



Los lobos marinos (otáridos) fueron recursos vitales para muchas poblaciones humanas costeras que habitaron Sudamérica. Estos mamíferos marinos constituyeron importantes fuentes de energía y nutrientes (Ambrústolo y Beretta Reference Ambrústolo and Beretta2018; Belardi et al. Reference Belardi, Carballo and L'Heureux2011; Borella Reference Borella2006; deFrance et al. Reference deFrance, Keefer, Richardson and Álvarez2001; Frontini y Bayón Reference Frontini, Bayón, Mondini, Muñoz and Fernández2017; Inda et al. Reference Inda, del Puerto, Castiñeira, Capdepont, García-Rodríguez, Menafra, Rodríguez-Gallego, Scarabino and Conde2006; Lavallée et al. Reference Lavallée, Julien, Béarez, Bolaños, Carré, Chevalier and Delabarde2011; López et al. Reference López, Piñeiro, Castiñeira and Gascue1997; Moreno Reference Moreno2008; Muñoz Reference Muñoz and Goñalons2004; Muñoz et al. Reference Muñoz, Cruz, Lemaire, Pretto, Zangrando, Barberena, Gil, Neme, Giardina, Luna, Otaola, Paulides, Salgán and Tivoli2013; Olguín et al. Reference Olguín, Castro, Castro, Peña-Villalobos, Ruz and Santander2015; Orquera y Piana Reference Orquera and Piana1987, Reference Orquera and Piana1999a; Reitz Reference Reitz1988; Salazar et al. Reference Salazar, Figueroa, Andrade, Salinas, Olguín, Power, Rebolledo, Parra, Orellana and Urrea2015; San Román Reference San Román2016; Schiavini Reference Schiavini1990; Volkmer de Castilho y Simões-Lopes Reference Volkmer de Castilho and Simões-Lopes2008), además de proporcionar una amplia diversidad de materias primas para la producción de tecnología de navegación (Orquera y Piana Reference Orquera and Piana1999a, Reference Orquera and Piana1999b), instrumentos de caza y procesamiento (Beretta et al. Reference Beretta, Zubimendi, Ciampagna, Ambrústolo and Castro2013; Christensen Reference Christensen2016; Orquera y Piana Reference Orquera and Piana1999a; Salazar et al. Reference Salazar, Figueroa, Andrade, Salinas, Olguín, Power, Rebolledo, Parra, Orellana and Urrea2015; San Román Reference San Román2016), contenedores (Emperaire Reference Emperaire1963; Orquera y Piana Reference Orquera and Piana1999b), ornamentos (Capdepont y Pintos Reference Capdepont, Pintos, Mazzanti, Berón and Oliva2002; Fiore Reference Fiore2012; Moreno Reference Moreno2008; Orquera y Piana Reference Orquera and Piana1999a, Reference Orquera and Piana1999b), e incluso artefactos que se trasladaron largas distancias como bienes de intercambio (Sayre et al. Reference Sayre, Miller and Rosenfeld2016).
En términos generales, los otáridos han sido capturados mediante un amplio rango de estrategias de aprovisionamiento, dependientes posiblemente de la dinámica espacial y estacional de estos animales (Lyman Reference Lyman1989, Reference Lyman1995), así como de la tecnología disponible (Orquera y Piana Reference Orquera and Piana1999a). Es así que la distribución actual y el comportamiento se han utilizado como marcos de referencia, en diferentes regiones de Patagonia, para dar cuenta de la variabilidad en sus modos de explotación (Borella Reference Borella2006; Cruz et al. Reference Cruz, Muñoz, Ercolano, Lemaire, Pretto, Nauto and Moreno2015; Lyman Reference Lyman1989, Reference Lyman1995; Orquera y Piana Reference Orquera and Piana1999a; Schiavini Reference Schiavini1990). No obstante, diversos factores vinculados a la formación de conjuntos arqueológicos requieren mayor discusión, especialmente porque algunos datos no son compatibles con los parámetros ecológicos actuales (Borella Reference Borella2006; Martinoli et al. Reference Martinoli, Crespo, Dellabianca and Zangrando2021a), o al menos conducen a cuestionamientos sobre el uso de la distribución actual de los otáridos para efectuar inferencias. Esta tensión en parte responde a que procesos tafonómicos, cambios en la distribución de los recursos como consecuencia de la explotación humana, y/o cambios ambientales y en la selección de las presas, son factores que intervienen en la formación del registro arqueológico y no siempre son considerados de manera integrada en los análisis. En efecto, los avances metodológicos generados a partir de estudios osteométricos (Borella y L'Heureux Reference Borella and L'Heureux2014; Borella et al. Reference Borella, L'Heureux and Grandi2014; L'Heureux y Borella Reference L'Heureux and Borella2011), tafonómicos (Borella y Borrero Reference Borella, Borrero, Gutiérrez, Nigris, Fernández, Giardina, Gil, Izeta, Neme and Yacobaccio2010, Reference Borella and Borrero2019; Borella y Muñoz Reference Borella and Muñoz2006; Martinoli et al. Reference Martinoli, Gutiérrez and Zangrando2021b; Muñoz Reference Muñoz2008), anatómicos (Martinoli Reference Martinoli2019a; San Román Reference San Román, López, Cartajena, García and Mena2009) y de isótopos estables (Vales et al. Reference Vales, Cardona, Zangrando, Borella, Saporiti, Goodall, de Oliveira and Crespo2017; Zangrando et al. Reference Zangrando, Panarello and Piana2014), han permitido mejorar la comprensión de la paleoecología de otáridos sudamericanos y la interacción de estos mamíferos con los grupos humanos durante el Holoceno.
