



Significant Findings
-
Lowered CD4+CB2+ T cells are associated with increased immune-inflammatory response system (IRS) in major depressive disorder (MDD)
-
Lowered CD3+CB2+% and increased CD20+CB2+% predict the severity of depression and suicidal behaviours.
-
Lowered CD3+/CD4+CB2+ may impact the immune homoeostatic processes, leading to enhanced IRS in MDD
-
Increased CD20+CB2+% may activate T-independent humoral immunity and enhance IRS responses.
-
Depletion of CB1-bearing T regulatory and CB2-bearing T and T helper cells and increased CB2+-bearing B cells are new drug targets in MDD.
Limitations
-
The findings deserve replication in other countries and cultures.
-
Future research should examine CB2-bearing macrophages, dendritic cells and natural killer cells in MDD.
Introduction
There is now evidence that major depressive disorder (MDD) is a neuroimmune disorder characterised by multiple immune system dysfunctions that culminate in an increase in neuronal and astroglial toxicity (Maes, Reference Maes1995; Reference Maes, Rachayon, Jirakran, Sodsai, Klinchanhom, Gałecki, Sughondhabirom and Basta-Kaim2022; Maes and Carvalho, Reference Maes and Carvalho2018; Al-Hakeim et al., Reference Al-Hakeim, Al-Naqeeb, Almulla and Maes2023). Initial studies demonstrated that MDD is associated with elevated levels of pro-inflammatory cytokines of the M1 macrophage lineage, such as interleukin (IL)-1β, soluble IL-1 receptor antagonist (sIL-1RA), IL-6, IL-8 (or CXCL8), and tumour necrosis factor (TNF)-α, and the T helper (Th)-1 lineage, such as IL-2, IL-2 receptor (sIL-2R), and interferon (IFN)-γ (Maes et al., Reference Maes, Bosmans, Suy, Vandervorst, De Jonckheere and Raus1990; Reference Maes, Bosmans, Suy, Vandervorst, DeJonckheere and Raus1991; Reference Maes, Scharpé, Meltzer, Okayli, Bosmans, D’Hondt, Vanden Bossche and Cosyns1994; Reference Maes, Vandoolaeghe, Ranjan, Bosmans, Bergmans and Desnyder1995b; Mikova et al., Reference Mikova, Yakimova, Bosmans, Kenis and Maes2001). Additional research conducted in the laboratories of Maes et al. demonstrated elevated levels of complement factors and positive acute phase proteins, such as haptoglobin, and decreased levels of negative acute phase proteins, such as albumin, in MDD, indicating that MDD is accompanied by a chronic mild inflammatory (‘acute phase’) response (Maes, Reference Maes1993). The cytokine, macrophage-T lymphocyte and immune-inflammatory response system (IRS) theories of MDD summarised these findings (Maes, Reference Maes1995; Maes et al., Reference Maes, Smith and Scharpe1995a). Recent systematic reviews and meta-analyses corroborate that MDD is associated with elevated serum levels of M1 macrophage and Th1 cytokines and immune activation (Kohler et al., Reference Köhler, Freitas, Maes, de Andrade, Liu, Fernandes, Stubbs, Solmi, Veronese, Herrmann, Raison, Miller, Lanctôt and Carvalho2017; Gasparini et al., Reference Gasparini, Callegari, Lucca, Bellini, Caselli and Ielmini2022; Zhang et al., Reference Zhang, Wang, Ye, Zou, Chen, Wang and Zou2023; Foley et al., Reference Foley, Parkinson, Mitchell, Turner and Khandaker2023; Mac Giollabhui et al., Reference Mac Giollabhui, Ng, Ellman and Alloy2021).
Recent research indicates that MDD is characterised not only by activation of M1, Th-1 and Th-17 activation but also the compensatory immunoregulatory system (CIRS), which prevents hyperinflammation and endeavours for immune homoeostasis (Maes et al., Reference Maes, Berk, Goehler, Song, Anderson, Gałecki and Leonard2012; Maes and Carvalho, Reference Maes and Carvalho2018). Th-2 (producing IL-4) and T regulatory (Treg, producing IL-10) phenotypes are major CIRS components that are activated in MDD (Maes et al., Reference Maes, Rachayon, Jirakran, Sodsai, Klinchanhom, Gałecki, Sughondhabirom and Basta-Kaim2022). In MDD, T cell growth factors such as IL-4, IL-9, IL-12 and GM-CSF contribute to the simultaneous activation of both IRS and CIRS components (Maes et al., Reference Maes, Rachayon, Jirakran, Sodsai, Klinchanhom, Gałecki, Sughondhabirom and Basta-Kaim2022). Importantly, certain IRS and CIRS cytokines (IL-1β, IL-2, IL-4, IL-6, TNF-α and IFN-γ) and chemokines (CCL1, CCL2, CCL11, CXCL5, CXCL8 and CXCL10) have neurotoxic effects and could, therefore, contribute to the neuronal and astroglial projection toxicity in MDD (Maes et al., Reference Maes, Rachayon, Jirakran, Sodsai, Klinchanhom, Gałecki, Sughondhabirom and Basta-Kaim2022; Al-Hakeim et al., Reference Al-Hakeim, Al-Naqeeb, Almulla and Maes2023).
Early machine learning and flow cytometry studies to quantify the percentage and number of peripheral blood mononuclear cells (PBMC) revealed IRS activation, particularly T cell activation, in patients with MDD with an increased expression of CD4+, CD45+, CD25+, Ig+ and HLA-DR + on PBMC (Maes et al., Reference Maes, Bosmans, Suy, Vandervorst, De Jonckheere and Raus1990; Reference Maes, Lambrechts, Bosmans, Jacobs, Suy, Vandervorst, de Jonckheere, Minner and Raus1992a; Reference Maes, Stevens, DeClerck, Bridts, Peeters, Schotte and Cosyns1992b; Reference Maes, Stevens, Declerck, Bridts, Peeters, Schotte and Cosyns1993). With a specificity of 92%, machine learning revealed that up to 64% of MDD patients exhibited elevated expression of CD7+CD25+ and CD2+HLA-DR + cells (Maes et al., Reference Maes, Stevens, Declerck, Bridts, Peeters, Schotte and Cosyns1993). Treg cells are also involved in the pathophysiology of MDD. For example, MDD patients have lower Treg levels than healthy controls and in animal models lowered Treg function is associated with depression-like behaviours (Grosse et al., Reference Grosse, Hoogenboezem, Ambrée, Bellingrath, Jörgens, de Wit, Wijkhuijs, Arolt and Drexhage2016; Kim et al., Reference Kim, Lee, Lee, Oh, Shin, Shim and Bae2012). A decrease in T effector cells and an increase in Treg expression characterise the symptomatic remission phase of a major depressive episode of bipolar disorder (BD) (Maes et al., Reference Maes, Nani, Noto, Rizzo, Hayashi and Brietzke2021). Antidepressant-treated depressed patients had a greater number of CD4+CD25+ and CD4+CD25+FoxP3+Treg than untreated depressed patients (Mohd Ashari et al., Reference Mohd Ashari, Mohamed Sanusi, Mohd Yasin, Che Hussin, Wong and Shafei2019). Treg cells, such as CD4+CD25+FoxP3+, CD4+CD25+FoxP3+CD152+ and CD4+CD25+FoxP3+GARP+, are essential for immune tolerance and homoeostasis, the suppression of T cell activation and immune proliferation (Maes et al., Reference Maes, Nani, Noto, Rizzo, Hayashi and Brietzke2021).
