



Introduction
In the past few years, the knowledge of the geologically youngest and therefore last European crocodylians has improved significantly thanks to the revision of historical collections and the discovery of new remains. On the basis of diagnostic and easily identifiable remains (i.e., skulls, lower jaws, ilia, osteoderms), three genera are recorded: the long-snouted Tomistoma Müller, Reference Müller1846 (or Gavialosuchus Toula and Kail, Reference Toula and Kail1885), the short-snouted Diplocynodon Pomel, Reference Pomel1847, and Crocodylus Laurenti, Reference Laurenti1768. Tomistomine crocodylians had been present in Europe since the Eocene, and their last local representatives come from marginal marine deposits of the late Miocene (Piras et al., Reference Piras, Delfino, Del Favero and Kotsakis2007; Zoboli et al., Reference Zoboli, Sanciu, Pillola and Delfino2019). The endemic alligatoroid Diplocynodon had inhabited Europe since the late Paleocene and went extinct in the middle Miocene (Böhme, Reference Böhme2003; Martin et al., Reference Martin, Smith, de Lapparent de Broin, Escuillié and Delfino2014; Díaz Aráez et al., Reference Díaz Aráez, Delfino, Luján, Fortuny, Bernardini and Alba2017; Luján et al., Reference Luján, Chroust, Čerňanský, Fortuny, Mazuch and Ivanov2019). Interestingly, in the late Miocene the crocodylid Crocodylus appeared in the central Mediterranean sector of southern Europe, where it lingered at least until the Messinian or even the Zanclean (Delfino et al., Reference Delfino, Böhme and Rook2007; Delfino and Rook, Reference Delfino and Rook2008; Delfino and Rossi, Reference Delfino and Rossi2013; Piñero et al., Reference Piñero2017, supplementary data). Ironically, the identification of Crocodylus in the European fossil record overturned the widely accepted opinion that the latter taxon never inhabited Europe, despite the fact that the name Crocodylus (or Crocodilus) was widely used in the nineteenth century for different European species that, once revised, turned out to belong to different genera (Brochu, Reference Brochu2000). Among others, worth mentioning are the cases of Crocodilus affuvelensis Matheron, Reference Matheron1869 from the Cretaceous of France, referred by Martin and Buffetaut (Reference Martin and Buffetaut2008) to the genus Massaliasuchus Martin and Buffetaut, Reference Martin and Buffetaut2008; Crocodilus arduini De Zigno, Reference De Zigno1880 from the Eocene of Italy, now referred to Megadontosuchus Mook, Reference Mook1955 (Piras et al., Reference Piras, Delfino, Del Favero and Kotsakis2007); Crocodylus depressifrons Blainville, Reference Blainville1855 from the Paleocene–Eocene of France, now referred to Asiatosuchus Mook, Reference Mook1940 (even if it clearly does not belong to Crocodylus, there are still questions on its generic attribution; see Delfino and Smith, Reference Delfino and Smith2009; Delfino et al., Reference Delfino, Martin, de Lapparent de Broin and Smith2019); and Crocodylus spenceri Buckland, Reference Buckland1836 from the Eocene of Great Britain, now referred to Kentisuchus Mook, Reference Mook1940 (Brochu, Reference Brochu2007).
The first published evidence for the presence of Crocodylus in Europe consists of few cranial and postcranial remains coming from the latest Miocene–earliest Pliocene (Messinian–Zanclean) fissure fillings of the Gargano promontory in Southern Italy. The presence of undetermined crocodylian remains had already been reported since the paper by Freudenthal (Reference Freudenthal1971) on the vertebrates from the area, but only a revision of the material housed in the collections of the Dipartimento di Scienze della Terra dell'Università degli Studi in Firenze (Italy), of the Naturalis Biodiversity Centre in Leiden (the Netherlands), and of the Universitätsinstitut und Staatssammlung für Paläontologie und historische Geologie in Munich (Germany) led to the identification of Crocodylus on the basis of phylogenetically relevant characters (Delfino et al., Reference Delfino, Böhme and Rook2007).
The following revision (Delfino and Rook, Reference Delfino and Rook2008) of the historical material from the late Miocene (Turolian) lignites of Southern Tuscany (Italy) initially described as Crocodylus bambolii Ristori, Reference Ristori1890, allowed tracking down all the original remains studied by Ristori (Reference Ristori1890). Crocodylian remains from this area (Casteani, Montebamboli, and Ribolla) are housed in three Italian collections: Museo di Storia Naturale dell'Accademia dei Fisiocritici di Siena; Museo di Storia Naturale, Sezione di Geologia e Paleontologia, dell'Università di Firenze; and Museo Geologico Giovanni Capellini in Bologna. Although this abundant material includes partial skulls, fragmentary lower jaws, and postcranial remains, the validity of the species C. bambolii could not be confirmed due to poor preservation, and the type material was therefore referred to cf. Crocodylus sp.
