


Introduction
Determining population status is important for effective wildlife management (Krausman, Reference Krausman2002; Bolen & Robinson, Reference Bolen and Robinson2003). Additionally, knowledge of abundances and trends are required to determine the conservation status of a species (IUCN, 2001) and the actions needed to conserve populations (Nowell & Jackson, Reference Nowell and Jackson1996), and to evaluate the performance of conservation programmes. In addition, understanding the factors underpinning variation in abundance allows managers to identify conservation interventions.
Despite the critical status of the Sumatran tiger Panthera tigris sumatrae (Morell, Reference Morell2007; Linkie et al., Reference Linkie, Wibisono, Martyr and Sunarto2008b), the population status of this subspecies in the wild remains uncertain (Soehartono et al., Reference Soehartono, Wibisono, Sunarto, Martyr, Susilo and Maddox2007; Wibisono & Pusparini, Reference Wibisono and Pusparini2010). Although tiger density has been well documented in some areas, trends in population size across habitat types and over time remain poorly documented in Sumatra.
Historically, Riau Province was estimated to hold c. 30% of the Sumatran tiger population (Borner, Reference Borner1978). Riau harbours not only hill and lowland forests, habitat types surveyed in previous studies in Sumatra, but also extensive peatland forests, which have never previously been surveyed for tigers. Although Riau Province has had an estimated deforestation rate of 65% over 25 years (Uryu et al., Reference Uryu, Mott, Foead, Yulianto, Budiman and Setiabudi2007) it still holds areas with high potential for tiger conservation and recovery (Sanderson et al., Reference Sanderson, Forrest, Loucks, Ginsberg, Dinerstein and Seidensticker2006; Walston et al., Reference Walston, Robinson, Bennett, Breitenmoser, da Fonseca and Goodrich2010; Wikramanayake et al., Reference Wikramanayake, Dinerstein, Seidensticker, Lumpkin, Pandav and Shrestha2011).
We investigated tiger abundance across the various forest types in southern Riau Province and monitored changes in abundance over time in the newly established Tesso Nilo National Park. Our goals were to gain understanding of spatial and temporal variation in tiger density, to identify the factors that influence tiger numbers, and to explore management actions needed to conserve and restore tigers in the landscape.
Study area
This study was conducted in several forest blocks in southern Riau Province in central Sumatra (Fig. 1; Table 1; Sunarto, Reference Sunarto2011). Systematic photographic sampling using camera traps was conducted in five blocks covering three forest types: the peat swamp forest of Kampar Peninsula and Kerumutan (Plate 1), the flat lowland forest of Tesso Nilo and Peranap, and the hilly lowland forest of Rimbang Baling. Except in Kampar Peninsula and Peranap, the blocks are protected as either wildlife reserves (Kerumutan and Rimbang Baling) or as a National Park (Tesso Nilo). These forest blocks, interspersed with non-forested lands comprising mainly acacia and oil palm plantations, form a multi-use landscape covering a total area of c. 30,000 km2.

Fig. 1 The study area, showing forest cover in 2007, the five sampling blocks (Table 1) in Riau Province, central Sumatra, and the locations of camera traps. The rectangle on the inset indicates the location of the main map in central Sumatra.

Plate 1 An adult female tiger photo-trapped in inundated peatland in Kerumutan (Fig. 1).
Table 1 Characteristics of the five sampling blocks (Fig. 1) in southern Riau Province, central Sumatra.

1 From a combination of geological, bioclimatic, geomorphological and topographical maps, as identified by Laumonier (Reference Laumonier1997)
2 Calculated by measuring forest area based on interpretation of Landsat images acquired in 2007 (by WWF Indonesia) corrected with 3-km buffer, assuming an edge effect
Methods
We estimated population densities of tigers using a capture–recapture approach, with standardized camera trapping procedures (Fig. 1), over 3 months for each sampling period to avoid violation of the closure assumption (Nichols & Karanth, Reference Karanth, Nichols, Karanth and Nichols2002). Each sampling block consisted of c. 40, 2 × 2 km grid cells and we established at least 20 camera stations in each block, in every other grid cell. This spacing assured at least three pairs of cameras in each tiger's home range, assuming a home range size of c. 50 km2 in Sumatra (Franklin et al., Reference Franklin, Bastoni, Siswomartono, Manansang, Tilson, Seidensticker, Christie and Jackson1999).