B cells and monocytes express higher levels of CB2 receptors than CD4+ (T helper) and CD8+ (T cytotoxic) T lymphocytes (Graham et al., Reference Graham, Angel, Schwarcz, Dunbar and Glass2010). Dendritic cells and macrophages may release endogenous cannabinoids in response to inflammatory stimuli, whereas activating CB2 receptors has profound anti-inflammatory effects by inhibiting T cell activation, M1 macrophage functions, immune cell proliferation and differentiation, and inducing apoptosis in both T and B cells (Klein et al., Reference Klein, Newton, Larsen, Chou, Perkins, Lu, Nong and Friedman2004; Ehrhart et al., Reference Ehrhart, Obregon, Mori, Hou, Sun, Bai, Klein, Fernandez, Tan and Shytle2005; Turcotte et al., Reference Turcotte, Blanchet, Laviolette and Flamand2016; Komorowska-Muller et al., Reference Komorowska-Müller and Schmöle2020; Morcuende et al., Reference Morcuende, García-Gutiérrez, Tambaro, Nieto, Manzanares and Femenia2022; Csoka et al., Reference Csóka, Németh, Mukhopadhyay, Spolarics, Rajesh, Federici, Deitch, Bátkai, Pacher and Haskó2009; Lombard et al., Reference Lombard, Nagarkatti and Nagarkatti2007; Rieder et al., Reference Rieder, Chauhan, Singh, Nagarkatti and Nagarkatti2010; Maresz et al., Reference Maresz, Pryce, Ponomarev, Marsicano, Croxford, Shriver, Ledent, Cheng, Carrier, Mann, Giovannoni, Pertwee, Yamamura, Buckley, Hillard, Lutz, Baker and Dittel2007). When CB2 is activated, M1 macrophages acquire a M2 phenotype and produce more IL-10, an important CIRS cytokine (Klein et al., Reference Klein, Newton and Friedman1998; Correa et al., Reference Correa, Mestre, Docagne and Guaza2005). Transgenic mice overexpressing CB2 (CB2xP) show reduced depressive-like behaviours and are resistant to acute anxiogenic stimuli (Garcia-Gutierrez et al., Reference García-Gutiérrez, Pérez-Ortiz, Gutiérrez-Adán and Manzanares2023).
CB2 is crucial for the homing, development and maintenance of marginal zone (MZ) B cells and enhances T-cell-independent humoral responses of natural IgM (Basu et al., Reference Basu, Ray and Dittel2011; Reference Basu, Ray and Dittel2013; Nolte et al., Reference Nolte, Arens, Kraus, van Oers, Kraal, van Lier and Mebius2004). These IgM are crucial as the first line of innate immune defence against pathogens, including commensal microorganisms (Zouali and Richard, Reference Zouali and Richard2011; Morris et al., Reference Morris, Puri, Olive, Carvalho, Berk and Maes2019). This is significant because MDD patients have elevated IgM antibody levels against a variety of endogenous antigens and lipopolysaccharide (LPS) of gut commensal bacteria as well (Maes et al., Reference Maes, Mihaylova, Kubera, Leunis and Geffard2011; 2013; Simeonova et al., Reference Simeonova, Stoyanov, Leunis, Murdjeva and Maes2021). In this respect, there are some data that CB2 may contribute to bacterial translocation via increased production of IL-10 (Csoka et al., Reference Csóka, Németh, Mukhopadhyay, Spolarics, Rajesh, Federici, Deitch, Bátkai, Pacher and Haskó2009). CB1 is also expressed on immune cells and, like CB2, and also in conjunction with CB2, displays immunosuppressive effects, such as suppression of Th-1 functions (Kaplan, Reference Kaplan2013). However, there is no data on the number of CB2/CB1-bearing CD3+ (T), CD4+, CD8+, CD20+ (B) and FoxP3+ cells in patients with MDD compared to healthy controls.
Cannabidiol (CBD), a phytocannabinoid extracted from the cannabis plant, is a potential antidepressant (Atalay et al., Reference Atalay, Jarocka-Karpowicz and Skrzydlewska2019). There is evidence that CBD may improve clinical depression and anxiety (Martin et al., Reference Martin, Strickland, Schlienz, Munson, Jackson, Bonn-Miller and Vandrey2021), although not all studies show positive results (Hasbi et al., Reference Hasbi, Madras and George2023). Some, but not all, animal models of depression show that cannabinoids have antidepressive effects (Hen-Shoval et al., Reference Hen-Shoval, Amar, Shbiro, Smoum, Haj, Mechoulam, Zalsman, Weller and Shoval2018; Hasbi et al., Reference Hasbi, Madras and George2023). CBD has a lower affinity for CB2 receptors than anandamide (AEA), 2-arachidonoylglycerol (2-AG) and delta-9-tetrahydrocannabinol (THC) (Pertwee, Reference Pertwee2008; Raich et al., Reference Raïch, Rivas-Santisteban, Lillo, Lillo, Reyes-Resina, Nadal, Ferreiro-Vera, de Medina, Majellaro, Sotelo, Navarro and Franco2021) but possesses anti-inflammatory and antioxidative properties, such as inhibiting the production of pro-inflammatory cytokines and reactive oxygen species (Atalay et al., Reference Atalay, Jarocka-Karpowicz and Skrzydlewska2019). CBD is a partial agonist at the CB2 receptor that modifies the efficacy of cannabinoid ligands, suggesting that CBD’s anti-inflammatory and antidepressant effects may be related to their effects on CB2 (An et al., Reference An, Peigneur, Hendrickx and Tytgat2020). However, there is no data indicating whether CBD may affect the number or percentage of CB2/CB1-bearing CD3+, CD4+, CD8+, CD20+ and FoxP3+ cells.
