Sissoko, Bouaré and O’leary2002), Niger (Batton, Reference Batton1965), Nigeria (Philippe et al. Reference Philippe, Cuny, Bamford, Jaillard, Barale, Gomez, Ouaja, Thévenard, Thiébaut and Von Sengbusch2003), Cameroon (Batton & Boureau, Reference Batton and Boureau1965; Dupéron-Laudoueneix, Reference Dupéron-Laudoueneix1976, Reference Dupéron-Laudoueneix1991a , Reference Dupéron-Laudoueneix1991b; Flynn et al. Reference Flynn, Brillanceau, Brunet, Coppens, Dejax, Duperon-Laudoueneix, Ekodeck, Flanagan, Heintz, Hell, Jacobs, Pilbeam, Sen and Djallo1988; Brunet et al. Reference Brunet, Dejax, Flynn and Jehenne1990; Dejax & Brunet, Reference Dejax and Brunet1995), Sudan (Philippe et al. Reference Philippe, Cuny, Bamford, Jaillard, Barale, Gomez, Ouaja, Thévenard, Thiébaut and Von Sengbusch2003), Lebanon (Philippe et al. Reference Philippe, Cuny, Bamford, Jaillard, Barale, Gomez, Ouaja, Thévenard, Thiébaut and Von Sengbusch2003) and Saudi Arabia (Philippe et al. Reference Philippe, Cuny, Bamford, Jaillard, Barale, Gomez, Ouaja, Thévenard, Thiébaut and Von Sengbusch2003).
Recently, a study on the Cretaceous terrestrial biota of northern Africa offered new evidence for the Late Cretaceous record of Metapodocarpoxylon in Egypt (El-Noamani et al. Reference El-Noamani, El-Saadawi and Philippe2021). In addition, some records of Metapodocarpoxylon are reported from Middle Jurassic to Lower Cretaceous strata in Libya (Philippe et al. Reference Philippe, Cuny, Bamford, Jaillard, Barale, Gomez, Ouaja, Thévenard, Thiébaut and Von Sengbusch2003), and Upper Jurassic to Lower Cretaceous strata in Egypt (Youssef, Reference Youssef2002), without a finer stratigraphic resolution.
Based on the occurrences of Metapodocarpoxylon reported so far, this genus had probably a wide occurrence in the Middle Jurassic–Cretaceous deposits of northern Gondwana, along with the only record in the Early Jurassic sediments of Laurasia presented in this study. It seems that Metapodocarpoxylon may have originated in Laurasia during the Early Jurassic, then migrated to Gondwana maybe owing to some possible climatic changes. However, we realize that this hypothesis may need support from other robust evidence owing to the limited fossil material of Metapodocarpoxylon in our current fossil locality. Thus, further analysis of additional and so far understudied occurrences of fossil wood from the Mesozoic, especially in the Middle East and other regions, would undoubtedly enhance our knowledge of the palaeogeographic patterns observed in Metapodocarpoxylon. Nevertheless, our new finding does extend the palaeobiogeographic range of Metapodocarpoxylon northward and eastward from northern Gondwana regions to Laurasia.
5.c. Palaeoclimatic implications of Metapodocarpoxylon
In the Metapodocarpoxylon wood from the Qaidam Basin, distinct growth rings with abrupt transitions between earlywood and latewood were well-developed when this tree was growing (Fig. 6). The latewood contains narrow bands of tracheids, usually 3 rows wide. These true growth rings with narrow latewood in our fossil wood M. libanoticum suggest that this tree’s growth conditions were slightly variable concerning mild seasonality during the whole year, and demonstrate that it enjoyed a long and optimal growth season of the year, then underwent slower growth for a short time. Moreover, Philippe et al. (Reference Philippe, Cuny, Bamford, Jaillard, Barale, Gomez, Ouaja, Thévenard, Thiébaut and Von Sengbusch2003) proposed that Metapodocarpoxylon trees grew in the summer-wet climatic zone in a palaeoclimatological map drawn by Rees et al. (Reference Rees, Ziegler, Valdes, Huber, Macleod and Wing1999) based on the occurrences of this genus. Following modern Köppen-Geiger climate classification, this summer-wet climate can be found in the Tropical Monsoon Climate, Humid Subtropical Climate and Temperate Climate zones (cf. Kottek et al. Reference Kottek, Grieser, Beck, Rudolf and Rubel2006), which is characterized by a distinct pattern of precipitation, that is, most rainfall occurs during the warmer months of the year. Putting them all together, we can assume that the Metapodocarpoxylon tree in the Qaidam Basin was growing in a usually warm and humid environment with mild seasonality during the Early Jurassic.
Palynological studies also play a significant role in demonstrating that a warm and humid climate prevailed throughout deposition of the Huoshaoshan Formation. The palynoflora of the Pliensbachian in the Qaidam Basin is represented by an assemblage of Protoconiferus funarius–Chasmatosporites hians, with a high mesophyte index and a zero thermophyte/xerophyte index (Wang et al. Reference Wang, Mosbrugger and Zhang2005). The dominant Pinaceae pollen indicates a temperate climate, and other plants demonstrate a humid tropical–subtropical climate. This climate condition is also consistent with the leaf flora assemblage in the Huoshaoshan Formation, which is dominated by ferns and conifers, along with some cycads and ginkgophytes, indicating a temperate and humid climate (Li et al. Reference Li, He, Wu, Mei and Li1988; Zhang et al. Reference Zhang, Li, Xiong, Zhang, Wang, He, Lin and Sun1998).
A warm and humid climate during the Early Jurassic in the Qaidam Basin is also supported by some recent geochemical investigations, based on chemical weathering parameters (e.g., chemical index of alteration, chemical index of weathering, as well as plagioclase index of alteration and clay mineral assemblages), and element geochemistry indices (e.g., Rb/Sr, K2O/Na2O, Na2O/A12O3, Na2O/TiO2 ratios, TOC and δ13Corg values) (Hu et al. Reference Hu, Ma, Li, Wu, Gao, Peng, Wei and Liu2020; Lu et al. Reference Lu, Zhou, Yang and Shao2020; Zhou et al. Reference Zhou, Lu, Zhang, Yang, Gao, Shao and Hilton2022, Reference Zhou, Wang, Yang, Gao, Liu, Shao and Lu2023).
Integrating the evidence of the present fossil wood, together with other data including leaf, sporopollen and sediments, the Early Jurassic deposit of the Huoshaoshan Formation of the Qaidam basin is interpreted as a warm and humid climate environment with mild seasonality.
Data availability
All the related data are described in the study.
Acknowledgements
This work was co-sponsored by the National Natural Science Foundation of China (NSFC 42330208, 42288201, 41790454, 41972007, 42002023, 42372044, 41972022), Strategic Priority Research Program (B) of the Chinese Academy of Sciences (XDB26000000), and the State Key Laboratory of Paleobiology and Stratigraphy (20172103, 20191103, 20192101). We thank Dr. Bas Van de Schootbrugge (Editor, Geological Magazine), and an anonymous reviewer for helpful comments.
Competing interests
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.








 Open access
Open access