En el sur de Tierra del Fuego, si bien resulta evidente la importancia económica que han tenido los otáridos para las poblaciones humanas (Horwitz Reference Horwitz1990; Orquera y Piana Reference Orquera and Piana1999a; Vidal Reference Vidal1985; Zangrando et al. Reference Zangrando, Tessone, Vázquez, Salemme, Santiago, Álvarez, Piana, Vázquez and Mansur2009), es posible observar cierta variabilidad en las estrategias de aprovisionamiento entre los distintos sectores que conforman ese amplio espacio costero (Martinoli Reference Martinoli2017, Reference Martinoli2018; Schiavini Reference Schiavini1990). Geográficamente, el sur de Tierra del Fuego incluye sectores de canales interiores (canal Beagle), espacios costeros exteriores (Península Mitre) e islas offshore (Isla de los Estados). Las características de los ecosistemas marinos circundantes a esos espacios presentan diferentes condiciones para el emplazamiento de grandes loberías reproductivas de otáridos (Túnez et al. Reference Túnez and Humberto Cappozzo2008), en tanto la disponibilidad de alimentos en sectores de frentes marinos como Península Mitre (zona de mareas) e Isla de los Estados (frente a la plataforma continental) es mayor que la de sectores costeros interiores (Acha et al. Reference Acha, Mianzan, Guerrero, Favero and Bava2004:86, Figura 1D). Si bien hemos identificado cambios en el comportamiento trófico de los otáridos a lo largo del Holoceno (Nye et al. Reference Nye, Zangrando, Martinoli and Fogel2020), por el momento no hay datos que indiquen cambios en su distribución a gran escala (dependiente de la productividad marina) ni en su comportamiento estacional en el archipiélago fueguino.
Nuestro objetivo es efectuar una revisión sobre la explotación humana de otáridos en la costa sur de Tierra del Fuego e Isla de los Estados. Planteamos una evaluación principalmente espacial sobre el tipo de presa explotada por sociedades cazadoras-recolectoras durante el Holoceno. Partimos de la hipótesis de que la explotación de otáridos en Península Mitre e Isla de los Estados implicó estrategias de aprovechamiento distintas a las del sector de canales interiores (canal Beagle), propuesta que luego se discute al integrar otras líneas de evidencia. A continuación presentamos la información antecedente que sustenta esta hipótesis y las expectativas para el registro zooarqueológico.
Distribución, comportamiento y dimensiones de otáridos en el extremo sur de Sudamérica: Explotación humana y expectativas arqueológicas
Las dos especies de lobos marinos (familia otariidae) más abundantes en Sudamérica y en el archipiélago fueguino son Arctocephalus australis y Otaria flavescens (Figura 1). Ambas ocupan espacios marinos y terrestres dependiendo de la época del año, de la edad y del sexo. Son poligínicas y su ciclo anual se divide en una etapa reproductiva en verano y un período de visitas a espacios costeros, en los que forman apostaderos de descanso. Los machos adultos y los individuos inmaduros se agrupan en asentamientos específicos, y permanecen más tiempo en el agua buscando alimentos. Las hembras regresan continuamente a las zonas de cría para amamantar a los cachorros por un período aproximado de un año (Bowen et al. Reference Bowen, Beck, Austin, Perrin, Würsig and Thewissen2009; Crespo et al. Reference Crespo, García, Dans and Pedraza2008). Por lo dicho, la presencia de individuos menores de un año y de hembras adultas se puede considerar un indicador de la explotación de espacios reproductivos (Borella Reference Borella2006; Etnier Reference Etnier2007; Lyman Reference Lyman1989), mientras que el predominio de subadultos y machos adultos se interpreta como consecuencia de la captura de individuos aislados (Orquera y Piana Reference Orquera and Piana1999a; Schiavini Reference Schiavini1990).

Figura 1. Mapa con indicación de los sitios arqueológicos estudiados y la distribución actual de Arctocephalus australis y de Otaria flavescens en Sudamérica, y de sus colonias en la costa sur de Tierra del Fuego e Isla de los Estados (Crespo et al. Reference Crespo, Schiavini, García, Franco-Trecu, Goodall, Rodríguez, Morgante and Oliveira2015; Falabella et al. Reference Falabella, Campagna and Croxall2009; Milano et al. Reference Milano, Grandi, Schiavini and Crespo2020a, Reference Milano, Grandi, Schiavini and Crespo2020b; Schiavini et al. Reference Schiavini, Crespo and Szapkievich2004; Sielfeld et al. Reference Sielfeld, Venegas, Atalah and Torres1978).
Estas dos especies presentan variaciones en sus tamaños. A. australis alcanza las siguientes dimensiones: crías entre 3,5 m y 5,5 kg; machos adultos: 1,9 m y 159 kg; hembras adultas: 1,4 m y 48,5 kg. Estos datos pertenecen a otáridos de las Islas Malvinas (Vaz Ferreira y Vallejo Reference Vaz Ferreira and Vallejo1980). En las costas uruguayas los machos adultos tienen un peso promedio de 80 kg (Schiavini Reference Schiavini1990). Los tamaños promedio de O. flavescens son: crías entre 10,2 m y 14,8 kg; machos adultos: 2,56 m y 300–340 kg (Vaz Ferreira Reference Vaz Ferreira, Ridgway and Harrison1981); hembras adultas: 2 m y 144 kg (Schiavini Reference Schiavini1990).