We placed cameras non-randomly (Karanth et al., Reference Karanth, Kumar, Nichols, Karanth and Nichols2002), based on characteristics such as suitable habitat and presence of tiger sign, within each pre-determined grid cell, to optimize captures of tigers whilst avoiding human disturbance/vandalism. At each site we installed a pair of opposing cameras c. 50 cm above the ground, at c. 2–3 m from the trail where animals were likely to pass. Individual tigers were distinguished by their unique stripe patterns (Karanth, Reference Karanth1995). The sampling period for each location is presented in Table 2.
Table 2 Survey period, number of camera-trap stations and effective trap nights, and camera-trapping effort and photographic capture rates (number of independent photographs per 100 trap nights) of humans and main potential tiger Panthera tigris prey in the five sampling blocks (Fig. 1).

1 Counted based on the position in 2 × 2 km grids. In the case of cameras being moved during the survey period for security or other reasons, they are considered as one trap station if location is still in the same 2 × 2 km grid. In the case of total loss/failure of cameras in a given grid cell during a survey period the trap station was not counted
2 Includes the presence of forest dweller indigenous people who live mainly by subsistence hunting and gathering
Density estimation
We applied two approaches for estimating tiger density. Firstly, we used spatially explicit capture–recapture modelling within a maximum likelihood framework (Efford, Reference Efford2004; Efford et al., Reference Efford, Dawson and Robbins2004), implemented in DENSITY 4.4 (Efford, Reference Efford2010). In the detection history one occasion represents a 24-hour period of camera trapping. For every occasion we marked each camera station as active (1) when at least one camera was operational, or inactive (0) when no camera was working, and enabled incomplete trap layout in the input option of DENSITY. We used forest cover geographical information system layers from the same year of camera trapping (available from WWF; Uryu et al., Reference Uryu, Mott, Foead, Yulianto, Budiman and Setiabudi2007, Reference Uryu, Purastuti, Laumonier, Sunarto, Setiabudi and Budiman2010), and added a 4-km buffer as the habitat mask. Tiger density was estimated using conditional likelihood with three detection functions (half-normal, hazard rate and negative exponential) assuming a Poisson distribution, all with a constant model for the intercept (i.e. the probability of capture when the trap and range centre coincide, g 0[.]) and spatial scale parameter (σ [.]). Covariates (such as response to capture) were not used because of the small sample sizes of captures and recaptures. We used the Akaike Information Criteria (AIC; Akaike, Reference Akaike, Petrov and Csaki1973) for model selection, or that corrected for small sample size (AICc; Burnham & Anderson, Reference Burnham and Anderson1998).
Secondly, for comparison with other studies in Sumatra using the same approach, we used CAPTURE (Rexstad & Burnham, Reference Rexstad and Burnham1992) to generate estimates of tiger abundance using K-sample closed population capture–recapture models (Nichols & Karanth, Reference Karanth, Nichols, Karanth and Nichols2002). As many zeros caused problems in the analysis with CAPTURE we collapsed every 10-day period into one sampling occasion. With a c. 3-month survey period this gave us 9–10 encounter occasions in the detection history. When the only competing model was M 0 (model assuming equal capture probability for all animals) we used the heterogeneity model (M h) with Jackknife estimator, which allows each individual to have different and unique detection probabilities (Otis et al., Reference Otis, Burnham, White and Anderson1978) and is considered robust when heterogeneity is assumed to occur for territorial animals such as tigers (Nichols & Karanth, Reference Karanth, Nichols, Karanth and Nichols2002).
