


Gustavson and his colleagues (Reference Gustavson, Miyake, Hewitt and Friedman2014) report a genetic correlation of 1.0 between procrastination and impulsivity based on model-fitting to data from 181 monozygotic (MZ) and 166 same-sex dizygotic (DZ) twin pairs from Colorado. As it happens, data are available from two considerably larger Australian twin samples that allow for a partial check on this value. One sample is of adult twins covering a range of ages (mean age 42.5 years, SD = 13.0), and the other was of young adult twins (mean age 23.2, SD = 2.19). The latter were similar in age to the Colorado sample (mean age 22.7 years, SD = 1.12). There were 1,310 MZ and 748 same-sex DZ pairs with appropriate data in the general adult Australian sample, and 702 MZ and 472 same-sex DZ pairs in the young adult sample. Only same-sex DZ pairs from the Australian samples were used for consistency with Gustavson et al.'s (Reference Gustavson, Miyake, Hewitt and Friedman2014) study.
Gustavson et al. (Reference Gustavson, Miyake, Hewitt and Friedman2014) argue that their evidence that procrastination and impulsivity have a genetic correlation of 1.0 tends to support the argument of Steel (Reference Steel2010) that procrastination is an evolutionary byproduct of impulsivity. However, they acknowledge that contemporary correlational evidence cannot be conclusive on this point. We would add that if the genetic correlation between procrastination and impulsivity is considerably less than 1.0, a variety of hypotheses concerning the relationships (evolutionary or otherwise) between the two traits become tenable.
Methods
Participants
The participants in the Australian samples were volunteer adult twins who completed mail questionnaires containing, among other topics, two sets of items describing their personalities. The general adult sample was a 1988–1989 follow-up of a 1980–1981 twin study. The younger sample comprised twins identified in the earlier study who were too young to participate at that time but were surveyed as young adults in 1988–1990. The samples for the present analyses comprised 394 male and 916 female MZ and 223 male and 525 female DZ pairs from the general adult sample, and 250 male and 452 female MZ and 162 male and 310 female DZ pairs from the young adult sample. The greater proportions of MZ and female pairs are characteristics of volunteer twin samples (Lykken et al., Reference Lykken, Tellegen and DeRubeis1978). The numbers given are after the exclusion of twins for missing data (see below).
Measures
The questionnaires completed by the twins in the general adult sample contained two sets of personality questionnaire items; the first set was of items largely from the Eysenck Personality Questionnaire (Eysenck et al., Reference Eysenck, Eysenck and Barrett1985), and the second set was of items largely from Cloninger's Tridimensional Personality Questionnaire (Cloninger et al., Reference Cloninger, Przybeck and Svrakic1991). The same personality items were included for both samples (in somewhat different order) with additional items added in the case of the young adult sample.
Among the Eysenck items was a direct question on procrastination: ‘Do you sometimes put off until tomorrow what you ought to do today?’ The questionnaires had several items related to the other trait, impulsivity. The ones used for this study (selected a priori) are listed in Table 1. (The Eysenck items had a ‘Yes’ or ‘No’ response format, and the Cloninger items had a ‘True’ or ‘False’ format.) Items reversed in scoring are marked (R).
TABLE 1 Items Used in the Present Study to Measure Procrastination and Impulsivity

The initial scoring of items was 1 for ‘No’ or ‘False’, and 3 for ‘Yes’ or ‘True’. If an individual failed to respond to more than 10% of the personality questionnaire items, he or she was excluded from further analyses; otherwise the occasional omitted items were scored 2 for ‘Don't know’. If one twin was excluded, the co-twin was dropped as well to retain intact pairs. From the initial sample, 22.9% of individuals were dropped as members of opposite-sex pairs; 11.6% of those remaining were excluded because one or both twins had excessive missing data.