Los A. australis se ubican sobre costas rocosas y con exposición al oleaje (Crespo et al. Reference Crespo, García, Dans and Pedraza2008; Schiavini Reference Schiavini1990; Sielfeld Reference Sielfeld1983), su distribución no es uniforme en el espacio costero y se concentran en islotes escarpados (Bastida y Rodríguez Reference Bastida and Rodríguez2003; Schiavini Reference Schiavini1990). En Tierra del Fuego, hay alta concentración (Figura 1) en Isla de los Estados y Cabo de Hornos (Crespo et al. Reference Crespo, Schiavini, García, Franco-Trecu, Goodall, Rodríguez, Morgante and Oliveira2015; Milano et al. Reference Milano, Grandi, Schiavini and Crespo2020a; Schiavini y Raya Rey Reference Schiavini and Rey2001), que son sectores de alta productividad marina donde confluyen la Corriente de Malvinas y la Corriente Circumpolar Antártica (Acha et al. Reference Acha, Mianzan, Guerrero, Favero and Bava2004). En el sector interior del canal Beagle, en los islotes Les Éclaireurs se encuentran entre febrero y septiembre (Schiavini y Raya Rey Reference Schiavini and Rey2001), es decir fuera del período reproductivo. Los O. flavescens, eligen costas reparadas con declives suaves, y su distribución geográfica es más continua a lo largo de la costa (Crespo et al. Reference Crespo, García, Dans and Pedraza2008; Milano et al. Reference Milano, Grandi, Schiavini and Crespo2020b; Schiavini Reference Schiavini1990; Schiavini y Raya Rey Reference Schiavini and Rey2001; Sielfeld Reference Sielfeld1983). La porción argentina de Tierra del Fuego aloja un total de 26 colonias y apostaderos (Schiavini et al. Reference Schiavini, Crespo and Szapkievich2004), siendo tres las áreas importantes de cría: las Islas Becasses (canal Beagle), Península Mitre y la Isla de los Estados (Milano et al. Reference Milano, Grandi, Schiavini and Crespo2020b). Nuevamente, es posible observar que los apostaderos reproductivos con mayor número de individuos se encuentran emplazados en espacios exteriores (Milano et al. Reference Milano, Grandi, Schiavini and Crespo2020b). En efecto, en la actualidad ambas especies presentan una distribución diferencial entre los sectores de costa interior y exterior, ya que hacia los espacios de frentes marinos los otáridos aumentan en general, y en particular las hembras adultas e individuos menores de un año, en concordancia con la presencia de apostaderos reproductivos de grandes dimensiones.
Análisis faunísticos previos indican variabilidad en la importancia económica y en las estrategias de aprovisionamiento de otáridos en distintos espacios costeros del archipiélago fueguino (Horwitz Reference Horwitz1990; Lanata Reference Lanata1995; Martinoli Reference Martinoli2017, Reference Martinoli2018; Muñoz Reference Muñoz1996, Reference Muñoz and Goñalons2004, Reference Muñoz2005; Orquera y Piana Reference Orquera and Piana1999a; Vázquez et al. Reference Vázquez, Zangrando, Tessone, Ceraso, Zangrando, Vázquez and Tessone2011; Vidal Reference Vidal1985; Zangrando et al. Reference Zangrando, Tessone, Vázquez, Salemme, Santiago, Álvarez, Piana, Vázquez and Mansur2009). Es posible hallar diferencias en lo que concierne al consumo y la explotación de estas presas entre la porción central de la costa norte del canal Beagle (Martinoli Reference Martinoli2015, Reference Martinoli2018; Orquera y Piana Reference Orquera and Piana1999a; Schiavini Reference Schiavini1990) y la vertiente atlántica de Península Mitre (Muñoz Reference Muñoz1996, Reference Muñoz and Goñalons2004, Reference Muñoz2005), espacios que se articularon a partir de diferentes estrategias de movilidad. En el primer espacio, los otáridos fueron las presas que aportaron el mayor ingreso energético (Orquera y Piana Reference Orquera and Piana1999a; Schiavini Reference Schiavini1990), aunque con variaciones a lo largo del Holoceno (Zangrando Reference Zangrando2009). Asimismo, los restos de la Capa D del sitio Túnel I (mediados del Holoceno medio), muestran el predominio de machos adultos y subadultos de A. australis muertos en invierno (Schiavini Reference Schiavini1990, Reference Schiavini1993). A partir de ello se propuso que estas presas eran capturadas en el sector de canal interior fuera del período reproductivo mediante el uso de técnicas de caza que aumentaban la previsibilidad de su captura, por lo que funcionaban como un subsidio energético desde aguas exteriores hacia aguas interiores (Orquera y Piana Reference Orquera and Piana1999a; Schiavini Reference Schiavini1990, Reference Schiavini1993). Por el contrario, en la costa atlántica de la Isla Grande (litoral norte de Península Mitre), los otáridos no fueron las presas más relevantes, sino los guanacos (Lanata Reference Lanata1995). El perfil etario registrado en otáridos indica el aprovechamiento de individuos inmaduros, lo que se interpretó como parte de una estrategia que buscaba el máximo rendimiento a partir de la predación de los individuos indefensos (Muñoz Reference Muñoz2005), en espacios de agregación en tierra (Lanata Reference Lanata1995; Lanata y Borrero Reference Lanata, Borrero, Lanata and Borrero1994). A su vez, los datos generados para el Holoceno tardío en la costa sur de Península Mitre e Isla de los Estados indican la explotación de ambas especies de otáridos y de todas las categorías de edad y sexo, aunque en porcentajes distintos (Martinoli Reference Martinoli2017, Reference Martinoli2018, Reference Martinoli2019b).