We calculated the density of tigers by dividing the abundance estimate by the area from which animals were sampled (Karanth & Nichols, Reference Karanth and Nichols1998). We estimated the area sampled as the polygon connecting the outermost cameras plus a buffer (the minimum convex polygon) of half the mean maximum distance moved (½MMDM) by individuals captured at multiple stations (Karanth & Nichols, Reference Karanth and Nichols1998). We calculated variance in density using the delta method (Nichols & Karanth, Reference Karanth, Nichols, Karanth and Nichols2002). For sampling blocks that yielded too few recaptures of tigers at more than one location we borrowed recapture/movement distance data from the nearest blocks to calculate variance in density.
As an indicator of disturbance we assessed the level of human activity in each sampling block using the rate humans were photographed and level of vandalism to the cameras. Using χ2 goodness-of-fit statistics we tested between expected (null hypothesis of no difference) and observed numbers of independent photographs of the main tiger prey species among sites, habitat types and for Tesso Nilo National Park, among years of sampling. The expected numbers of independent photographs were calculated based on the proportion of effective trap nights for a given sampling block, forest type, or survey period. We defined photographs as independent following O'Brien et al. (Reference O'Brien, Kinnaird and Wibisono2003). Statistical tests were performed in Excel 2007 (Microsoft, Redmond, USA) or R v. 2.10.1 (R Development Core Team, 2008).
We defined effective trap nights as the number of nights multiplied by the number of camera stations minus any days where both cameras at a station malfunctioned. Paired cameras that operated for the same 24-hour period at the same station were considered one trap night. We calculated the photographic capture rates (O'Brien et al., Reference O'Brien, Kinnaird and Wibisono2003) by calculating the number of independent photographs per 100 trap nights. We focused on humans and the preferred prey of tigers: medium to large ungulates (the wild pig Sus scrofa, bearded pig Sus barbatus, muntjac Muntiacus muntjak, and sambar deer Rusa unicolor) and ground-dwelling primates (the pig-tailed macaque Macaca nemestrina; Karanth & Sunquist, Reference Karanth and Sunquist1995; Carbone et al., Reference Carbone, Mace, Roberts and Macdonald1999; O'Brien et al., Reference O'Brien, Kinnaird and Wibisono2003; Sriyanto, Reference Sriyanto2003; Kawanishi & Sunquist, Reference Kawanishi and Sunquist2004).
We compiled published information on tiger abundance in Sumatra for comparison with our results. We identified location, terrain, habitat type, year, tiger abundance and estimation method used. Where available, we extracted the 95% confidence limits of the abundance estimate. If the confidence limits were not available we determined plausible lower and upper bounds given the description in the original text.
Results
From 10,690 effective trap nights we obtained a total of 174 photographs of tigers (Plate 1; Tables 2 & 3) from which we identified 18 individual tigers. Effective trap nights ranged from 1,132 (Kampar) to 1,868 (Kerumutan), and the number of active stations ranged from 18 (Kampar) to 25 (Tesso Nilo National Park). Overall, we obtained most samples, both the number of tiger photographs and number of unique individuals, from Tesso Nilo National Park. The number of individual tigers identified from Tesso Nilo in 2005, 2007 and 2008 was five, seven and six, respectively. Except for two tigers that were captured (and recaptured) in all three sampling years in the Park, other individuals were unique to each survey period.
Despite the presence of tiger sign in every sampling block we failed to photograph tigers in Kampar and Peranap during the systematic camera trapping. In these two blocks camera operations were disrupted by vandalism or theft of 16 cameras in Kampar and three in Peranap. In addition to losing camera units, we also documented relatively high human activity (Table 2).