The third, and so far last, ascertained possible evidence of the presence of Crocodylus in Europe comes from the late Miocene (Tortonian) sandstones of Scontrone in Central Italy (material housed in the collections of the Soprintendenza per i Beni Archeologici dell'Abruzzo, Chieti, Italy). The referral to Crocodylus sp. of the first teeth discovered in this locality (Rustioni et al., Reference Rustioni, Mazza, Azzaroli, Boscagli, Cozzini, Di Vito, Masseti and Pisanò1992) was questioned by Kotsakis et al. (Reference Kotsakis, Delfino and Piras2004) and Delfino et al. (Reference Delfino, Böhme and Rook2007), but the subsequent discovery of a lower jaw (along with few other fragmentary, noninformative skeletal elements) confirmed that the crocodylian from Scontrone likely belongs to the genus Crocodylus (cf. Crocodylus sp.; Delfino and Rossi, Reference Delfino and Rossi2013). Worth mentioning is that the latter remains represent the geologically oldest evidence of Crocodylus, being dated to at least 9 Ma (see Delfino and Rossi, Reference Delfino and Rossi2013 and references therein), whereas the oldest remains from Africa date back to about 7 Ma (Brochu and Storrs, Reference Brochu and Storrs2012). Because Europe cannot be considered the center of origin of Crocodylus, it is therefore likely that the first evolutionary steps of this genus were not recorded or are still to be discovered either in Africa or in Asia (Meredith et al., Reference Meredith, Hekkala, Amato and Gatesy2011; Oaks, Reference Oaks2011; Brochu and Storrs, Reference Brochu and Storrs2012; Nicolaï and Matzke, Reference Nicolaï and Matzke2019).
Other European late Miocene–early Pliocene localities yielded remains of crocodylians: Alhaurín el Grande-1 (Guerra-Merchán et al., Reference Guerra-Merchán, Serrano, Ruiz Bustos, Garcés, Insua-Arévalo and García-Aguilar2013); Brisighella (Delfino, Reference Delfino2002); Castro (Alfaro et al., Reference Alfaro, Soria and Ruiz Bustos1995); Crevillente 2 and 15 (Montoya, Reference Montoya1994; Montoya and Alberdi, Reference Montoya and Alberdi1995); Crevillente 4 (Böhme and Ilg, Reference Böhme and Ilg2003); Fiume Santo (Abbazzi et al., Reference Abbazzi, Delfino, Gallai, Trebini and Rook2008) and other Sardinian localities (Zoboli et al., Reference Zoboli, Sanciu, Pillola and Delfino2019); Masía de la Roma 4B (Böhme and Ilg, Reference Böhme and Ilg2003); Moncucco Torinese (Colombero et al., Reference Colombero2017); Plakias (Georgalis et al., Reference Georgalis, Villa, Vlachos and Delfino2016); Puerto de la Cadena (Piñero et al., Reference Piñero2017); and Ribatejo (Böhme and Ilg, Reference Böhme and Ilg2003). However, so far it has not been possible to ascertain or discard the presence of Crocodylus because the materials consist nearly exclusively of very fragmentary remains (in many cases just isolated teeth sometimes referred to Diplocynodon or Tomistoma) that are not diagnostic at genus rank and should be considered as indeterminate crocodylians (Brochu, Reference Brochu2000).
The only other possible evidence for the presence of Crocodylus in Europe comes from the late Miocene of Venta del Moro (VM) in Valencia, Spain, where Crocodylus cf. C. checchiai Maccagno, Reference Maccagno1947 was preliminarily reported by Montoya et al. (Reference Montoya, Morales, Robles, Abella, Benavent, Marín and Ruiz Sánchez2006). Here we describe for the first time the crocodylian material from this locality, and besides reconsidering its identification, we discuss its relevance in the context of the dispersal of Crocodylus across the late Miocene Tethys.