A partir de estos antecedentes, y desde la perspectiva de una densidad menor de otáridos en sectores con una productividad marina menor, se plantean distintas expectativas sobre la forma de explotación de estos mamíferos. En sectores interiores del archipiélago (Beagle y Moat) se espera un predominio de machos adultos y subadultos de ambas especies, es decir individuos no necesariamente asociados a loberías reproductivas. En el caso de los espacios costeros e insulares exteriores (costa sur de Península Mitre e Isla de los Estados) la expectativa es una representación similar de todas las categorías de edad y sexo, y de ambas especies, aunque con una presencia comparativamente mayor (entre espacios interiores y exteriores) de hembras en edad reproductiva y especialmente de individuos menores de un año.
Materiales y Métodos
Se analizan doce conjuntos zooarqueológicos de otáridos provenientes de distintos sectores de la costa sur de Tierra del Fuego y de la Isla de los Estados (Figura 1), los que corresponden a distintas ocupaciones a partir del Holoceno medio (Tabla 1). Para medir la abundancia relativa de las distintas especies y categorías de edad y sexo se utilizaron los indicadores zooarqueológicos usuales: NISP, NME y NMI (Lyman Reference Lyman1994). Se aplicó un criterio amplio para el cálculo del NMI, que incluye (además de la identificación anatómica y de la lateralidad) la determinación de la especie, la edad y el sexo del individuo (Lyman Reference Lyman1994).
Tabla 1. Conjuntos arqueológicos de otáridos analizados. Los datos de las fechas calibradas fueron extraídos de Orquera y Piana (Reference Orquera and Piana2020).

Para la identificación taxonómica se utilizaron rasgos morfológicos del cráneo y del poscráneo (Borella et al. Reference Borella, Vales, Grandi and García2018; Cadegán Sepúlveda Reference Cadegán Sepúlveda2013; Legoupil Reference Legoupil1989; Pérez García Reference Pérez García2003) y colecciones de referencia disponibles en el Museo Acatushún y en el CADIC-CONICET (Ushuaia, Argentina). Entre los primeros consideramos: a) la forma del hueso palatino; b) la forma de la arcada dentaria superior y c) la forma de los maxilares inferiores (Legoupil Reference Legoupil1989). En elementos del poscráneo, la diferenciación se realizó a partir de los huesos de las cinturas de los miembros, del atlas, del axis y del sacro (Pérez García Reference Pérez García2003). Los huesos pertenecientes a la cintura escapular de O. flavescens indican mayor rendimiento muscular plasmado en un tamaño comparativamente mayor de las estructuras óseas (Cadegán Sepúlveda Reference Cadegán Sepúlveda2013). También fueron considerados los rasgos diagnósticos propuestos para: escápula, húmero, cúbito, escafolunar, fémur, tibia/peroné, calcáneo, astrágalo, pelvis, esternebras, sacro y atlas (Borella et al. Reference Borella, Vales, Grandi and García2018; Martinoli Reference Martinoli2018).
Para las categorías de edad utilizamos el esquema preliminar de fusión epifisaria para el esqueleto poscraneal de otáridos (Borella et al. Reference Borella, Grandi, Vales, Goodall, Crespo, Zangrando, Barberena, Gil, Neme, Giardina, Luna, Otaola, Paulides, Salgán and Tivoli2013). A su vez, se emplearon los cuatro estadios ontogénicos propuestos por Pérez García (Reference Pérez García2008). Para la determinación del sexo se observó la forma de los coxales en el isquion y el pubis (King Reference King1983), y el dimorfismo sexual que presentan ambas especies. En el caso de A. australis, se midió el ancho de la corona de los caninos; valores menores/iguales a 8 mm corresponden a hembras, mientras que medidas mayores a ese valor se asignan a machos (Schiavini Reference Schiavini1990). Respecto de O. flavescens el ancho mandibular presenta diferencias entre machos y hembras, con una media de 26,2 mm (±8,9 mm) para los primeros y de 15,6 mm (±3,1 mm) para las segundas (Crespo Reference Crespo1984).
Resultados
En la Tabla 2 se presentan los especímenes identificados a nivel de especie, así como el NMI. El NISP total correspondiente a otáridos ascendió a 14.618, y en el 9,6% pudo ser determinada la especie. Esto no responde a un problema de la integridad de las muestras analizadas: los conjuntos muestran índices bajos de fragmentación, ya que la relación NISP/NME no supera a 2 (Tabla 2). Dicho porcentaje de identificación responde a las dificultades analíticas de diferenciar a nivel de especie a través de elementos post-craneales como costillas, vértebras y articulares, elementos que son significativamente mayoritarios en los conjuntos zooarqueológicos de estos mamíferos (ver Martinoli Reference Martinoli2018). En efecto, si analizamos el grado de identificación a partir de los valores de MNI que no se ven afectados por este factor, el 94,2% (NMI = 163/173) de los individuos identificados pudo ser determinado a nivel de especie.
Tabla 2. Índice de fragmentación general (NISP/NME), NISP, NISP%, NME, NMI y NMI% de otáridos según especie, edad y sexo en conjuntos de la costa sur de Tierra del Fuego e Isla de los Estados.