Density estimates
Density estimates using the best spatially explicit capture–recapture models range from 0.34 individuals per 100 km2 in Rimbang Baling in 2006 to 0.87 per 100 km2 in Tesso Nilo in 2007 (Table 3 & 4). Tiger density in Tesso Nilo National Park from three different year samples estimated using spatially explicit capture–recapture remained relatively constant with no obvious trend. Estimates of tiger density in Kerumutan and Rimbang Baling were lower than those for any survey period in Tesso Nilo.
Table 3 Summary of the estimates of tiger density (individuals per 100 km2) in the five sampling blocks (Fig. 1).

1 Tigers confirmed to be present based on their sign (pug-mark and/or scat)
2 Number of independent photographs per 100 trap nights
3 Calculated based on minimum convex polygon of outer camera points buffered with ½MMDM; for sampling blocks without tiger photographic captures, ½MMDM was borrowed from the nearest block
4 Mean maximum distance moved (km)
5 Based on capture history, with one occasion representing 10 trap days
6 Estimated total number of individuals in the sampling block
7 Estimated density (individuals per 100 km2)
8 Because of the limited number of individuals photographed density was calculated by borrowing data on tiger distance moved from the neighbouring sampling blocks
Table 4 Model selection (ranked for each survey session based on AIC) and estimates of tiger density (individuals per 100 km2) for Kerumutan (in 2006), Tesso Nilo (in 2005, 2007, 2008) and Rimbang Baling (in 2006) based on spatially explicit capture–recapture models with conditional maximum likelihood estimators in DENSITY.

1 All models used did not consider covariates (g 0[.]s[.]); length of buffer used was 10,000 m
2 AIC, Akaike Information Criteria
3 AICc, AIC corrected for small sample size
4 W i, Akaike weights (the probability that the given model is the best model. The value ranges from 0 to 1.)
5 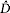 $\hat D$, estimated density (individuals per 100 km2)
$\hat D$, estimated density (individuals per 100 km2)
6 g0, intercept (the probability of capture when the trap and range centre coincide)
7 σ = sigma (a spatial scale parameter)
Estimates using spatially explicit capture–recapture models were consistently and substantially lower than those from traditional capture–recapture models. In CAPTURE abundance was estimated using model M h in all cases (Table 3). Closure tests indicated that the data did not violate the closure assumption at any site (P > 0.15), except in one survey in Tesso Nilo in 2005 (P = 0.04). With traditional capture–recapture models estimates of tiger density were 1.2 ± SE 0.54 individuals per 100 km2 in flat lowland forest of Tesso Nilo, 0.5 ± SE 0.27 per 100 km2 in peatland forest of Kerumutan, and 0.9 ± SE 0.43 per 100 km2 in hilly lowland forest of Rimbang Baling.
Although point estimates of tiger density varied, no estimate was found to be significantly different from any others because of their large confidence intervals. For Kampar and Peranap, where we did not photograph tigers but observed their sign, we assumed tiger density was lower than the site with the lowest density estimate (Kerumutan).
Human activity
Despite mitigation measures there was considerable vandalism of the camera stations and we lost 24 camera units during the systematic sampling. Only in Kerumutan and Rimbang Baling was there no trap loss. We documented high human activities in Kampar, Peranap and Tesso Nilo (Table 2).
Photographic capture rates of potential prey
Photographic capture rates of prey, by species or prey groups, was significantly different among sites (all χ2 > 61, df = 4, all P < 0.01) and among forest types (all χ2 > 20, df = 2, all P < 0.01; Table 2). Within Tesso Nilo National Park we found a significant difference among years only for muntjac (χ2 = 5.9, df = 2, P = 0.05) and no significance for the combination of three ungulate species (wild pig, muntjak, and sambar; χ2 = 4.23, df = 2, P = 0.12), whereas other species and combinations of ungulates and pig-tailed macaque varied significantly across the years (all χ2 > 13, df = 2, all P < 0.01).
Some species of ungulates were apparently absent from some areas. Sambar, for example, were photographed in only two sites, and in one of those sites captures were low, considering the effort. No sambar and zero, or extremely low numbers, of other ungulates were photographed in peatland forest (Table 2).