Geological setting
The locality of VM was first reported by Aguirre et al. (Reference Aguirre, Robles, Thaler, López Martínez, Alberdi and Fuentes1973). Located 2 km southeast of the village with the same name (UTM Zone 30S, 642494E, 4370967N), in the Cabriel Basin (Valencia, Spain; Fig. 1), it has yielded an abundant and diverse fossil assemblage of plants (pollen and macroflora; Casas-Gallego et al., Reference Casas-Gallego, Lassaletta, Barrón, Bruch and Montoya2015) and animals (both invertebrates and vertebrates; Montoya et al., Reference Montoya, Morales, Robles, Abella, Benavent, Marín and Ruiz Sánchez2006, Reference Montoya, Morales and Abella2009). The faunal list includes 48 species of mammals, of which nine have VM as their type locality. VM also records the first appearance datum (FAD) in Europe for the ursoid Agriotherium (Morales, Reference Morales1984; Abella et al., Reference Abella, Montoya and Morales2014, Reference Abella, Hontecillas, Valenciano, Montoya, Morales, Pesquero, Alcalá, Bonis and Werdelin2019), the camel Paracamelus (Morales et al., Reference Morales, Soria and Aguirre1980; Morales, Reference Morales1984; Pickford et al., Reference Pickford, Morales and Soria1993, Reference Pickford, Morales and Soria1995; van der Made and Morales, Reference van der Made, Morales, Rössner and Heissig1999), and the murid Paraethomys (Gibert et al., Reference Gibert, Scott, Montoya, Ruiz-Sánchez, Morales, Luque, Abella and Lería2013; Mansino et al., Reference Mansino, Crespo, Montoya and Ruiz-Sánchez2017). For additional information on the faunal assemblage from VM see Morales (Reference Morales1984), Montoya et al. (Reference Montoya, Morales, Robles, Abella, Benavent, Marín and Ruiz Sánchez2006, Reference Montoya, Morales and Abella2009, Reference Montoya, Morales and Abella2011), Pesquero et al. (Reference Pesquero, Alberdi and Montoya2007), Salesa et al. (Reference Salesa, Antón, Turner, Alcalá, Montoya and Morales2010), Alba et al. (Reference Alba, Montoya, Pina, Rook, Abella, Morales and Delson2015), Mansino et al. (Reference Mansino, Ruiz-Sánchez, Freudenthal and Montoya2014, Reference Mansino, García-Alix, Ruiz-Sánchez and Montoya2015a, Reference Mansino, Crespo, Montoya and Ruiz-Sánchez2017, Reference Mansino, Ruiz-Sánchez and Montoya2018), and Crespo et al. (Reference Crespo, Sevilla, Mansino, Montoya and Ruiz-Sánchez2018).
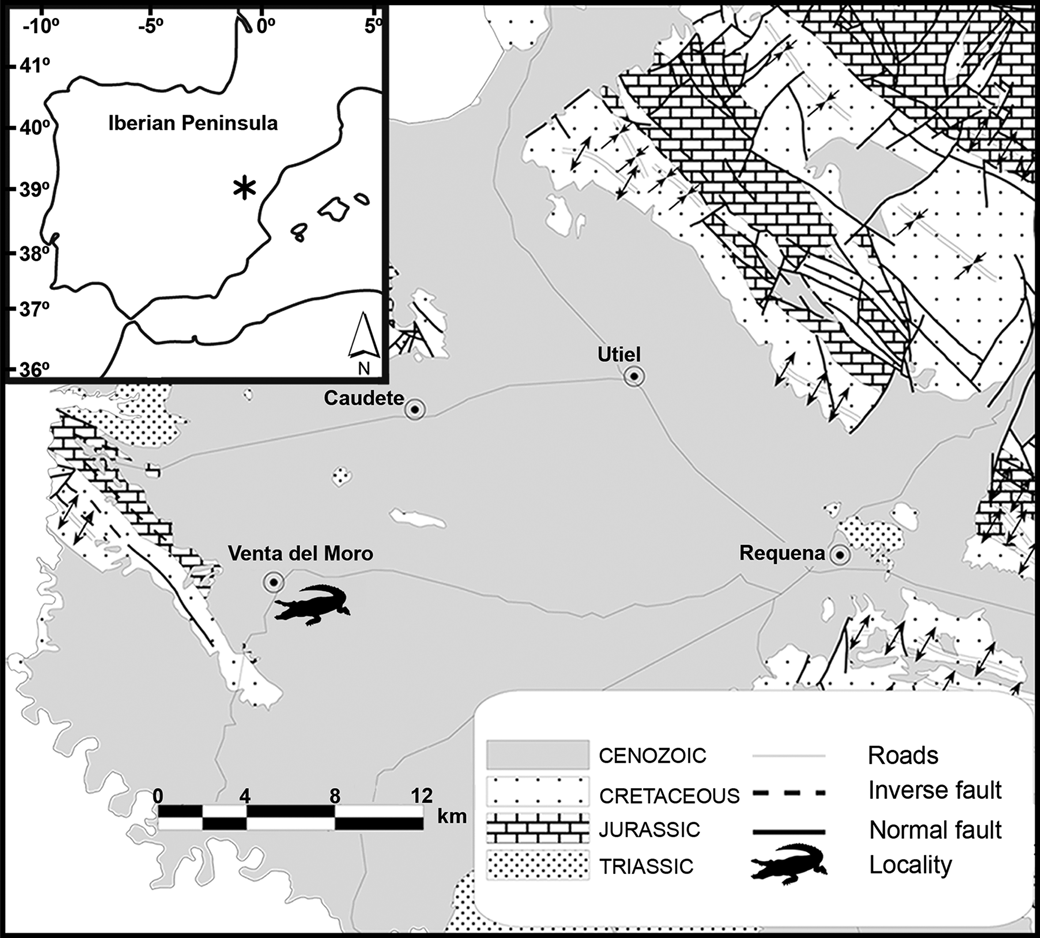
Figure 1. Map of the Iberian Peninsula showing the position of Venta del Moro.