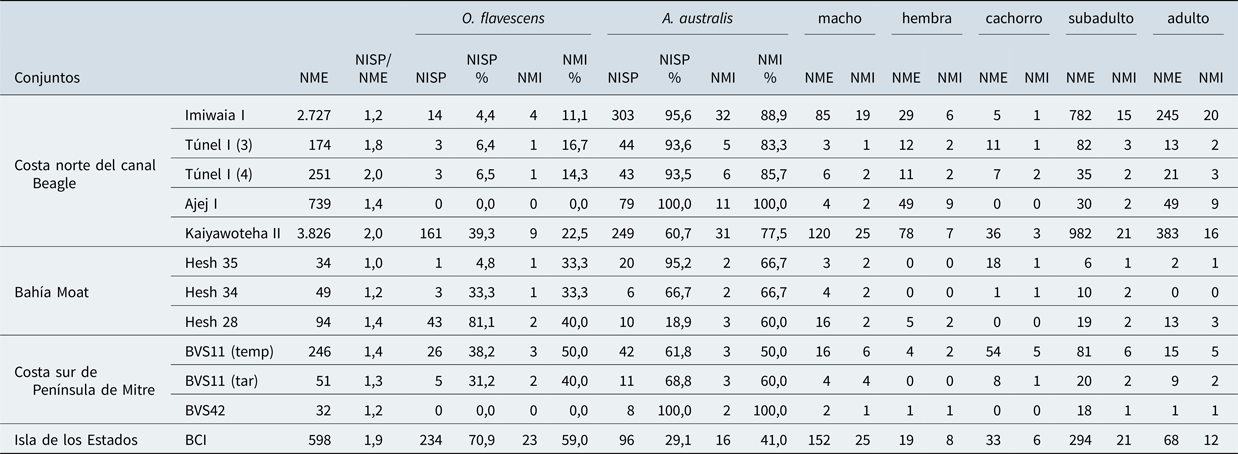
En cuanto a la distribución y abundancia de ambas especies, es posible observar algunas diferencias entre los distintos espacios geográficos (Figura 2). En cuanto a los sectores interiores, tanto en el canal Beagle (NMI% = 87) como en bahía Moat (NMI% = 64,5) la especie más abundante en promedio es A. australis, aunque en el último caso las representaciones de ambas especies se aproximan más entre sí. En la costa sur de Península Mitre continúa el predominio de A. australis (NMI% promedio = 70), incluso se registran casos donde sólo se encuentra representada esta especie (BVS42). En cambio, Isla de los Estados presenta una situación diferente a los conjuntos del sur de Tierra del Fuego dado que el 59% de los individuos identificados en el sitio BC1 corresponden a O. flavescens.

Figura 2. Histograma comparativo de la abundancia de pinnípedos según especie en los conjuntos de la costa sur de Tierra del Fuego e Isla de los Estados.
En lo que concierne a la determinación del sexo de los individuos capturados, es posible hallar variabilidad en cada uno de los sectores geográficos (Tabla 2), aunque con una tendencia general a la mayor presencia de machos. En primer lugar, en el canal Beagle no todos los conjuntos indican la misma relación entre la presencia de machos y hembras, ya que mientras los primeros son predominantes en las Capas K, L y M del sitio Imiwaia I y en Kaiyawoteha II, las hembras predominan en la Capa C de Túnel I y en Ajej I; por su parte, en la Capa α de Túnel I, los machos y las hembras están representadas por un NMI de 2 (Figura 1). En Moat, los machos son los más abundantes en Heshkaia 35 y 34, sin embargo, en Heshkaia 28 los machos y las hembras están presentes de manera equivalente. En la costa sur de Península Mitre, en ambos bloques del sitio Bahía Valentín 11, la categoría más abundante son los machos. En el sitio Bahía Valentín 42 tanto machos como hembras indican una abundancia similar. En Bahía Crossley I, los machos son los individuos más representados. En cuanto a las categorías de edad, también fue posible hallar variabilidad (Tabla 2). En la costa norte del canal Beagle, los adultos son predominantes en las Capas K, L y M de Imiwaia I, en la Capa α de Túnel I y en Ajej I, mientras que los subadultos son abundantes en la Capa C de Túnel I y en Kaiyawoteha II. Los individuos menores a un año están presentes en la Capa α de Túnel I de manera similar a los subadultos. En la localidad de Heshkaia, en términos generales, todas las categorías de edad se encuentran representadas de manera similar. En la costa sur de Península Mitre, en BVS11 (bloque temprano) los adultos y los individuos menores a un año son las categorías más abundantes, mientras que en el bloque tardío de dicho sitio son los adultos y subadultos, al igual que en BVS42. Por último, en BCI la categoría de edad más abundante corresponde a subadultos (ver Martinoli Reference Martinoli2019b:Figura 1).