Comparison with tiger abundances in other areas
We found substantially lower tiger densities compared to studies elsewhere in Sumatra, which have ranged from 1.5–2.1 individuals per 100 km2 in submontane primary forest to 3.3 per 100 km2 in lowland primary forest (Table 5).
Table 5 Estimates of tiger density in various locations in Sumatra and, for comparison, the results from this study in the five sampling blocks in central Sumatra (in bold).

1 An asterisk indicates use of the standardized traditional capture–recapture approach (Karanth & Nichols, Reference Karanth, Nichols, Karanth and Nichols2002) to estimate tiger density
2 Estimate in Gunung Leuser was based on a combination of the total number of camera-trap photographs and extrapolation with a geographical information system, with density derived from estimates of the total population divided by total area (Griffiths, Reference Griffiths, Tilson, Sumarna, Ramono, Lusli, Traylor-Holzer and Seal1994)
3 Value presented is the median between the low and high estimates. Density in Way Kambas was estimated based on total number of resident individuals photographed over 16 months of sampling and additional projected monthly mean of non-resident individuals (Franklin et al., Reference Franklin, Bastoni, Siswomartono, Manansang, Tilson, Seidensticker, Christie and Jackson1999).
4 Maddox et al. (Reference Maddox, Priatna, Gemita and Salampessy2007) estimated the density based on photographic capture rate using the conversion factor presented by Carbone et al. (Reference Carbone, Coulson, Christie, Conforti, Seidensticker and Franklin2001)
5 Estimates were made based on traditional capture–recapture models with individuals identified based on pelage patterns on the left flank (O'Brien et al., Reference O'Brien, Kinnaird and Wibisono2003)
6 Estimate for montane forest was based on encounter rate (Linkie et al., Reference Linkie, Chapron, Martyr, Holden and Leader-Williams2006)
7 Wibisono et al. (Reference Wibisono, Figel, Arif, Ario and Lubis2009)
8 Linkie et al. (Reference Linkie, Haidir, Nugroho and Dinata2008a)
9 Although confirmed to be present, tigers were not photographed in camera traps during systematic camera-trap sampling. Density is, therefore, assumed to be lower than the site with the lowest density (Kerumutan)
10 Mean values from three consecutive estimates using traditional capture–recapture models (Table 3)
Discussion
This study is the first in Sumatra to investigate trends in tiger population across habitat types and over time, and the first to provide information on tiger abundance in Riau Province and in peatland areas. It is also the first study estimating the population of Sumatran tigers using spatially explicit capture–recapture models, and provides an insight into the ecology of a threatened large carnivore living in a landscape under intense pressure.
Although point estimates for tiger density in central Sumatra appear to vary by habitat, sampling block, and through time in the same area, our results had low precision. Our low sample sizes (i.e. the number of individuals photographed) were not because of lack of effort as our study had equal or larger numbers of trap nights and area sampled compared to other studies. For example, in India, Karanth et al. (Reference Karanth, Nichols, Kumar and Hines2006) photographed 74 individual tigers in a total of 5,725 total trap nights and took a mean of 7.73 trap nights to photograph one individual, whereas we photographed 18 individuals in 10,690 trap nights and took an average of 590 trap nights to photograph an individual.
Compared to other studies using similar techniques our estimates of tiger densities in Sumatra are lower than expected, often < 1 tiger per 100 km2, even compared to similar habitat elsewhere (Karanth & Nichols, Reference Karanth and Nichols1998; Kawanishi & Sunquist, Reference Kawanishi and Sunquist2004; Karanth et al., Reference Karanth, Nichols, Kumar and Hines2006; Linkie et al., Reference Linkie, Chapron, Martyr, Holden and Leader-Williams2006; Rayan & Mohamad, Reference Rayan and Mohamad2009; Table 5). Using spatially explicit capture–recapture models our estimates of tiger density were almost 50% lower compared to densities estimated with traditional capture–recapture models, a finding noted in other studies (Tredick & Vaughan, Reference Tredick and Vaughan2009; Obbard et al., Reference Obbard, Howe and Kyle2010; Sharma et al., Reference Sharma, Jhala, Qureshi, Vattakaven, Gopal and Nayak2010; Gerber et al., Reference Gerber, Karpanty and Kelly2012).