Magnetostratigraphic correlation of VM with chron C3An indicates an estimated age of 6.23 Ma (Gibert et al., Reference Gibert, Scott, Montoya, Ruiz-Sánchez, Morales, Luque, Abella and Lería2013), while the mammal assemblage implies a correlation with MN13 (Mein, Reference Mein, Lindsay, Fahlbush and Mein1990, Reference Mein, Rössner and Heissing1999; De Bruijn et al., Reference De Bruijn, Daams, Daxner-Höck, Falhbusch, Ginsburg, Mein and Morales1992) and, more precisely, the Teruel local biozone M3 (van Dam et al., Reference van Dam, Aziz, Álvarez-Sierra, Hilgen, van den Hoek Ostende, Lourens, Mein, van der Meulen and Peláez-Campomanes2006) within the late Turolian or Ventian mammal age (Morales et al., Reference Morales, Peláez-Campomanes, Abella, Montoya, Gibert, Scott, Cantalapiedra and Sanisidro2013). This time span is characterized by important dispersal events of multiple mammalian taxa into western Europe (Pickford and Morales, Reference Pickford and Morales1994; Agustí et al., Reference Agustí, Garcés and Krijgsman2006; van der Made et al., Reference van der Made, Montoya and Morales2006; Minwer-Bararat et al., Reference Minwer-Barakat, García-Alix, Agustí, Martín Suárez and Freudenthal2009; Gibert et al., Reference Gibert, Scott, Montoya, Ruiz-Sánchez, Morales, Luque, Abella and Lería2013; Mansino et al., Reference Mansino, Ruiz-Sánchez, de Luque, Montoya, Gibert, Morales, Abella, Crespo and Scott2015b; García-Alix et al., Reference García-Alix, Minwer-Barakat, Martín Suárez, Freudenthal, Aguirre and Kaya2016; Minwer-Barakat et al., Reference Minwer-Barakat, Agustí, García-Alix and Martín-Suárez2018), mainly from Asia (camels, canids, colobines, ursoids), but also from Africa (hippos, crocodiles). VM also records other taxa of African affinities, such as the murid rodent Paraethomis meini Michaux, Reference Michaux1969 and the chiropteran Myotis podlesicensis Crespo et al., Reference Crespo, Sevilla, Mansino, Montoya and Ruiz-Sánchez2018 (Mansino et al., Reference Mansino, Crespo, Montoya and Ruiz-Sánchez2017; Crespo et al., Reference Crespo, Sevilla, Mansino, Montoya and Ruiz-Sánchez2018).
Materials and methods
The study presented here is based on all fossil crocodylian remains found in VM (Fig. 1), which were formerly housed at the Museu de Geologia de la Universitat de València, Spain (MGUV) and are currently at the Museu de la Universitat de València d'Història Natural. Pictures of the most relevant specimens were taken using a Canon EXUS 170 digital camera. Measurements were taken with a digital caliper to the nearest 0.1 mm.
Repository and institutional abbreviation
Museu de Geologia de la Universitat de València, Spain (MGUV), Valencia, Spain.
Systematic paleontology
Order Crocodylia Gmelin, Reference Gmelin1789
Family Crocodylidae Laurenti, Reference Laurenti1768
Genus Crocodylus Laurenti, Reference Laurenti1768
Type species
Crocodilus vulgaris Cuvier, Reference Cuvier1807
cf. Crocodylus sp.
Figures 2, 3

Figure 2. Photos and line drawings of crocodylian remains from the late Miocene of Venta del Moro. (1–4) Frontal (MGUV 14472): (1, 2) dorsal view; (3, 4) ventral view. (5–10) Right quadrate sutured with the overlying exoccipital and partial squamosal (MGUV 18828): (5, 6) posterior view; (7, 8) dorsal view; (9, 10) detail of the quadrate condyle in posterior view. (11–14) Right partial quadrate (MGUV 14467): (11, 12) dorsal view; (13, 14) ventral view. (15–18) Fragment of skull table (MGUV 14470) with part of the parietal and the supraoccipital: (15, 16) dorsal view; (17, 18) posterior view. cqg = cranioquadrate groove; exo = exoccipital; f = frontal; fae = foramen aëreum; p = parietal; q = quadrate; sq = squamosal; soc = supraoccipital. Oblique lines denote missing portions or cast.