Si integramos la información correspondiente a las determinaciones taxonómicas, de edad y de sexo es posible tener un panorama completo de las presas capturadas en los distintos espacios del sur de Tierra del Fuego (Tabla 3). En la costa norte del canal Beagle, dos conjuntos presentan perfiles de mortalidad comparables, las Capas K, L y M de Imiwaia I y Kaiyawoteha II: machos adultos y machos subadultos de A. australis representan las categorías más abundantes, mientras el resto de las clases están presentes por debajo del 10% del NMI total. No obstante, en los otros tres conjuntos es posible hallar mayor variabilidad. En primer lugar, si bien en la Capa C de Túnel I los machos subadultos de A. australis son los más representados, el resto de las clases están presentes todas en porcentajes similares (MNI% = 16,7). En segundo lugar, en la Capa α del mismo sitio, las categorías más abundantes son los individuos menores de un año y los subadultos de ambos sexos de A. australis, seguido por machos adultos de ambas especies. En Ajej I las hembras adultas de dicha especie dominan ampliamente el conjunto. En Moat, en todos los conjuntos, las clases taxonómicas de edad y sexo representadas indican porcentajes equivalentes (entre 20% y 35%). En Heshkaia 35 identificamos machos subadultos de O. flavescens, así como machos adultos e individuos menores de un año de A. australis. En Heshkaia 34 las categorías presentes corresponden a machos subadultos de ambas especies, además de individuos menores de un año de A. australis. En Heshkaia 28 se determinó la presencia de machos adultos de O. flavescens, hembras adultas de las dos especies y subadultos de ambos sexos de A. australis. En la costa sur de Península Mitre todas las clases identificadas están presentes en porcentajes similares, es decir ninguna predomina sobre otra (Tabla 3). En el bloque temprano de BVS11 fue determinada la presencia de machos adultos de ambas especies, hembras subadultas e individuos menores de un año de O. flavescens, así como hembras adultas e individuos menores de un año de A. australis. En el bloque tardío del mismo sitio identificamos machos adultos y subadultos de ambas especies e individuos menores de un año de A. australis. En BVS42, sólo se hallaron dos individuos de A. australis: una hembra adulta y un macho subadulto (Tabla 3). Por último, en el sitio de Isla de los Estados, la categoría más abundante corresponde a machos subadultos de O. flavescens, seguido por machos adultos de la misma especie, machos subadultos de A. australis, hembras subadultas de O. flavescens e individuos menores de un año de ambas especies (Tabla 3).
Tabla 3. Perfiles de mortalidad de los conjuntos de otáridos recuperados en el sur de Tierra del Fuego e Isla de los Estados.

Discusión
Desde hace más de tres décadas los parámetros ecológicos, el comportamiento y estructura de edad y sexo de poblaciones actuales de otáridos son las variables mayormente empleadas para evaluar las modalidades de explotación de esas presas por grupos cazadores-recolectores de América durante el Holoceno (Borella Reference Borella2006; Lyman Reference Lyman1989, Reference Lyman1995; Muñoz Reference Muñoz, Bicho, Haws and Davis2011; Schiavini Reference Schiavini1993; Zangrando Reference Zangrando2009; entre otros). En este trabajo tomamos estos criterios para generar expectativas arqueológicas sobre la explotación de otáridos en diferentes ámbitos del sur de Tierra del Fuego, considerando que nuestra área de estudio nos permite analizar comparativamente espacios que presentan diferentes condiciones para su captura.
La información zooarqueológica proporcionada en este trabajo muestra un panorama diferente al esperado a partir de la distribución actual de los apostaderos reproductivos. En principio, se destaca la presencia de individuos menores de un año de A. australis en seis de los 11 conjuntos analizados del canal Beagle, Moat y Península Mitre con proporciones superiores al 16% del número mínimo de los ejemplares identificados. Si a este parámetro sumamos además la representación de hembras adultas (Figura 3), en siete de los 11 conjuntos analizados los ejemplares correspondientes a hembras adultas e individuos menores de un año superan el 20%. A su vez, no se observa una mayor representación en esas categorías de sexo y edad en Isla de los Estados. La representación de O. flavescens en conjuntos zooarqueológicos del canal Beagle, Moat y sur de Península Mitre es significativamente menor a la de A. australis, con la excepción de algunos sitios (por ejemplo Heshkaia 28). Sin embargo, en Bahía Crossley I esta relación se invierte (Figura 2). Por su parte, la abundancia de individuos menores de un año de O. flavescens sólo es importante en el bloque temprano de BVS11, y se observan hembras adultas en sitios del canal Beagle y Moat. En resumen, los resultados aquí presentados señalarían que parte de la explotación de otáridos podría haber ocurrido sobre apostaderos reproductivos en todos los ámbitos del archipiélago fueguino, a excepción de Isla de los Estados: los perfiles de edad y sexo de O. flavescens indicarían un aprovisionamiento mayormente centrado en individuos generalmente desagregados de los espacios reproductivos. Esto discrepa con las expectativas planteadas en función de la distribución y comportamiento actuales de los otáridos en la región y plantea la revisión de algunas interpretaciones previas.

Figura 3. Proporción de categorías de edad y sexo de pinnípedos (NMI%) representados en los distintos espacios que conforman la costa sur de Tierra del Fuego e Isla de los Estados.
En primer lugar, debemos considerar los métodos aplicados para la determinación de las crías en período de lactancia de A. australis y O. flavescens. Los esquemas preliminares de fusión epifisaria, si bien pueden ser útiles para determinar si un individuo es menor de un año, no necesariamente indican que el animal se está aún amamantando, ya que los ritmos de destete son graduales y pueden presentar variaciones (Heckel y Schramm Reference Heckel, Schramm, Heckel and Schramm2021). Los estudios de isotopos estables constituyen una vía complementaria de evaluación. En las crías de A. australis y O. flavescens la lactancia es de aproximadamente un año y es posible esperar valores de δ15N entre 2‰ y 3‰ más enriquecidos que en hembras adultas de la misma especie (Lima de Albernaz et al. Reference Lima de Albernaz, Secchi, de Oliveira and Botta2017). La Figura 4 muestra la correlación entre valores de δ13C y δ15N para muestras arqueológicas de hembras adultas (círculos blancos) y crías (círculos grises) procedentes de diferentes sitios del sur de Tierra del Fuego (Nye et al. Reference Nye, Zangrando, Martinoli and Fogel2020, Tabla A1). Se observó que sobre un total de 10 crías identificadas a partir de los criterios previamente señalados, sólo cuatro presentan dietas compatibles con individuos lactantes, el resto de los ejemplares se encuentran dentro del rango de valores de hembras adultas, es decir que deben considerarse ejemplares juveniles, los que pueden estar asociados tanto a colonias reproductivas como de descanso (Heckel y Schramm Reference Heckel, Schramm, Heckel and Schramm2021). Los estudios de isótopos estables ofrecen parámetros sobre los comportamientos de los otáridos en el pasado que pueden ser articulados con procedimientos zooarqueológicos tradicionales (Zangrando et al. Reference Zangrando, Panarello and Piana2014) y ajustar así nuestras interpretaciones sobre las modalidades de explotación de estos mamíferos. La complementación de análisis de isótopos estables con estimaciones morfométricas en A. australis y O. flavescens (Borella y L'Heureux Reference Borella and L'Heureux2014; Borella et al. Reference Borella, L'Heureux and Grandi2014; L'Heureux y Borella Reference L'Heureux and Borella2011) resulta particularmente sugestiva para este propósito.