Analyses
The scores for each individual on the impulsivity items were summed after reflecting as necessary. Polychoric correlations between the procrastination item and the impulsivity score were calculated across twin pairs by PRELIS (version 2.80) in each of the eight subgroups defined by sample, sex, and zygosity. The correlations were then fitted by LISREL (version 8.80) to the model illustrated in Figure 1. In line with Gustavson et al. (Reference Gustavson, Miyake, Hewitt and Friedman2014), the model fitted comprised additive genes (A) and non-shared environment (E) — the latter including errors of measurement. Models were fit separately for the two sexes and for the two samples and for the four sex and sample combinations as well as for the combined MZs and DZs. Degrees of freedom (df) were adjusted for the use of correlation matrices, as described by Neale and Cardon (Reference Neale and Cardon1992, p. 256).
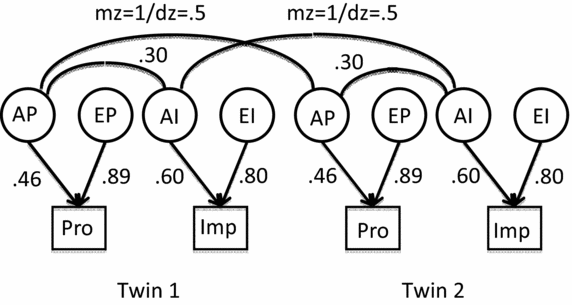
FIGURE 1 Path model fit to Australian twin data. Note: Observed variables (squares) Pro = procrastination, Imp = Impulsivity. Latent variables (circles) AP = additive genetic effects for procrastination, EP = unshared environmental effects for procrastination, and so on. Curved lines at top of figure = correlations among latent variables not shown are cross-twin correlations between AP and AI, equal to the within-twin correlation for MZs, and one half that for DZs. All variables are standardized.
Results
The obtained polychoric correlations (based on one categorical and one continuous variable) are given in Table 2.
TABLE 2 Correlations for Procrastination and Impulsivity for MZ and DZ Twin Pairs for Men and Women in Two Australian Samples (MZ Twins Below Diagonal, DZ Above Diagonal)

P = Procrastination, I = Impulsivity; subscripts 1 and 2 refer to the 1st and 2nd twin of a pair.
Combined N: 2,012 MZ, 1,220 same-sex DZ pairs.
The model of Figure 1 was fit to correlation matrices for the eight groups, with five parameters constrained to equality: the paths from the additive genes, A, to the two traits, the paths from the unshared environments, E, to the two traits, and the genetic correlation r g between procrastination and impulsivity. With these large samples, one can confidently rule out an exact fit to the data (χ2 = 201.95 for df = 45, p < .001). Allowing for different r g values for men and women did not lead to a significant improvement in fit, χ2 diff = 1.22 for df = 1, p = .27; nor did allowing r g to differ between the general adult and the young adult samples, χ2 diff = 2.16 for df =1, p = .14; nor did allowing r g to differ for both, χ2 diff = 5.55 for df = 3, p = .14. Therefore, the four MZ and the four DZ subgroups were pooled, and the Figure 1 model fit to the correlations in the combined groups. An exact fit could still be rejected (χ2 = 34.90 for df = 10, p < .001). However, the overall fit was good as judged by the Root Mean Square Error of Approximation (RMSEA) and the various goodness of fit indices reported by LISREL. RMSEA values of <0.05 are considered to represent close model fits (Browne & Cudeck, Reference Browne, Cudeck, Bollen and Long1993). The RMSEA value for the combined Australian data was 0.029, with a 90% Confidence Interval of 0.017–0.042, making it reasonable to consider the same parameters as holding across MZ and DZ groups. The six goodness-of-fit indices lay in the range of 0.926 to 1.158, with a median value of 0.956; values above 0.90 are usually considered to represent satisfactory model fits. Obtained values of the parameters for this model are shown on the paths in Figure 1.
The crucial question for comparison with the Gustavson et al.'s (Reference Gustavson, Miyake, Hewitt and Friedman2014) finding is whether the obtained r g of 0.299 is significantly different from 1.0. The LISREL solution gives the standard error of r g as 0.043, suggesting that values in the neighborhood of 0.2 to 0.4 for this parameter might be plausible, but that 1.0 is extremely unlikely. Fitting the model with r g fixed to 1.0 produced a χ2 of 120.53 for df = 11, p < .001. Clearly, the Australian twin data failed to support the value of 1.0 obtained by Gustavson et al. (Reference Gustavson, Miyake, Hewitt and Friedman2014) for the genetic correlation between procrastination and impulsivity, although these are consistent with a genetic correlation of around 0.2 or 0.3.