Figure 3. Photos and line drawings of crocodylian remains from the late Miocene of Venta del Moro. (1–4) Right surangular (MGUV 18835) joined with the corresponding angular MGUV 14466: (1, 2) lateral view; (3, 4) medial view. (5–12) Isolated teeth (MGUV 16002, 16001, 16004, 14504) in labial views. (13–16) Right articular (MGUV 14475): (13, 14) dorsal view; (15, 16) medial view. (17–20) Centra of isolated vertebrae (MGUV 14464-5) in left lateral views. (21, 22) Fragmentary osteoderm (MGUV 14471) in external view. an = angular; art = articular; sur = surangular. Oblique lines denote missing portions or cast.
Description
The only available jugal (MGUV 144699) is very fragmentary, and its only relevant character is the presence of a slender postorbital bar that is inset relative to the lateral surface of the element (it is not clear whether all the bone fragments cataloged under this collection number belong to the same skeletal element).
A well-preserved frontal (MGUV 14472; Fig. 2.1–2.4) clearly lacks any trace of a ‘step’ at the base of the frontal process (however, most of this process is broken off). The frontoparietal suture is deeply concave anteriorly (it is nearly V-shaped as in derived crocodylids), and the supratemporal fenestrae did not reach the frontal. The dorsal surface of the bone is markedly concave, devoid of a midsagittal crest, but ornamented with large pits (Fig. 2.1, 2.2).
The squamosal, partially preserved in MGUV 18828, has a long and relatively slender paroccipital process (Fig. 2.5–2.10). The single exoccipital available from VM (MGUV 18828; Fig. 2.5–2.8) has no trace of a paroccipital boss. The posterolateral edge of this element forms a laminar structure that overhangs the wide opening of the cranioquadrate passage. The foramen vagi is the largest of the foramina of the area, followed by a considerably smaller foramen caroticum posterius (which is placed medioventral to the first) and then by the small foramen for the cranial nerve XII (which is located close to the medial corner of the foramen vagi). The exoccipital preserves the contact surface with the basioccipital and part of the rim of the foramen magnum (Fig. 2.5–2.8).
Information on the morphology of the quadrate is available thanks to MGUV 18828, a right quadrate still sutured with the overlying exoccipital and partial squamosal (Fig. 2.5–2.10), and the isolated right quadrate MGUV 14467 (Fig. 2.11, 2.14). MGUV 18828 is smaller (mediolateral breadth of 27.2 mm) and better preserved than MGUV 14467 (original mediolateral breadth >34 mm). The medial hemicondyle is preserved only in MGUV 18828, in which it is clearly expanded so that it does not significantly slope when seen in posterior view (Fig. 2.5, 2.6, 2.9, 2.10). As for the original opening of the foramen aëreum, none of the two available quadrates shows it, but both preserve a trace revealing its presence very close to the medial edge of the quadrate (Fig. 2.5, 2.6, 2.9–2.14). The medial and dorsomedial surfaces of MGUV 14467 are eroded so that the ventrolateral surface of a large canal can be seen at the medial margin of the quadrate ramus. The posterior termination of this canal originally corresponded to the foramen aëreum (Fig. 2.11–2.14). The medial surface of MGUV 18828 is well preserved close to the medial hemicondyle, but it is broken off in its anterior portion, so that also in this case the ventral surface of the canal corresponding to the foramen aëreum is visible close to the medial edge of the quadrate (Fig. 2.9, 2.10). The squamosal of MGUV 18828 is not completely preserved, but it seems that it did not develop a significant ‘squamosal horn.’
A very small portion of a parietal is preserved in MGUV 14470 (Fig. 2.15–2.18). The dorsal surface is nearly flat and reaches the posteromedial rim of the left supratemporal fenestra, which is slightly raised (Fig. 2.15, 2.16).
The supraoccipital is part of the fragment of the skull table MGUV 14470 and is only very modestly visible in dorsal (Fig. 2.15, 2.16) and posterior (Fig. 2.17, 2.18) views. Its dorsal surface is lowered relative to that of the surrounding parietal (Fig. 2.15, 2.16). In posterior view, it has a vaguely hexagonal shape (Fig. 2.17, 2.18).
The only available ectopterygoid is so fragmentary that it bears no significant information.
The surangular MGUV 18835 and the angular MGUV 14466 clearly belong to the same individual, because they are congruent in terms of size and general shape, and the fine morphology of the preserved portion of the surangular–angular suture matches very well on both elements (Fig. 3.1, 3.4). They both belong to a rather large specimen, the length of their preserved parts being 19.0 and 18.4 mm, respectively. The surangular terminates anteriorly with a long dorsal process overhanging a small ventral process; the posterodorsal tip seems to be complete (Fig. 3.1, 3.4). The external mandibular fenestra was quite large, but not enough to see the foramen intermandibularis caudalis in lateral view (Fig. 3.1, 3.2). The angular–surangular suture reaches the external mandibular fenestra at its posterior angle. The lateral surface of both the surangular and the angular is ornamented with large and deep pits separated by evident ridges (Fig. 3.1, 3.2).