In light of this, the current study examined the number of CB2/CB1-containing CD3+, CD4+, CD8+, CD20+ and FoxP3+CB1+ cells in patients with MDD compared to healthy controls, as well as the in vitro effects of CBD on these cell populations.
Methodology
Participants
We recruited outpatients with MDD admitted to the psychiatry department of the Chulalongkorn Hospital in Bangkok, Thailand, for this study. Healthy controls were recruited from the same geographic region (Bangkok, Thailand) as the patients via posters and word of mouth. The inclusion criteria were patients between the ages of 18 and 65 years who spoke Thai, had been diagnosed with MDD by a senior psychiatrist using DSM-5 criteria and had a Hamilton Depression Rating Scale (HAM-D) score of at least 17. Excluded were healthy participants with a diagnosis of any axis 1 DSM-5 disorder or a positive family history of MDD or BD. Patients with BD, schizophrenia, substance use disorders, schizoaffective disorders, psycho-organic disorders, obsessive-compulsive disorder and post-traumatic stress disorder were excluded from the study. Exclusion criteria for both patients and controls were: neuroinflammatory and neurodegenerative disorders such as multiple sclerosis, Alzheimer’s disease, epilepsy, or Parkinson’s disease; (auto)immune illnesses such as inflammatory bowel disease, chronic obstructive pulmonary disease, cancer, diabetes type 1, asthma, psoriasis, Long COVID and COVID-19 infection (lifelong) or rheumatoid arthritis; and allergic or inflammatory responses in the past 3 months. Participants who took anti-inflammatory medications (NSAIDs or steroids) within 1 month of the study, therapeutic doses of antioxidant or omega-3 polyunsaturated fatty acid supplements within 3 months prior to the study or those who had ever received immunomodulatory drugs were also excluded. Neither lactating women nor pregnant women were allowed to participate.
Some patients were taking psychotropic medications, such as sertraline (18 patients), other antidepressants (8 patients, such as fluoxetine, venlafaxine, escitalopram, bupropion and mirtazapine), benzodiazepines (22 patients), atypical antipsychotics (14 patients) and mood stabilisers (4 patients). In the statistical analysis, the possible effects of these pharmacological variables were taken into consideration.
Before participating in the investigation, all subjects and controls provided written consent. The research was conducted in accordance with both international and Thai privacy and ethics regulations. The Institutional Review Board of the Faculty of Medicine at Chulalongkorn University in Bangkok, Thailand, approved the study (#528/63) in accordance with international guidelines for the protection of human research subjects, including the Declaration of Helsinki, The Belmont Report, CIOMS Guideline and the International Conference on Harmonization in Good Clinical Practice (ICH-GCP).
Clinical measurements
A well-trained research assistant conducted semi-structured interviews to collect demographic data, including age, gender, medical and psychiatric history. An experienced psychiatrist administered the 17-item HAMD to determine the severity of depression (Hamilton, Reference Hamilton1960). The Mini-International Neuropsychiatric Interview (M.I.N.I.) was used to diagnose axis-1 psychiatric illness (Kittirattanapaiboon and Khamwongpin, Reference Kittirattanapaiboon and Khamwongpin2005). The Thai state version of the State-Trait Anxiety Inventory (STAI) was used to assess the level of self-rated state anxiety (Spielberger, Reference Spielberger1983). Suicidal ideation and behaviour were evaluated using the lifeline version of the Columbia-Suicide Severity Rating Scale (C-SSRS) (Posner et al., Reference Posner, Brown, Stanley, Brent, Yershova, Oquendo, Currier, Melvin, Greenhill, Shen and Mann2011). Recent suicidal behaviours were computed as the first principal component (PC) (labelled ‘Current_SB’) extracted from nine C-SSRS items: ‘wish to be dead, non-specific active suicidal thoughts, active suicidal ideation with any methods, active suicidal ideation with some intent to act, active suicidal ideation with a specific plan/intent, frequency and duration of suicidal ideation, actual attempts, and the total number of actual attempts’. This first PC extracted from the nine SB items possessed satisfactory psychometric properties (Maes et al., Reference Maes, Rachayon, Jirakran, Sodsai, Klinchanhom, Gałecki, Sughondhabirom and Basta-Kaim2022). The ‘phenome of depression’ was calculated by extracting the first PC from the total HAMD, STAI and Current_SB scores (Maes et al., Reference Maes, Rachayon, Jirakran, Sodsai, Klinchanhom, Gałecki, Sughondhabirom and Basta-Kaim2022). This PC exhibited excellent psychometric properties and loadings > 0.90 on all indicators. Body mass index (BMI) was calculated by dividing body weight (in kilograms) by length squared (in metres squared). Using DSM-5 criteria, a tobacco use disorder (TUD) was diagnosed.
Assays
Fasting blood samples of 20 mL were collected between 8 and 10 a.m. in BD Vacutainer® EDTA (10 mL) and BD Vacutainer® SST™ (5 mL) tubes (BD Biosciences, Franklin Lakes, NJ, USA). To obtain serum, blood was allowed to clot at room temperature for 30 minutes. The tubes were spun at 1100 ×g for 10 minutes at 4°C, and the resulting serum or plasma was stored at -80°C. PBMCs were separated through density gradient centrifugation (30 min at 900 × g) using Ficoll® Paque Plus (GE Healthcare Life Sciences, Pittsburgh, PA, USA). Cell count and viability were checked using hemocytometer and trypan blue, 0.4% solution, pH 7.2–7.3 (Sigma-Aldrich Corporation, St Louis, MO, USA). The number of total and blue staining cells were counted, ensuring that more than 95% of cells were viable for all conditions.