Figura 4. Valores isotópicos de δ13C y δ15N en individuos determinados como crías (círculos grises) y hembras adultas (círculos blancos) de A. australis en conjuntos de la costa sur de Tierra del Fuego e Isla de los Estados.
Aunque algunos de los individuos menores de un año, presentes en los conjuntos del canal Beagle y de Moat, no se encontraban lactando al momento de su captura, en muchos de los conjuntos provenientes de sectores interiores hay una elevada presencia de hembras adultas, mayor a la esperada según la distribución actual de apostaderos reproductivos. Por lo cual, es posible que parte de las capturas de otáridos llevada a cabo en esos espacios se haya realizado efectivamente en espacios de colonias de descanso y/o reproductivas, considerando en conjunto la presencia de hembras adultas, de juveniles (menores de un año) y de algunos animales lactantes. A este respecto, es posible pensar que al menos la tasa de encuentro de estas categorías de edad y sexo de A. australis en sectores interiores del archipiélago fueguino puede haber sido mayor durante el Holoceno. En este punto, es necesario recordar que desde finales del siglo dieciocho hasta mediados del siglo veinte, los lobos marinos sufrieron explotación industrial, lo que diezmó las poblaciones de otáridos en el ámbito fuego-patagónico (Bonner Reference Bonner1982; Carrara Reference Carrara1952; Orquera Reference Orquera, Briones and Lanata2002; Weddell Reference Weddell2006 [1825]). Si bien hasta el momento no se registraron relocalizaciones de colonias de reproducción de O. flavescens y A. australis desde los censos de Carrara (Reference Carrara1952; Milano et al. Reference Milano, Grandi, Schiavini and Crespo2020a, Reference Milano, Grandi, Schiavini and Crespo2020b; Túnez et al. Reference Túnez and Humberto Cappozzo2008), tampoco es posible descartar la posibilidad de que hayan ocurrido cambios en las localizaciones de colonias (Martinoli Reference Martinoli2018; Martinoli et al. Reference Martinoli, Crespo, Dellabianca and Zangrando2021a; Vidal y Winograd Reference Vidal and Winograd1986) como consecuencia de variaciones ambientales de corto plazo (Saporiti et al. Reference Saporiti, Bala, Otero, Crespo, Piana, Aguilar and Cardona2014). Esto puede haber implicado la presencia de colonias reproductivas de pequeñas dimensiones en aguas de canales interiores con menor productividad marina y/o un número mayor de colonias de descanso de A. australis. De todos modos, el estado actual de los estudios en δ13C y δ15N sobre muestras zooarqueológicas procedentes de la región, si bien marcan variaciones en los nichos isotópicos de otáridos a lo largo del Holoceno (Nye et al. Reference Nye, Zangrando, Martinoli and Fogel2020; Vales et al. Reference Vales, Cardona, Zangrando, Borella, Saporiti, Goodall, de Oliveira and Crespo2017; Zangrando et al. Reference Zangrando, Panarello and Piana2014), estos no se vinculan necesariamente con cambios geográficos en la localización de las grandes colonias de reproducción de otáridos.
