No CrossRef data available.
Published online by Cambridge University Press: 17 October 2024
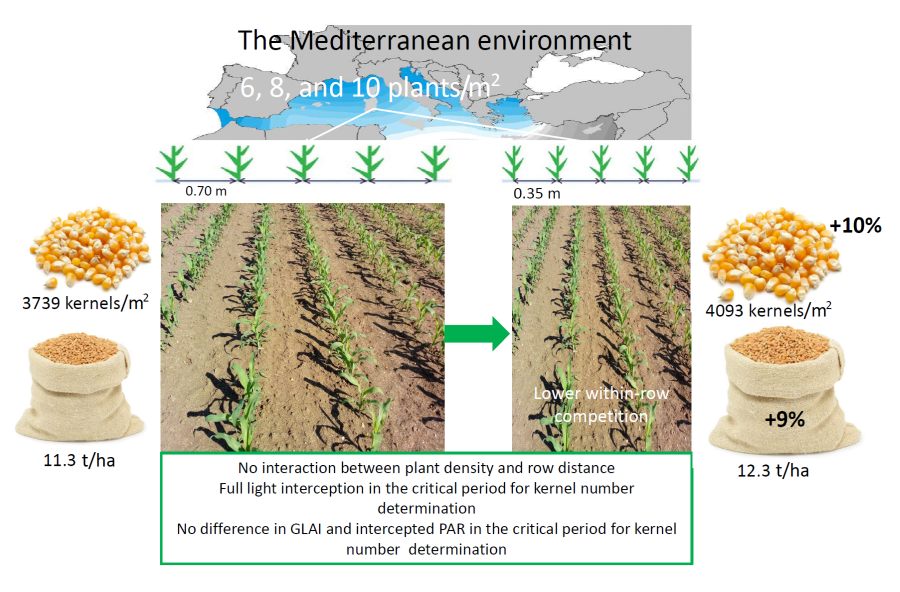
The spatial distribution of maize plants influences the number of kernels set per plant by modifying the radiation environment during the critical period for kernel number determination. A two-year field experiment was conducted in Sardinia to explore whether, in a Mediterranean environment, a reduction in row distance can have a positive effect on grain yield of full-season maize hybrids and, if it does, whether this effect is also affected by plant density. Treatments were a factorial combination of three plant densities (6, 8 and 10 plants/m2) and two row spacings (35 and 70 cm). Grain yield and grain number benefited from a reduction in row distance from the canonical 70 cm to 35 cm irrespective of plant density and yield level (14.7 t/ha on average in 2021, 9.9 t/ha in 2022 vs 13.7 and 8.8 t/ha with canonical row distance). Row distance did not affect either the coefficient of extinction of radiation or the intercepted photosynthetically active radiation (PAR) during the critical period for grain number determination. The higher grain number per unit area at the smaller row distance translated into a greater grain yield, which cannot solely be attributed to radiation interception and plant growth rate during the period of kernel number determination. Halving the common row distance is a valuable management option for full-season hybrids grown in Mediterranean environments without nitrogen and water limitations as it seems capable of increasing grain yield via an increase in kernel number per unit area, regardless of plant density and yield level.