To evaluate PBMC stimulation, the 96-well plates were coated with 5 µg/mL of the anti-human CD3 antibody (OKT3, from eBioscience), overnight. PBMCs, 3 × 105 cells each, were added to each well, along with 5 μg/mL of the anti-human CD28 antibody (CD28.2, eBioscience). The cells were grown in a culture medium of RPMI 1640 with L-glutamine that was supplemented with 10% foetal bovine serum and 1% penicillin-streptomycin solution (Gibco Life Technologies, Carlsbad, CA, USA). The cells were allowed to grow for 3 days at 37° C in an environment with 5% CO2. A negative control was conducted by growing PBMCs without anti-CD3 and anti-CD28 antibodies for the same period. After 3 days, the immunophenotypes of CD20, CD3, CD4, CD8 and Treg cells as well as CB2/CB1 receptors were determined through flow cytometry. Thus, the specimens can be divided into two conditions: unstimulated and stimulated with anti-CD3 and anti-CD28, after being incubated for 72 hours. The effects of three CBD concentrations on the stimulated immune cells were examined using CBD 0.1 µg/ml (CBDlow), 1 µg/mL (CBDmedium) and 10 µg/mL (CBDhigh) (Rachayon et al., Reference Rachayon, Jirakran, Sodsai, Klinchanhom, Sughondhabirom, Plaimas, Suratanee and Maes2022). The natural CBD (99.89% isolate) stock preparation was manufactured by Love Hemp Ltd, United Kingdom (batch 8406), and the Medical Cannabis Research Institute, College of Pharmacy, RSU, Bangkok, Thailand, guaranteed its quality. Concentrations between 0.1 and 1.0 µg/mL are therapeutic concentrations of CBD, whereas levels as high as 5–20 µg/mL may be obtained when combining oral CBD with dietary lipids (Rachayon et al., Reference Rachayon, Jirakran, Sodsai, Klinchanhom, Sughondhabirom, Plaimas, Suratanee and Maes2022).
To study lymphocyte phenotypes, 3 × 105 PBMCs were labelled with a combination of monoclonal antibodies for 30 minutes, including CD3-PEcy7, CD4-APCcy7, CD8-APC, CD20-FITC and CB2-AF700 antibodies (Biolegend). Electronic Supplementary File (ESF1), Fig. 1 shows our FoxP3+CB1+ gating strategy. We first stained PBMCs with surface markers CD3-PEcy7, CD4-APCcy7, CD25-APC, 7-AAD and CB1-AF700 (human CB1 receptor) (Biolegend). Intracellular Foxp3 staining was performed using the FoxP3/Transcription Factor Staining Buffer Set (eBioscience) for fixation and permeabilisation and following staining with antibody to Foxp3-FITC (Biolegend). Flow cytometry was performed directly after staining, and at least 50,000 lymphocytes were identified based on size and granularity. ESF, Fig. 2 shows our gating strategy to assay CD4/CD8+/CD20+CB2+ cells. The flow cytometry was performed using an LSRII flow cytometer (BD Biosciences), and data were analysed using FlowJo X software (Tree Star Inc., Ashland, OR, USA). As such, we determined the percentage of five different phenotypes, namely CD20+CB2+, CD3+CB2+, CD4+CB2+, CD8+CB2+ and FoxP3+CB1+ cells and this in five conditions: unstimulated or baseline, anti-CD3 and anti-CD28 stimulated, stimulated + 0.1 µg/mL CBD, stimulated + 1.0 µg/mL CBD and stimulated + 10.0 µg/mL CBD.

Figure 1. Partial regression of the depression phenome on stimulated CD20+CB2+ cells.

Figure 2. Partial regression of the depression phenome on baseline CD3+CB2+ cells.
Using the same blood samples, we also quantified cytokines/chemokines in stimulated diluted whole blood culture supernatant, as explained previously (Maes et al., Reference Maes, Rachayon, Jirakran, Sodsai, Klinchanhom, Gałecki, Sughondhabirom and Basta-Kaim2022). We utilised RPMI-1640 medium supplemented with L-glutamine and phenol red and containing 1% penicillin (Gibco Life Technologies, USA), with or without 5 µg/mL PHA (Merck, Germany) + 25 µg/mL LPS (Merck, Germany). On 24-well sterile plates, 1.8 mL of each of these two mediums was mixed to 0.2 mL of 1/10 diluted whole blood. The specimens from each individual were incubated for 72 hours at 37°C, 5% CO2 in a humidified atmosphere. After incubation, the plates were centrifuged at 1500 rpm for 8 minutes. Supernatants were carefully extracted under sterile conditions, divided into Eppendorf tubes, and immediately frozen at −70°C until thawed for cytokine/chemokines/growth factor assays. The cytokines/chemokines/growth factors were measured using the Bio-Plex Pro human cytokine 27-plex assay kit (BioRad, Carlsbad, California, USA) using the LUMINEX 200 equipment (BioRad, Carlsbad, California, United States of America). For all investigations, the intra-assay CV values are less than 11%. ESF, Table 1 shows the immune profiles used in the present study.
Table 1. Clinical and immune features of heathy controls (HC) and major depressed patients (MDD)

Current_SB, current suicidal behaviours; HAMD, Hamilton Depression Rating Scale; IRS, immune-inflammatory response system; Phenome, a score based on the HAMD, STAI and Current_SB scores; M1, macrophage M1; MWUT, Mann–Whitney U test; STAI, State-Trait Anxiety Inventory; Th, T helper.
All results are shown as mean (SD) or as frequencies, F : results of ANOVA, χ2: results of analysis of contingency tables.
T cell growth: a composite score built using cytokines/growth factors that stimulate T cell growth.
Statistics
We compared nominal variables using analysis of contingency tables (χ2-test) and continuous variables using analysis of covariance (ANCOVA) among research groups. For the purpose of examining the connections between scale variables, we employed Pearson’s product moment correlation coefficients. A first prespecified generalised estimating equation (GEE), repeated measures, was used to assess the responsivity of the cell populations to administration of anti-CD3/CD28 (unstimulated vs. stimulated condition), the differences between patients and controls, cell types (CD3, CD4, CD8 and CD20) and all interactions, while allowing for the effects of age, sex, BMI, smoking and drug status of the patients. A second pre-specified GEE, repeated measures, performed on the stimulated cell population only, included fixed categorical effects of diagnosis, treatment (four conditions: anti-CD3/CD28 with and without the three CBD concentrations), cell type and their interactions, while allowing for the effects of the above confounding variables. It should be stressed that there were no missing variables in any of the cell populations or clinical data. Manual multiple regression analysis was used to assess the effects of explanatory variables (cell populations) on dependent variables (clinical profiles). In order to decide which variables would be included and which would be excluded in the final regression model, we also used an automatic forward stepwise regression strategy with a p-to-enter of 0.05, and a p-to-remove of 0.06. We estimated the standardised coefficients with t-statistics and exact p-values, F statistics (and p values), and total variance used as effect size (partial Eta squared). We investigated the likelihood of multicollinearity and collinearity using the tolerance (cut-off value: 0.25) and variance inflation factor (cut-off value: > 4), as well as the condition index and variance proportions from the collinearity diagnostics table. The modified Breusch–Pagan test and the White test were used to investigate possible heteroskedasticity. In the final model, the residuals, residual plots and data quality were always examined. Using the outcomes of the linear modelling analyses, we also computed partial regression analyses, including partial regression plots. The primary statistical analysis is the multiple regression analysis that delineates the prediction of the depression phenome on the immune cell populations. To normalise the data distribution of the indicators, we employed transformations such as logarithmic, square-root, rank-based inversed normal (RINT), and Winsorization when necessary. All tests were two-tailed at p = 0.05. We utilised IBM’s Windows version of SPSS 28 to conduct the aforementioned statistical analyses. A priori power analysis (G*Power 3.1.9.4) for a linear multiple regression shows that given an effect size of 0.25 (equivalent to 20% explained variance), power = 0.8, alpha = 0.05 and three covariates, the minimum sample size should be 48.