Prey availability
Prey availability has been identified as the single most important factor influencing tiger abundance (Karanth et al., Reference Karanth, Nichols, Kumar, Link and Hines2004), and depletion of prey significantly affects tiger population viability (Karanth & Smith, Reference Karanth, Smith, Seidensticker, Christie and Jackson1999). We found that photographic capture rate of prey per se was not a predictor of tiger density. For example, Peranap had a very low tiger density but the highest level of photographic capture rate of prey and the most diverse prey assemblage. Potential prey in this area included sambar, which were not detected in most of the other sites and is the preferred prey of tigers (Carbone & Gittleman, Reference Carbone and Gittleman2002).
Although we did not detect tigers by cameras in Peranap we found tiger pug-marks during camera placement. We believe that the low detection of tigers in this area was a result of the high level of human activity, including farming, hunting/trapping, and gathering of forest products by villagers, migrants or nomadic indigenous communities. Riau Province is known to be a major transit area for the illegal wildlife trade (Ng & Nemora, Reference Ng and Nemora2007) and one of the main sources of tiger body parts (Osmantri/WWF anti-poaching team leader, pers. comm.; Shepherd & Magnus, Reference Shepherd and Magnus2004). Notorious hunters, including one local person who claimed to have killed more than 50 tigers (Abi, 2010), formerly lived and operated in the Province. Unprotected areas such as Peranap can easily become the target for poachers.
The failure to detect tigers in Kampar is probably because of a combination of low prey availability and the high level of human activity. Prey populations are low in peatland forests, perhaps related to low primary productivity and a high level of plant defence mechanisms, discouraging herbivory (Whitten et al., Reference Whitten, Damanik, Anwar and Hisyam2000). The loose and porous texture of peat makes movement difficult, especially for ungulates with their relatively high body mass and pointed hooves. Additionally, many of these areas are inundated during the rainy season, making movement and feeding difficult in that period. With the ability to climb trees when needed to move and forage, pig-tailed macaques thrive in these areas, and are the main prey available for tigers.
Determinants of tiger abundance
The low capture rates and estimated densities of tigers documented in this study are most likely related to a combination of habitat quality, prey availability and human activity. High levels of human activity were expected because, during the last decade, Riau had the highest human population growth (c. 4%) in Indonesia (BAPPENAS, 2005). During their operation from 2007 to early 2010, WWF and the Forestry Department anti-poaching team detected and confiscated 87 metal snares typically used for tigers and 265 nylon snares used for other animals (Osmantri/WWF anti-poaching team leader, pers. comm.).
We experienced no camera loss in Kerumutan and Rimbang Baling. These are the only blocks in our survey that have been legally protected for 3 decades or longer, suggesting that long-term legal protection is recognized by local people and migrants, discouraging them from trespassing.
In northern Sumatra Griffiths & van Schaik (Reference Griffiths and van Schaik1993) documented significantly reduced activities of tigers and other wildlife species in areas with higher levels of human activity. We documented a decline in human activity over time and a stable tiger density in Tesso Nilo National Park in the same period. Our observations in the field suggested that in 2008 people appeared to halt activities following the establishment and active management of the National Park. A similar result was documented in Rajaji National Park, India, where tiger density increased following the removal of anthropogenic influences (Harihar et al., Reference Harihar, Pandav and Goyal2009), showing that strong protection can be effective in maintaining a high density of tigers (Karanth & Nichols, Reference Karanth and Nichols1998; Karanth et al., Reference Karanth, Nichols, Kumar, Link and Hines2004; Ahmed et al., Reference Ahmed, Borah, Das, Basumatary, Sarma and Gogoi2010).