Un segundo aspecto para considerar es el predominio de machos adultos y subadultos en el conjunto proveniente de Isla de los Estados, siendo que este sitio se encuentra emplazado en las cercanías de las loberías reproductivas de A. australis más grandes de Patagonia sur, sobre el borde de la plataforma continental con alta productividad marina, situación geográfica que era similar en el momento de las ocupaciones aquí estudiadas de la isla (Ponce Reference Ponce2009). Por lo cual, es necesario considerar algunos puntos. Por un lado, la formación de los conjuntos arqueofaunísticos puede promediar diferentes episodios de captura de otáridos a partir de la explotación de diferentes tipos de apostaderos (Muñoz Reference Muñoz, Bicho, Haws and Davis2011; Muñoz et al. Reference Muñoz, Cruz, Lemaire, Pretto, Zangrando, Barberena, Gil, Neme, Giardina, Luna, Otaola, Paulides, Salgán and Tivoli2013). Por otro lado, la estructura de los conjuntos estudiados muchas veces se encuentra mediada por procesos tafonómicos en sentido amplio (Lyman Reference Lyman2003; Borella y Borrero Reference Borella and Borrero2019). La evidencia recuperada en bahía Tethis y generada durante la explotación industrial de otáridos en un período de pocos años en Península Mitre, ilustra el alcance que los factores tafonómicos pueden tener en nuestras interpretaciones a partir de las categorías de sexo y edad. La factoría de bahía Tethis funcionó entre 1940 y 1948 y la información histórica indica que la explotación se focalizaba en dos apostaderos reproductivos: Las Chapas y Caleta Tres Amigos (Vázquez et al. Reference Vázquez, Fernando and Zangrando2019). Las carcasas eran descartadas en la bahía, dando lugar a extensas acumulaciones de huesos en sus costas. El estudio zooarqueológico (Vázquez et al. Reference Vázquez, Fernando and Zangrando2019) de estos conjuntos señala que la explotación se focalizó en O. flavescens y con una representación mayor de machos (80%) sobre hembras (20%). Los elementos correspondientes a crías no superan el 5%. Estas proporciones no se corresponden con las esperadas para colonias de reproducción (Milano et al. Reference Milano, Grandi, Schiavini and Crespo2020b). Si bien existen razones para creer que las crías no eran trasladadas hasta la factoría en igual magnitud que ejemplares adultos y subadultos (ver Vázquez et al. Reference Vázquez, Fernando and Zangrando2019), la baja proporción de hembras resulta igualmente llamativa. Es probable que esto responda a que la selección de ejemplares haya tenido un sesgo hacia individuos de mayor tamaño, de manera independiente de la composición social de los apostaderos, así como el uso de los huesos para tareas específicas, como por ejemplo combustible. Además, es posible la pérdida de aquellos elementos correspondientes a individuos juveniles y cachorros, de pequeñas dimensiones y/o de baja densidad mineral ósea (Borella y Borrero Reference Borella and Borrero2019; Martinoli et al. Reference Martinoli, Gutiérrez and Zangrando2021b). Este ejemplo, donde se encuentran documentadas las actividades de explotación de un tipo particular de apostadero, muestra que el alcance de las predicciones basadas únicamente en la estructura de edad y sexo de colonias actuales debe ser manejado con cautela, en tanto los conjuntos zooarqueológicos presentan diferentes trayectorias de formación. En efecto, las proporciones por sexo, como las registradas en las acumulaciones de huesos de bahía Tethis, fueron interpretadas como estrategias de explotación marcadamente diferentes en otros contextos del archipiélago fueguino (ver Schiavini Reference Schiavini1993).
Conclusión
En términos generales, y al contrario de las expectativas generadas a partir del marco ecológico que ofrece la distribución actual de los otáridos en el archipiélago fueguino y de los antecedentes arqueológicos, es posible observar similitud en muchos de los perfiles de mortalidad determinados en la costa norte del canal Beagle, en Moat y en la costa sur de Península Mitre. En el caso de los espacios de canales interiores, las estrategias de aprovisionamiento de otáridos señalan una mayor heterogeneidad respecto del modelo propuesto previamente para el área (Schiavini Reference Schiavini1990). Es decir, en función de la abundancia de hembras adultas, lactantes e individuos menores de un año en sectores interiores del archipiélago, no es posible descartar la explotación de áreas de agregación (reproductivos y/o descanso) de A. australis. Lo dicho implica que puede haber habido variaciones locales en la abundancia y distribución de colonias reproductivas y de descanso de otáridos en los canales interiores, las que pueden haberse visto afectadas por la explotación industrial llevada a cabo durante los siglos dieciocho y diecinueve (Vales et al. Reference Vales, Cardona, Zangrando, Borella, Saporiti, Goodall, de Oliveira and Crespo2017).
Por su parte, el perfil de mortalidad en Bahía Crossley I, dominado por machos adultos y subadultos, indicaría que la captura de otáridos se articuló a partir de una estrategia centrada en individuos desagregados de espacios reproductivos. Sin embargo, como indican los resultados obtenidos en bahía Tethis, la presencia mayoritaria de individuos machos adultos y subadultos, no necesariamente descarta la posibilidad de que se hayan explotado espacios reproductivos, sino que habla de la selección de presas (relacionada posiblemente con su tamaño), así como de la intervención de posibles factores posdepositacionales que pueden haber afectado la abundancia de individuos jóvenes, así como de huesos pequeños y/o de baja densidad mineral ósea (Borrero y Borella Reference Borella and Borrero2019; Martinoli Reference Martinoli2019b; Martinoli et al. Reference Martinoli, Gutiérrez and Zangrando2021b).
En resumen, es posible que las estrategias de aprovisionamiento de otáridos hayan sido similares en todo el sur de Tierra del Fuego. Lo dicho alerta sobre la importancia de realizar análisis que incluyan la consideración de distintos factores vinculados a la formación de conjuntos arqueológicos, así como a los aspectos relacionados con la paleoecología y comportamiento de los otáridos, la selección de presas y los posibles cambios ambientales antrópicos y no antrópicos.
Agradecimientos
Este trabajo se realizó en el marco del PICT 2017-1230. Agradecemos a Luis A. Orquera y a Ernesto L. Piana por darnos acceso a los conjuntos provenientes del canal Beagle. A Florencia Borella por los valiosos comentarios realizados al manuscrito. Agradecemos a los evaluadores anónimos por sus sugerencias.
Declaración de Financiamiento
Este trabajo se financió con el proyecto PICT 2017-1230 -Agencia Nacional de Promoción Científica y Tecnológica: “Dinámica de poblaciones humanas y paisajes arqueológicos costeros en el litoral sur de Tierra del Fuego durante el Holoceno”, dirigido por el Dr. Atilio Francisco Zangrando.
Declaración de disponibilidad de datos
Repositorio institucional Centro Austral de Investigaciones Científicas (CADIC-CONICET). Bernardo Houssay 200, Ushuaia, Tierra del Fuego, Argentina.
Conflicto de intereses
Los autores declaran no tener conflicto de intereses.