Results
Sociodemographic and clinical data
Table 1 shows that there are no differences in age, sex or years of education among the study groups. BMI was somewhat higher in patients, but covarying for BMI in the different analyses did not show any effect of BMI on the immune data. The HAM-D, STAI, Current_SB and phenome scores were significantly higher in patients as compared with controls. Table 1 also shows that the M1, IRS and T cell growth profiles were significantly increased in MDD, while there was a trend towards increased Th-1 and Th-17 scores.
Baseline and stimulated CB2 and CB1 frequencies in controls and patients
GEE analysis performed on the frequency of CB2-bearing cells in the unstimulated and stimulated conditions showed significant effects of stimulation (Wald = 7.37, df = 1, p = 0.007), cell type (Wald or W = 60.75, df = 3, p < 0.001), diagnosis X cell type (W = 15.42, df = 4, p = 0.004) and stimulation X cell type (17.41, df = 3, p < 0.001). All GEE analyses concerning the CB2+/CB1+ data were adjusted for the possible effects of in vivo treatment with psychotropic drugs; however, no significant effects could be found. Table 2 shows the significant interaction pattern between stimulation X cell type. After stimulation, the CB2+ percentage on CD3+ (p < 0.001), CD4+ (P < 0.001) and CD8+ (p = 0.003), but not CD20+ (p = 0.110) cells, had increased. In the baseline and stimulated conditions, the percentage of CB2-bearing cells decreased from CD20+ → CD3+ and CD4+ → CD8+ (p < 0.001). GEE analysis performed on the CD25+FoxP3+CB1+ percentage showed a significant stimulating effect of time (W = 9.33, df = 1, p = 0.002), as shown in Table 2. In baseline conditions, the CD25+FoxP3+CB1+ percentage was lower in MDD than in controls (p = 0.042), whereas after stimulation, no differences could be found (p = 0.902).
Table 2. Differences in CB2- and CB1-bearing cells between the unstimulated (baseline) and anti-CD3/CD28-stimulated condition

Results of generalised estimating equation, repeated measures, indicating a significant interaction between cell type and stimulation by antiCD3/CB28.
Table 3 shows the significant interaction between diagnosis X 4 cell types (CD20+, CD3+, CD4+ and CD8+). GEE analysis performed on the baseline and stimulated conditions showed that there were significant effects of diagnosis (W = 4.58, df = 1, p = 0.032) and interaction diagnosis X stimulation (W = 3.83, df-1, p = 0.050). The percentage of CD20+CB2+ was significantly higher in patients with MDD (8.15 ± 1.05%) compared with controls (4.98 ± 1.06%), whereas there were no significant differences in CD3+CB2+, CD4+CB2+ and CD8+CB2+ cells between MDD and controls. The interaction pattern showed that the percentage of CD20+CB2+ was higher in the stimulated MDD groups than in the three other groups.
Table 3. The significant interaction pattern between diagnosis and cell type in patients with major depressive disorder (MDD) and healthy controls (HC)

Results of generalised estimating equation.
Regression analyses of severity of illness on CB2 percentages
Consequently, we have examined whether the different immune populations and the immune profiles (M1, Th-1, Th-17, IRS and T cell growth) predict the clinical scores. Table 4 shows the results of these multiple regression analyses of clinical severity scores on the unstimulated and stimulated percentages of the immune cells with and without the immune profiles. Regression #1 shows that 29.6% of the variance in the HAMD score was explained by the regression on basal FoxP3+CB2+% and education (both inversely) and stimulated CD20+CB2+% (positively). Regression #2 shows that 34.8% of the variance in the total STAI score was explained by FoxP3+CB2+% and basal CD3+CB2+% (both inversely) and stimulated CD20+CB2+% (positively). Regression #3 shows that 21.2% of the variance in Current_SB was explained by the stimulated values of CD20+CB2+% (positively). Regression #4 shows that 33.2% of the variance in the phenome score was explained by the regression on stimulated values of CD20+CB2+% (positively) and CD3+CB2+%. Figs. 1 and 2 show the partial regression of the phenome on CD20+CB2+% and CD3+CB2+%, respectively.
Table 4. Results of multiple regression analyses with clinical scores or immune profiles as dependent variables and percentage of CB2- and CB1-bearing lymphocytes as explanatory variables

Basal, unstimulated condition; Current_SB, current suicidal behaviours; HAMD, Hamilton Depression Rating Scale; Phenome, a score based on the HAMD, STAI and current_SB scores; STAI, State-Trait Anxiety Inventory; Stim, anti-CD3/CD28 stimulation.
We also examined whether the immune profiles could be predicted by the immune cell populations and found that CD4+CB2+% was the single best significant predictor (inversely) of M1 (explained variance: 8.4%), Th-1 (11.5%), Th-17 (14.4%), IRS (11.0%) and T cell growth (11.9%). Fig. 3 shows the regression of T cell growth on CD4+CB2+%.

Figure 3. Partial regression of T cell growth on CD4+CB2+ cells.
Effects of in vitro administration of CBD
GEE analysis performed on all subjects with stimulated conditions (including the three CBD conditions) showed significant effects of the stimulation/CBD treatment condition (Wald = 18.59, df = 3, p < 0.001), cell type (W = 156.34, df = 3, p < 0.001), diagnosis (W = 4.00, df = 1, p < 0.045), diagnosis X cell type (W = 37.05, df = 3, p < 0.001) and stimulation/CBD treatment X cell type (W = 21.48, df = 3, p < 0.001) on the frequency of CB2-bearing cells. Table 5 shows the mean percentage of CB2-bearing cells in the stimulated and three CBD conditions. The results show that CBDlow and CBDmedium had no significant effect above and beyond anti-CD3/CD28, whereas CBDhigh significantly enhanced all four cell types highly significantly (p < 0.001).