Management implications and conclusion
The conservation community plans to double or significantly increase the global tiger population in the next decade (Global Tiger Recovery Program, 2010). This vision seems overly optimistic considering the widespread deforestation that is still taking place in many areas, including central Sumatra. It is possible, however, to increase tiger density if limiting factors can be removed. In central Sumatra the most important factor to control is the high level of illegal human activity. Legal protection of an area, followed by strong protection and intensive management can reduce the level of human disturbance and facilitate the recovery of the habitat and prey, and thus tigers.
Opportunities to protect legally or intensify management of tiger habitats are still available in Riau. For example, the majority of the Kampar Peninsula, a large portion of the Kerumutan block to the east, the Peranap forest area, and forests around Rimbang Baling Wildlife Reserve currently have little to no protection. Without action these areas may soon lose their forest because of ubiquitous and rampant conversion of forests to plantations.
Traditional protection such as that afforded by national parks and wildlife reserves has proven effective in reducing deforestation in some areas (Gaveau et al., Reference Gaveau, Epting, Lyne, Linkie, Kumara and Kanninen2009) but there are other alternatives to improve tiger habitat whilst promoting local and national economic development, such as through agroforestry or selective and sustainable logging. Conversion of forest into acacia and oil palm plantations is driven by the global demand for these products. Therefore, the international community needs to share the responsibility and join with national and local communities to take a range of measures to conserve the tiger.
Tigers appear to live at much lower densities in central Sumatra than documented elsewhere. This strongly indicates that tigers may not do well in some areas even when the forest is seemingly intact. More intensive monitoring and proactive management of tiger populations and their habitats are needed if we are to avoid local tiger extinctions. Our findings suggest that the high level of human activity is one key limiting factor preventing tigers from attaining higher density. Although our study generally presents a somewhat bleak picture for the tiger in central Sumatra, we documented a potentially stable tiger population in Tesso Nilo National Park during this study, where legal protection is discouraging destructive human activities and allowing the tiger and its prey to recover.
Acknowledgements
WWF Indonesia and Networks, the Hurvis Family, the Critical Ecosystem Partnership Fund, the National Fish and Wildlife Foundation/Save the Tiger Fund, USFWS Rhino Tiger Conservation Fund, and Virginia Tech provided technical and financial support for this study. We thank the Indonesian Ministry of Forestry, specifically the Director of Biodiversity Conservation and the managers and staff of the protected areas (especially those from BBKSDA Riau and Balai Taman Nasional Tesso Nilo) for supporting this study. Jim Nichols provided guidance in study design and useful input. This study relied on the support of the WWF team and many field technicians and volunteers, including H. Kurniawan, E. Septayuda, Kusdianto, F. Panjaitan, A. Suprianto, E. Tugiyo, L. Subali and H. Gebog. Koko Yulianto assisted with geographical information systems and mapping. Hariyo T. Wibisono, T. Maddox and J. Sanderson shared their approach to organizing camera trap data. J.E. Hines and Brian Gerber helped with analytical procedures. R. Sugiyanti, Mark Rayan D. and two anonymous reviewers provided useful comments.
Biographical sketches
Sunarto has research interests in the ecology and conservation of threatened wildlife, and is investigating the ecology of tigers in the matrix of forests and plantations in Sumatra. Marcella J. Kelly carries out research on the conservation of large carnivores using non-invasive techniques. Sybille Klenzendorf's research interest is the conservation of large carnivores. Michael R. Vaughan is interested in the ecology and management of large carnivores, including the American black bear. Zulfahmi has been part of the WWF tiger research team for more than 5 years. M.B. Hutajulu is a forest ranger working for Tesso Nilo National Park, with research interests in the management of wildlife. Karmila Parakkasi's research interest is the use of habitat corridors by tigers.