Discussion
Although the estimate of the genetic correlation between procrastination and impulsivity from the Australian twin data is markedly lower than that reported by Gustavson et al. (Reference Gustavson, Miyake, Hewitt and Friedman2014), some differences between the two analyses must be considered. The Australian study had much larger samples, but the Colorado study had more reliable measurement. In the Australian data, procrastination was measured by a single item, in the Colorado data, this was measured by three scales. For impulsivity, the measurements were based on 9 and 42 items respectively. The reliabilities of the individual scales in the Colorado study ranged from 0.77 to 0.92 (Gustavson et al., Reference Gustavson, Miyake, Hewitt and Friedman2014, Table 3). The internal consistency reliability (Cronbach α) for the nine-item impulsivity scale in the Australian study was 0.70 in the younger and 0.72 in the older sample. The test-retest reliability for the single procrastination item was available for a subsample of 881 individuals from the general adult sample who were tested twice. It was 0.67 (polychoric correlation). The test-retest correlation for the impulsivity score for the same individuals was 0.71, in good agreement with the internal consistency α of 0.72 for the entire sample. Lower reliability would affect the estimated heritability of a measure — decreasing the value of the path from A and increasing the path from E by the virtue of the added error variance. However, the genetic correlations, which are of principal interest, are between latent (i.e., error-free) variables in both studies, and should be independent of the reliabilities of the measures. That is, with lower reliability one would expect a lower phenotypic correlation and lower paths from A, but not necessarily a lower correlation between the As for the two traits. If the phenotypic r and the heritabilities are reduced proportionately by unreliability, then the r g values should remain the same. Thus, a lesser reliability of measures cannot in itself account for the difference in r g values between the Colorado and Australia data.
Another difference between the two analyses was that in the Figure 1 model the correlation between the unshared environments for the two traits (call it r e) is assumed to be zero whereas in the Colorado data it was estimated as 0.33. In the Australian model-fitting, it was possible to estimate r g and r e simultaneously — the values came out to be 0.21 and 0.06 — that is, an even greater discrepancy for r g. However, the model shown in Figure 1, with r e fixed at zero, was used to maximize the estimate of r g; thus it would bring it closer to the Colorado 1.0 than specifying any other (positive) value for r e.
If the genetic correlation between procrastination and impulsivity is really more like 0.3 (or 0.2) than like 1.0, what are the implications for the hypothesis of an evolutionary connection between the two traits? Not very strong either way, in our view. Procrastination may have been (in part) an evolutionary derivative of impulsivity, but the level of impulsivity itself must have been stabilized in human evolution by other traits (100% impulsive behavior would hardly have been conducive to survival), and there are presumably other biologically based traits such as short- and long-term memory abilities that would affect the degree to which impulsivity is linked to procrastination.
In short, anyone who wishes to believe that procrastination is an evolutionary byproduct of impulsivity may continue to do so even if the genetic correlation between the two traits is 0.30 rather than 1.0. In fact, he or she might even have a somewhat easier theoretical task, in that it is hard to believe in a complete genetic overlap between the two. Even if procrastination were indeed an evolutionary byproduct of impulsivity, it seems likely that not all aspects of impulsivity led to procrastination and that some aspects of procrastination developed independently of impulsivity. However, a genetic correlation of 0.30 between the two traits would be susceptible to many differing interpretations, such as current causation in either direction, the effect of a genetically influenced third variable on the two traits, mutual interaction, shared neural sub-processes, and so on.
Acknowledgments
We are grateful to Andrew Heath for insuring that the impulsivity items were included in the initial study, and to Anthony Conciatore for his assistance in obtaining the retest data. Data collection was supported by Alcohol, Drug Abuse and Mental Health Administration (ADAMHA), Grant AA07728, and by a grant from the Australian National Health and Medical Research Council.