Table 5. Mean percentage of cannabinoid type 2 receptor (CB2)-bearing cells in the anti-CD3/CD28-stimulated condition and three cannabidiol (CBD) treatment conditions
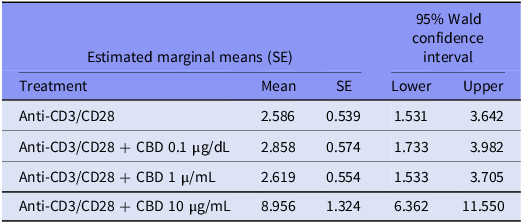
Results of generalised estimating equation.
ESF, Table 2 shows the effects of the three concentrations of CBD on the anti-CD3/CD28 stimulated FoxP3+CB1+ percentage. GEE analysis showed a significant effect of treatment (W = 18.87, df = 3, p < 0.001) but no significant effects of diagnosis, diagnosis X treatment, and use of in vivo psychotropic drugs. Pairwise comparisons show that the FoxP3+CB1+ percentage was significantly increased by 10 µg/mL CBD as compared with the other three stimulated conditions (p < 0.001).
Discussion
CB2/CB1-bearing T and B lymphocytes
The first major finding of this study is that the CB2% was significantly higher in CD20+ cells than CD3+ and CD4+ cells and again higher than in CD8+ cells; stimulation with anti-CD3/CD8 increases the number of CB2-bearing CD3+, CD4+ and CD8+ cells, as well as CB1-bearing FoxP3+ cells; and lower levels of CD4+CB2+ cells are predictive of increased M1, Th-1, Th-17, IRS and T cell growth profiles.
CB2 receptors are predominantly expressed in the immune system and the central nervous system (CNS) (including hippocampal neurons and glial cells), whereas CB1 receptors are predominantly expressed in the CNS, although immune cells also express CB1 receptors (Cabral and Griffin-Thomas, Reference Cabral and Griffin-Thomas2009; Bie et al., Reference Bie, Wu, Foss and Naguib2018). In fact, previous research has demonstrated that B cells express the greatest quantity of CB2 protein and mRNA, followed by natural killer cells, macrophages, and T cells (Galiegue et al., Reference Galiègue, Mary, Marchand, Dussossoy, Carrière, Carayon, Bouaboula, Shire, Le Fur and Casellas1995; Schatz et al., Reference Schatz, Lee, Condie, Pulaski and Kaminski1997; Lee et al., Reference Lee, Newton, Widen, Friedman, Klein, Friedman and Klein2002). Additionally, there is evidence that immune activation is accompanied by increased CB2 expression on tonsillar B cells via CD40 activation (Lee et al., Reference Lee, Newton, Widen, Friedman, Klein, Friedman and Klein2002) and that during inflammation, there are more CB2 molecules available for stimulation (Cabral and Griffin-Thomas, Reference Cabral and Griffin-Thomas2009). Increased CB2 expression in the brain occurs in response to inflammatory conditions and is predominantly localised in activated microglia, but not in quiescent microglia (Nunez et al., Reference Núñez, Benito, Pazos, Barbachano, Fajardo, González, Tolón and Romero2004; Cabral and Marciano-Cabral, Reference Cabral and Marciano-Cabral2005; Fernandez-Ruiz et al., Reference Fernández-Ruiz, Romero, Velasco, Tolón, Ramos and Guzmán2007). Importantly, peripherally activated immune cells that infiltrate the CNS via blood–brain barrier breakdown may contribute to neuroinflammation (Cabral et al., Reference Cabral, Raborn, Griffin, Dennis and Marciano-Cabral2008). Stimulation of immune cells, including with antibodies, may increase CB1 mRNA in murine whole spleen cells, whereas stimulation with anti-CD3 antibodies may decrease CB1 mRNA (Noe et al., Reference Noe, Newton, Widen, Friedman, Klein, Friedman and Klein2002).
In our study, decreased levels of CD4+CB2+ cells predicted increased immune-inflammatory profiles. This is consistent with previous research demonstrating that increased CB2 expression or B2 ligation maintains immune homoeostasis and has anti-proliferative and anti-inflammatory effects by downregulating T cell functions and T cell effector activities, Th-17 activation, CD4+ cell proliferation, IFN-γ and IL-2 production, inducing Th-2 polarisation, and attenuating macrophage functions and chemotaxis (Ziring et al., Reference Ziring, Wei, Velazquez, Schrage, Buckley and Braun2006; Cabral and Griffin-Thomas, Reference Cabral and Griffin-Thomas2009; Guillot et al., Reference Guillot, Hamdaoui, Bizy, Zoltani, Souktani, Zafrani, Mallat, Lotersztajn and Lafdil2014; Eisenstein and Meissler, Reference Eisenstein and Meissler2015; Gentili et al., Reference Gentili, Ronchetti, Ricci, Di Paola, Gugliandolo, Cuzzocrea, Bereshchenko, Migliorati and Riccardi2019). It should be noted that activated CD20+CB2+ cells may increase IL-10, a negative immunoregulatory cytokine, and are implicated in Th-2 polarisation (Agudelo et al., Reference Agudelo, Newton, Widen, Sherwood, Nong, Friedman and Klein2008; Carter et al., Reference Carter, Rosser and Mauri2012; Saroz et al., Reference Saroz, Kho, Glass, Graham and Grimsey2019). On the basis of these findings, CB2 ligation with selective agonists is deemed an effective treatment for conditions marked by immune activation and autoimmunity (Eisenstein and Meissler, Reference Eisenstein and Meissler2015). CB2 on autoreactive T cells, for instance, may suppress autoimmune inflammation in the CNS (Maresz et al., Reference Maresz, Pryce, Ponomarev, Marsicano, Croxford, Shriver, Ledent, Cheng, Carrier, Mann, Giovannoni, Pertwee, Yamamura, Buckley, Hillard, Lutz, Baker and Dittel2007). Immune CB1 may also possess anti-inflammatory and immunosuppressive properties, inhibit the production of pro-inflammatory cytokines such as IL-2 and IL-12, and induce apoptosis in immune cells (Kaplan, Reference Kaplan2013).