The partially preserved right articular MGUV 14475 (Fig. 3.13–3.16) does not show the position of the foramen aëreum because of poor preservation, but the lamina lying against the surangular is relatively well developed. The articular–surangular suture within the glenoid fossa is not linear, but bowed (Fig. 3.13–3.14). The retroarticular process is long, slender, and probably directed in a posterodorsal direction (Fig. 3.13–3.16).
Isolated teeth (Fig. 3.5–3.12) are numerous, and their sizes vary from a few millimeters (e.g., 6.5 mm in MGUV 25924; also very small are MGUV 14440, 16002) to 39.0 mm (MGUV 16006). The shape ranges from elongate, slender, and pointed as in MGUV 16002 (Fig. 3.5, 3.6), 24317, and 16001 (Fig. 3.7, 3.8) to more massive and broadly conical as in MGUV 16006, 25993, and 14504 (Fig. 3.11, 3.12) to short, proportionally massive, and apically blunt as in MGUV 14443, 16004 (Fig. 3.9, 3.10), and 18932. The largest tooth (MGUV 16006) only partially preserves the root (of the 39.0 mm of total length, 25 mm correspond to the crown) and shows characters common to most of the other teeth: the crown is provided with two mesiodistal carinae, which are not serrated; the base of the crown is slightly elliptical (17.0 mm long and 15.3 mm wide in MGUV 16006); and there is no constriction between the crown and the root. MGUV 14436 is an exception because it has a modest constriction at the base of the crown. On some teeth, the surface has several longitudinal ridges (about 10 on the lingual surface of MGUV 14436). Both shed teeth, with a totally reabsorbed root (MGUV 14443, 16004: Fig. 3.9, 3.10), and teeth still preserving the root (MGUV 16006) are present.
Three isolated vertebrae (MGUV 14463, 14464, and 14465) are represented by an incomplete, procoelous centrum, whose estimated original length was about 25–30 mm (Fig. 3.17–3.20). The centra were separated from the neural arch at the level of the neurocentral synchondrosis in at least MGUV 14464 and 14465. A well-developed hypapophysis is present in MGUV 14463; MGUV 14465 does not show any hypapophysis, and its ventral surface is wide and flat (the other centrum is ventrally eroded, but probably the hypapophysis was not present; Fig. 3.17, 3.18).
The only clearly identifiable but fragmentary osteoderm is MGUV 14471. It is characterized by very deep and relatively large, elongated pits separated by rounded ridges (Fig. 3.21, 3.22). Only two original edges are preserved: one is smooth and straight and thins gradually; the other is thick and hosts a well-developed suture. The entire osteoderm was probably rather large, the preserved portion being 31 mm long and 25 mm wide (Fig. 3.21). MGUV 14474 is a possible second, very fragmentary, osteoderm.
Materials
MGUV 14469: right jugal; MGUV 14472: frontal; MGUV 14467: right quadrate; MGUV 18828: right squamosal, exoccipital, and quadrate; MGUV 14470: fragmentary parietal and supraoccipital; MGUV 14816: ectopterygoid; MGUV 18835: right surangular; MGUV 14466: right angular; MGUV 14475: right articular; MGUV 14436-62, 14468, 14490, 14502, 14607, 15853, 16001–16006, 18932, 24317, 25923–25926, 25993: isolated teeth; MGUV 14463–14465: isolated vertebrae; MGUV 14471: osteoderm; MGUV 14473: unidentified fragment of skull bone; MGUV 14474: fragmentary osteoderm.