Lowered immune homoeostasis in MDD
The second main finding of this study is that MDD is associated with decreased baseline levels of FoxP3+CB1+ cells, and that decreased FoxP3+CB1+% and CD3+CB2+% strongly predicted the phenome of depression, including the severity of depression, anxiety and current suicidal behaviours. Reduced FoxP3+ and CD3+CB2+ are likely to reduce the immune homoeostatic processes of CB2/CB1 on T cells and FoxP3+ cells, thereby reducing the immunosuppressive and anti-inflammatory capacities of CB2 and CB1. Consequently, deficiencies in CB receptor markers may account for at least some of the immune findings in depression, including increased T cell activation, CD4+ cell proliferation, increased IL-2 and IFN-γ production, Th-1 polarisation, Th-17 activation, and decreased Treg differentiation (Maes et al., Reference Maes, Bosmans, Suy, Vandervorst, De Jonckheere and Raus1990; Maes et al., Reference Maes, Rachayon, Jirakran, Sodsai, Klinchanhom, Gałecki, Sughondhabirom and Basta-Kaim2022). Moreover, the cell population markers evaluated in this study are better predictors of the phenome of depression than the immune profiles, which we computed using pro-inflammatory cytokines, chemokines and T cell growth factors. Interestingly, LPS, which is elevated in MDD due to leaky gut and increased bacterial translocation (Maes et al., Reference Maes, Kubera, Leunis, Berk, Geffard and Bosmans2013), may downregulate CB2 in splenocytes (Lee et al., Reference Lee, Newton, Widen, Friedman, Klein, Friedman and Klein2002). Therefore, it is safe to hypothesise that the increased bacterial translocation in MDD may have impacted CB2%. The consequent decreased immune homoeostasis as a result of deficits of not only CD3+CB2+% and CD4+CB2+%, but also FoxP3+CB1+% may, therefore, contribute to the peripheral inflammatory and neuroinflammatory responses, which contribute to the pathogenesis of severe MDD.
Increased CD20+CB2+ cells in MDD
The third main finding of this study is that MDD is associated with a strong increase in CD20+CB2+% following anti-CD3/CD28 activation, and that this is markedly and positively associated with the depression phenome and recent suicidal behaviours. In significant subgroups of persons with MDD, previous research has reported both T cell activation and B cell proliferation (Maes, Reference Maes1995). Importantly, we found that patients with MDD have a substantially increased percentage and expression of CD154+ (CDL40+) on CD3+ and CD4+ cells, indicating T cell activation via the T cell receptor (TCR) complex, which is a component of CD3 and CD4/8 (Rachayon et al., submitted). T cell activation via anti-CD3/CD28 involves three steps: a) TCR engagement (with beads or antibodies by antigen-presenting cells); b) co-stimulatory signals through CD28, which further activate CD4+ and CD8+ T cells; and c) T-dependent activation of B cells via CD40-CD40L interactions (Thermo-Fisher, 2023; Klaus et al., Reference Klaus, Pinchuk, Ochs, Law, Fanslow, Armitage and Clark1994; Grewal and Flavell, Reference Grewal and Flavell1996). In fact, the upregulation of CD40L is a strict requirement for T-dependent B cell activation (Klaus et al., Reference Klaus, Pinchuk, Ochs, Law, Fanslow, Armitage and Clark1994). Moreover, CD40 ligation results in the upregulation of CB2 on B cells, with CB2+ B cells playing an essential role in B cell differentiation (Carayon et al., Reference Carayon, Marchand, Dussossoy, Derocq, Jbilo, Bord, Bouaboula, Galiègue, Mondière, Pénarier, Fur, Defrance and Casellas1998). These findings indicate that the increased proportion of CB2+ B cells in MDD is likely the result of T cell activation and, more specifically, CD40-CD40L interactions.
However, CB2 is also a crucial component of T-independent humoral immunity and B cell-associated immunity (Basu et al., Reference Basu, Ray and Dittel2011). Importantly, CB2 upregulates MZ B cells, which, as part of the innate immune response, produce natural IgM (see Introduction; Rayman et al., Reference Rayman, Lam, Laman, Simons, Löwenberg, Sonneveld and Delwel2004; Basu et al., Reference Basu, Ray and Dittel2011). Recent reviews demonstrate that the polyreactive natural IgM produced by B1 cells is directed against a number of self-antigens and commensal bacterial antigens, exerts protective, anti-inflammatory, and antioxidative effects, clears cell debris, and serves as a first-line defence against pathogen invasion (Morris et al., Reference Morris, Puri, Olive, Carvalho, Berk and Maes2019; Allman et al., Reference Allman, Wilmore and Gaudette2019). CB2 deficiency induces deficits in natural IgM and humoral immune responses (Basu et al., Reference Basu, Ray and Dittel2011). In addition, T cell-independent antigens, such as certain commensal microbes and viruses, may prime MZ B cells to respond quickly to activation stimuli (Basu et al., Reference Basu, Ray and Dittel2011). As discussed in the Introduction, MDD is associated with elevated levels of IgM antibodies to numerous self-antigens, oxidatively and nitrosatively modified neoepitopes, and the LPS of commensal Gram-negative bacteria. Consequently, T cell activation and the resulting increase in CD20+CB2+% may play a crucial role in the increase in natural IgM levels.
Nevertheless, these B cells can also have detrimental effects, such as pro-inflammatory effects, activating innate response activator cells, acting as antigen-presenting cells that stimulate T cells to produce more pro-inflammatory cytokines, inducing Th-1 and Th-17 responses, and inhibiting Treg cell formation (Morris et al., Reference Morris, Puri, Olive, Carvalho, Berk and Maes2019). In addition, some B1 cells produce high-affinity IgM which activates the complement system and induces inflammation (Kerfoot et al., Reference Kerfoot, Szczepanik, Tung and Askenase2008; Askenase et al., Reference Askenase, Bryniarski, Paliwal, Redegeld, Groot Kormelink, Kerfoot, Hutchinson, van Loveren, Campos, Itakura, Majewska-Szczepanik, Yamamoto, Nazimek, Szczepanik and Ptak2015). Abnormal B1 cell trafficking may contribute to autoimmunity, as seen in an autoimmune animal model of systemic lupus erythematosus (Morris et al., Reference Morris, Puri, Olive, Carvalho, Berk and Maes2019). B1 cells also produce granulocyte macrophage colony-stimulating factor (GM-CSF), one of the growth factors that is increased in MDD (Maes et al., Reference Maes, Rachayon, Jirakran, Sodsai, Klinchanhom, Gałecki, Sughondhabirom and Basta-Kaim2022) and which may further activate the production of IgM (Weber et al., Reference Weber, Chousterman, Hilgendorf, Robbins, Theurl, Gerhardt, Iwamoto, Quach, Ali, Chen, Rothstein, Nahrendorf, Weissleder and Swirski2014; Zasada et al., Reference Zasada, Rutkowska-Zapała, Lenart and Kwinta2016). Finally, when triggered by bacterial antigens by ligation of their toll-like receptors or the B cell receptor (BCR), B cells egress to the spleen and develop into plasma cells secreting cytokines and pathogen-specific IgM (Savage and Baumgarth, Reference Savage and Baumgarth2015; Baumgarth et al., Reference Baumgarth, Waffarn and Nguyen2015; Rahman et al., Reference Rahman, Sing, Golabkesh, Fiskesund, Gustafsson, Jogestrand, Frostegård, Hafström, Liu and Frostegård2016). It should be added that in some conditions, CB2 ligation with agonists may enhance inflammatory responses (Karsak et al., Reference Karsak, Gaffal, Date, Wang-Eckhardt, Rehnelt, Petrosino, Starowicz, Steuder, Schlicker, Cravatt, Mechoulam, Buettner, Werner, Di Marzo, Tüting and Zimmer2007). Furthermore, 2-AG may stimulate the secretion of chemokines that enhance leucocyte adhesion and recruitment, and the production of TNF-α by endothelial cells (Gasperi et al., Reference Gasperi, Evangelista, Chiurchiù, Florenzano, Savini, Oddi, Avigliano, Catani and Maccarrone2014; Atalay et al., Reference Atalay, Jarocka-Karpowicz and Skrzydlewska2019; Jehle et al., Reference Jehle, Eich, Danisch, Bagheri, Avraamidou, Pfeifer, Tiyerili, Bindila, Lutz and Nickenig2021). CB2 may act as a costimulator in mitogen-activated kinase (MAPK) and stress-activated kinase (SAPK) pathways, which promote mitosis, cell survival, and differentiation (Carayon et al., Reference Carayon, Marchand, Dussossoy, Derocq, Jbilo, Bord, Bouaboula, Galiègue, Mondière, Pénarier, Fur, Defrance and Casellas1998).
Overall, the T cell activation in MDD may have stimulated B cells to acquire more CB2, which, in conjunction with the increased bacterial translocation in MDD, may result in an overproduction of IgM antibodies, which may have a variety of deleterious effects that exacerbate the primary T cell activation and thus the severity of the illness’s phenome. Moreover, increased CB2 may contribute to increased bacterial load in the systemic circulation due to increased production of IL-10 (Csoka et al., Reference Csóka, Németh, Mukhopadhyay, Spolarics, Rajesh, Federici, Deitch, Bátkai, Pacher and Haskó2009).
Effects of CBD
The fourth major finding of this study is that CBD concentrations between 0.1 and 1 µg/dL (therapeutic levels) had no discernible effect on any of the stimulated lymphocyte populations evaluated in this study. Previously, we did not observe any effect of CBD on the elevated levels of pro-inflammatory markers (M1, Th-1, Th-17, IRS and T cell proliferation) and cytokines in MDD patients or healthy controls (Rachayon et al., Reference Rachayon, Jirakran, Sodsai, Klinchanhom, Sughondhabirom, Plaimas, Suratanee and Maes2022). These results demonstrate that CBD does not normalise the abnormalities in the prevalence of CD20+CB2+ populations and does not ameliorate the deficiencies in CD3+/CD4+CB2+ populations that predict the phenome of depression. CBD has a lower affinity for CB1 and CB2 compared to other cannabinoids such as AEA, 2-AG and THC, as described in the Introduction. CBD is a partial agonist at CB2, which may at least partially mediate CBD’s anti-inflammatory effects (An et al., Reference An, Peigneur, Hendrickx and Tytgat2020). In addition, CBD desensitises the transient receptor potential cation channel, subfamily V member 1, which has inflammatory activities, and has agonist activities at the transient receptor potential cation channel, subfamily A, member 1 (the Wasabi Receptor), which promotes inflammation, depression, and anxiety, and acts as an antagonist at the transient receptor potential cation channel, subfamily M, member 8, which attenuates inflammation (Muller et al., Reference Muller, Morales and Reggio2019; An et al., Reference An, Peigneur, Hendrickx and Tytgat2020).
In addition, the higher CBD levels of 10 µg/dL substantially increase the stimulated levels of CD20+/CD3+/CD + 4/CD8+ CB2+ cells following anti-CD3/CD28 treatment. Previously, we reported that 10 µg/dL CBD had significant effects on the immune system, inhibiting CIRS functions such as the stimulated production of IL-10, IL-13, and sIL-1RA, and IRS profiles, such as Th-1 and Th-17 (Rachayon et al., Reference Rachayon, Jirakran, Sodsai, Klinchanhom, Sughondhabirom, Plaimas, Suratanee and Maes2022). Consequently, higher CBD doses may have detrimental effects.
Limitations
This study would have been more interesting if we had measured CB2 in other immune populations, including macrophages, dendritic cells and natural killer cells. Future research should investigate the effects of CBD and other cannabinoids on the stimulated number and expression of CB2+ immunocytes. It could be argued that the sample size is rather small, although the minimal sample size was determined using power analysis. Furthermore, post hoc power analysis performed on the primary analysis (regression of phenome on immune cell populations) shows that the actual power obtained is 0.98.
Conclusions
There was an inverse relationship between the number of reduced CD4+CB2+ cells and M1, Th-1 and Th-17 phenotypes. MDD is characterised by a reduced FoxP3+CB1+ percentage and a higher CD20+CB2+ percentage. CD20+CB2+% (positively) and CD3+CB2+% (negatively) explained 33.2% of the variance in the depression phenome which comprises severity of depression, anxiety and current suicidal behaviours. Decreases in FoxP3+CB1+% and CD3+/CD4+CB2+% contribute to immune homoeostasis deficits in MDD, whereas an increase in CD20+CB2+% may contribute to the pathogenesis of MDD by activating T-independent humoral immunity. CB1 on Treg cells and CB2 on CD3/CD4+ and CD20+ cells are new drug targets to treat MDD.
Supplementary material
The supplementary material for this article can be found at https://doi.org/10.1017/neu.2023.35.
Data availability statement
The dataset generated during and/or analysed during the current study will be available from the corresponding author (M.M.) upon reasonable request and once the authors have fully exploited the dataset.
Acknowledgements
AMERI-ASIA MED CO, Ltd, supported this work.
Author contribution
Design of the study: MM and MR. Recruitment of the participants: MR and KJ. Assays: PS. Statistical analyses: MM. Visualisation: MM and PS. First draft: MM. Editing: MR, KJ, AS, AFA and PS. All authors agreed to publish the final version of the manuscript.
Competing interests
None.
Ethical statement
All subjects gave their informed consent for inclusion before they participated in the study. The study was conducted in accordance with the Declaration of Helsinki of 1975, revised in 2013, and the protocol was approved by the Ethics Committee of the Institutional Review Board of the Faculty of Medicine, Chulalongkorn University, Bangkok, Thailand (#528/63).











 Open access
Open access