Remarks
The crocodylian remains from VM, despite being very fragmentary, show characters that clearly allow us to exclude their referral to either Diplocynodon or tomistomines—the only crocodylians besides Crocodylus known to have inhabited the Mediterranean Basin during the late Neogene. The described remains do not belong to Diplocynodon because the latter, as all alligatoroids, is characterized by a dorsal opening of the foramen aëreum, whereas both MGUV 14467 and MGUV 18828, despite not showing the original position of the foramen, clearly indicate that it was placed close to the dorsomedial edge of the quadrate. Moreover, the quadrate medial hemicondyle of MGUV 18828 is clearly dorsally expanded, instead of sloping with a notch corresponding to the foramen aëreum as in Diplocynodon. In addition, the frontal MGUV 14472 does not show the step at the base of the frontal process that characterizes, among others, Diplocynodon, and at the same time, it shows a deeply concave frontoparietal suture instead of being rectilinear (or nearly so) as in tomistomines (see character 151 in Brochu and Storrs, Reference Brochu and Storrs2012). Even if highly fragmentary, the preserved morphology of the osteoderm MGUV 14471 does not fit with either that of tomistomines (characterized by relatively few and comparatively very large pits; see Zoboli et al., Reference Zoboli, Sanciu, Pillola and Delfino2019, fig. 8) or that of Diplocynodon (e.g., see Delfino and Smith, Reference Delfino and Smith2012; Martin et al., Reference Martin, Smith, de Lapparent de Broin, Escuillié and Delfino2014). Moreover, the development of the lamina of the articular lying against the surangular (among others, see Brochu and Storrs, Reference Brochu and Storrs2012) also allows us to rule out the extant African crocodylid Mecistops, which was never recovered in the North African and European fossil record. The morphology of the crocodylian remains from VM is congruent with that of Crocodylus, although it is not possible to detect any of the apomorphies of this genus (Brochu, Reference Brochu2000) among the currently available material. Consequently the morphology of the remains from VM does not permit us to confirm their referral to C. checchiai, a species originally described by Maccagno (Reference Maccagno1947, Reference Maccagno1952) on the basis of well-preserved skulls from the latest Miocene locality of As Sahabi (Libya). Further remains of C. checchiai from As Sahabi were later described by Delfino (Reference Delfino, Boaz, El-Arnauti, Pavlakis and Salem2008), but it is thanks to the new material from the Turkana Basin (Kenya) recently described by Brochu and Storrs (Reference Brochu and Storrs2012) that the phylogenetic relationships of this taxon have been further clarified, confirming its referral to Crocodylus. New evidence (Delfino et al., Reference Delfino, Iurino, Mercurio, Piras, Rook and Sardella2020) based on the revision of material originally described by Maccagno strengthens the referral of C. checchiai to Crocodylus by placing it at the base of the American extinct and extant species and, supporting and African American dispersal, testify for the dispersal abilities of the late Miocene crocodylids that inhabited the Mediterranean area. Other late Miocene crocodylians from Northern Africa have not been described and identified in detail, but worth mentioning are the materials from Tizi N'Tadderht (Morocco; Zouhri et al., Reference Zouhri, Geraads, El Boughabi and El Harfi2012) and Djebel Krechem el Artsouma (Tunisia; Geraads, Reference Geraads1989). The Moroccan locality yielded a skull referred to Crocodylus cf. C. niloticus Laurenti, Reference Laurenti1768 whereas the Tunisian material has been referred to Crocodylus cf. C. checchiai but should be revised now that the morphology of the topotypic remains is better known than before (Delfino et al., Reference Delfino, Iurino, Mercurio, Piras, Rook and Sardella2020). The African C. checchiai is characterized by a set of features (among others, the mid-rostral boss) that are not accessible among the material from VM. Even if C. checchiai is so far the only well-known short-snouted crocodylid species that inhabited the Mediterranean Basin during the late Miocene, it does not seem advisable to refer the material from VM to this species on the basis of geography alone (especially considering that the former has only been unambiguously identified in Libya and Kenya so far). Thus, based on its preserved morphology, biogeography, and chronology, the VM crocodylian is here referred to cf. Crocodylus sp., an assignment that could be confirmed by the retrieval of additional and more-informative material.
Discussion
The geographic area of origin of the genus Crocodylus is currently unresolved by the fossil record and could be located in either Asia or Africa (Oaks, Reference Oaks2011; Brochu and Storrs, Reference Brochu and Storrs2012). This lack of knowledge, besides the poor taxonomic resolution provided by the VM material, precludes a firm biogeographical interpretation of the VM crocodylian. According to Pickford and Morales (Reference Pickford and Morales1994), MN13 is a phase of northward latitudinal shift of the boundary zone between tropical and boreal biogeographic realms. This shift could explain the presence of Crocodylus at Gargano and its possible presence at VM (both MN13) but does not explain its possible presence at Montebamboli (MN12) and, even less, Scontrone (MN11), which are older. Delfino and Rossi (Reference Delfino and Rossi2013) proposed that Crocodylus could have repeatedly dispersed from Africa to Europe across the Mediterranean Basin. As the latter was a small sea scattered with islands, especially during the late Neogene, it would not have represented an important biogeographic barrier to members of this genus, which is well known for being able to survive in saltwater (Leslie and Taplin, Reference Leslie, Taplin, Grigg, Seebacher and Franklin2001) and includes at least some extant species that are able to quickly disperse across saltwater (Campbell et al., Reference Campbell, Watts, Sullivan, Read, Choukroun, Irwin and Franklin2010). The fact that the paleo-Mediterranean Basin could be easily crossed is further corroborated by the slightly earlier (6.3 Ma) FAD of the hippopotamid Hexaprotodon in Spain, which is thought to have dispersed from Africa by swimming through the Rifian Corridor before the subsequent intercontinental faunal exchanges between Iberia and North Africa at 6.2 Ma (Gibert et al., Reference Gibert, Scott, Montoya, Ruiz-Sánchez, Morales, Luque, Abella and Lería2013, and references therein). The evolution of endemic hippopotamus species in fossil islands has been classically explained by their purported good swimming abilities (e.g., Sondaar, Reference Sondaar, Hecht, Goody and Hecht1977). Although this has recently been challenged (Mazza, Reference Mazza2014, Reference Mazza2015) on the grounds that Hippopotamus amphibius Linnaeus, Reference Linnaeus1758 adults are too dense to float on freshwater (Coughlin and Fish, Reference Coughlin and Fish2009), this does not apply to juvenile individuals and, hence, must not necessarily be the case for extinct hippopotamid species, particularly on saltwater due to increased buoyancy (van der Geer et al., Reference van der Geer, Anastasakis and Lyras2015). There is no geological evidence for the land bridges hypothesized by Mazza (Reference Mazza2014, Reference Mazza2015) to explain the dispersal of hippopotami to islands such as Cyprus, Madagascar, and Crete (van der Geer et al., Reference van der Geer, Anastasakis and Lyras2015). In the case of the Iberian Peninsula during the latest Miocene, the earlier dispersal of hippopotami as compared with terrestrial mammals argues in favor of dispersal through seawater, once the Betic Seaway was already closed and the Riftian corridor very restricted (Gibert et al., Reference Gibert, Scott, Montoya, Ruiz-Sánchez, Morales, Luque, Abella and Lería2013). It is therefore interesting that material potentially referable to Crocodylus is recorded in the Iberian Peninsula because, so far, the evidence for the presence of this taxon in the northern Mediterranean area was limited to three localities in the Apennine Peninsula.
Conclusions
The crocodylian fossil remains from the late Miocene (6.23 Ma) of VM are too fragmentary to assess their previous assignment to C. checchiai but allow us to rule out an assigment to either Diplocynodon or tomistomines, the two other crocodylian clades previously recorded from the late Miocene of Europe. Therefore, a tentative assignment to cf. Crocodylus is warranted on morphological grounds. The identification of at least two individuals (as testified by the two right quadrates) indicates that the possible presence of Crocodylus at VM cannot be due to the occasional dispersal of just one individual but suggests that a population could have been present in the area. All the European localities with late Miocene crocodylians, VM included, were at that time close to the northern shores of the Mediterranean and therefore easily reachable by specimens dispersing via seawater. The fact that C. checchiai from Libya is phylogenetically intermediate between the African C. niloticus and the American Crocodylus, coupled with the fact that C. checchiai predates the appearance of this genus in America, supports good dispersal abilities of the late Miocene African crocodylids. Therefore, like the hippopotamid Hexaprotodon, the genus Crocodylus could have dispersed by seawater from Africa into Europe somewhat before the establishment of a clear land connection between these two continents. However, it is quite frustrating that, so far, all these late Miocene crocodylians from Europe are represented by isolated teeth, osteoderms, or highly fragmentary remains that hinder a precise specific identification and therefore the retrieval of precise information unveiling the biogeographic history of the last European crocodylians.
Acknowledgments
Fieldwork in Venta del Moro from 1995 to 2012 was supported by the Conselleria de Cultura of the Valencian Government and by the Conselleria d'Empresa, Universitat i Ciència of the Generalitat Valenciana (project number GV06/304), with the permission of the Ministerio de Fomento and the Company ADIF, of the Spanish Government. This research has been funded by the Ministerio de Ciencia e Innovación (PGC2018-094122-B-100), the Agencia Estatal de Investigación (CGL2016-76431-P; AEI/FEDER, UE), the Research Groups CSIC 64 1538 and CAM-UCM 910607, and the Generalitat de Catalunya (CERCA Programme, consolidated research group 2017 SGR 116 GRC to D.M.A., and Beatriu de Pinós contract 2017 BP 00223 from AGAUR to J.A.). E.T. contributed to this project while funded by the European Union's FP7 program under the Marie Skłodowska-Curie grant agreement no. 609402 - 2020 researchers: Train to Move (T2M). À.H.L. is financially supported through a postdoctoral grant from Operational Programme Research, Development and Education—Project ‘Postdoc@MUNI’ (no. CZ.02.2.69/0.0/0.0/16_027/0008360). M.D. acknowledges also the University of Turin (Fondi di Ateneo 2018 and 2019). We further thank J. Martin for providing relevant literature. The latter, along with C.A. Brochu, reviewed the manuscript and provided useful suggestions.



