



No CrossRef data available.
Article contents
Impact of crown architecture on light availability, gas exchange, flowering and fruiting in jamun (Syzygium cumini [L.] Skeels)
Published online by Cambridge University Press: 27 November 2024
Abstract
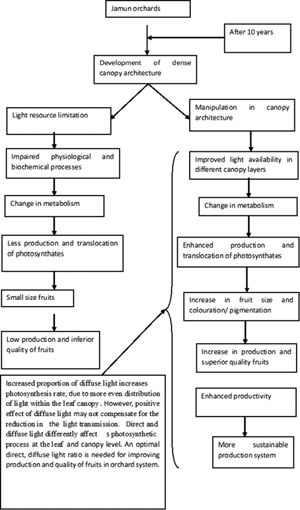
Experiments were conducted to assess the impact of crown architecture on light availability beneath the trees, flowering, fruiting, yield and quality of jamun (Syzygium cumini [L.] Skeels). Trees were maintained as control, palmette and open centre crown. Impact was evaluated for three consecutive years, i.e. 2017–2019. Diffuse light beneath the trees ranged from 69.7 ± 2.22 to 45.9 ± 1.45%, whereas direct light varied from 30.4 ± 0.97 to 54.1 ± 1.78%. At flowering and fruit development stage (June), photosynthesis rate (A) in control trees was 12.5 ± 0.43 μmol CO2/m2/s; however, at fruit maturity and dormancy (August), it was only 9.5 ± 0.35 μmol CO2/m2/s. Similarly, in palmette and open centre trees, photosynthesis rate at flowering and fruit development stage was 13.5 ± 0.46 and 15.7 ± 0.54 μmol CO2/m2/s, respectively; whereas at fruit maturity and dormancy, photosynthesis rate dropped to 10.5 ± 0.39 and 11.7 ± 0.43 μmol CO2/m2/s, respectively. Substantial variation in stomatal conductance (gs), vapour pressure deficit (VPD) and transpiration rate (E) was also found. Days to start flowering ranged from 92 ± 0.33 to 98 ± 0.33. Similarly, days to end flowering varied from 99 ± 0.07 to 107 ± 0.36, days to fruit set 132 ± 0.33 to 139 ± 0.33 and days to fruit maturity 176 ± 0.48 to 184 ± 0.63. Significant variation in fruit length, fruit width and fruit weight was also found. Total soluble solids in fruit pulp varied from 9.0 ± 0.15 to 12.2 ± 0.149°Brix and fruit yield 62.3 ± 1.5 to 86.7 ± 1.33 kg per tree. Noteworthy variation in fruit quality traits was also recorded. This study illustrates that crown architecture has considerable impact on gas exchange parameters, flowering, fruiting, yield and quality of jamun.
- Type
- Crops and Soils Research Paper
- Information
- Copyright
- Copyright © INDIAN COUNCIL OF AGRICULTURAL RESEARCH, 2024. Published by Cambridge University Press
References
Abdul-Hamid, A and Luan, YS (2000) Functional properties of dietary fibre prepared from defatted rice bran. Food Chemistry 68, 15–19.CrossRefGoogle Scholar
Acreche, MM, Briceno-Felix, G, Sanchez, JAM and Slafer, GA (2009) Radiation interception and use efficiency as affected by breeding in Mediterranean wheat. Field Crops Research 110, 91–97.CrossRefGoogle Scholar
Association of Official Analytical Chemists (2005) Methods 2001.03 Dietary fiber in foods containing resistant maltodextrin. In Horwithz, W (ed.), Official Methods of Analysis of AOAC International, 18th Edn. Gaithersburg, MD: AOAC International, p 44–54.Google Scholar
Bagchi, DC, Sen, K, Bagchi, M and Atalay, M (2004) Anti-angiogenic, antioxidant and anti-carcinogenic properties of a novel anthocyanin-rich berry extract formula. Biochemistry 69, 75–80.Google ScholarPubMed
Benvenuti, S, Pellati, F, Melegari, M and Bertelli, D (2006) Polyphenols, anthocyanins, ascorbic acid, and radical scavenging activity of Rubus, Ribes, and Aronia. Journal of Food Science 69, 164–169.Google Scholar
Bird, R and Riordan, C (1986) Simple solar spectral model for direct and diffuse irradiance on horizontal and tilted planes at the earth's surface for cloudless atmospheres. Journal of Climate and Applied Meteorology 25, 87–97.2.0.CO;2>CrossRefGoogle Scholar
Bodin, P and Franklin, O (2012) Efficient modeling of sun/shade canopy radiation dynamics explicitly accounting for scattering. Geoscientific Model Development 5, 535–541.CrossRefGoogle Scholar
Brodersen, C and Vogelmann, T (2007) Do epidermal lens cells facilitate the absorptance of diffuse light? American Journal of Botany 94, 1061–1066.CrossRefGoogle ScholarPubMed
Brodersen, CR, Voglmann, TC, Williams, WE and Gorton, HL (2008) A new paradigm in leaf-level photosynthesis: direct and diffuse lights are not equal. Plant, Cell and Environment 31, 159–164.CrossRefGoogle Scholar
Byers, RE, Carbaugh, DH, Presley, CN and Wolf, TK (1991) The influence of low light on apple fruit abscission. Journal of Horticultural Science 66, 7–17.CrossRefGoogle Scholar
Calvo-Rodriguez, S and Sanchez-Azofeifa, GA (2016) Light diffusion in the tropical dry forest of Costa Rica. In The International Archives of the Photogrammetry, Remote Sensing and Spatial Information Sciences, Volume XLI-B8, 2016 XXIII ISPRS Congress, 12–19 July 2016, Prague: Czech Republic.CrossRefGoogle Scholar
Cheng, GW and Breen, PJ (1991) Activity of phynylalanine ammonia-lyase (PAL) and concentrations of anthocyanins and phenolics in developing strawberry fruit. Journal of the American Society for Horticultural Science 116, 865–869.CrossRefGoogle Scholar
Davis, SHR and Masten, SJ (1991) Spectrophotometric method for ascorbic acid using dichlorophenolindophenol: elimination of the interference due to iron. Analytica Chemica Acta 248, 225–221.CrossRefGoogle Scholar
Earles, JM, Théroux-Rancourt, G, Gilbert, ME, McElrone, AJ and Brodersen, CR (2017) Excess diffuse light absorption in upper mesophyll limits CO2 drawdown and depresses photosynthesis. Plant Physiology 174, 1082–1096.CrossRefGoogle ScholarPubMed
Geider, RJ, Delucia, EH, Falkowski, PG, Finzi, AC, Grime, JP, Grace, J, Kana, TM, Roche, JL, Long, SP, Osborne, BA, Platt, T, Prentice, IC, Raven, JA, Schlesinger, WH, Smetacek, V, Stuart, V, Sathyendranath, S, Thomas, RB, Vogelmann, TC, Williams, P and Woodward, FI (2001) Primary productivity of planet earth: biological determinants and physical constraints in terrestrial and aquatic habitats. Global Change Biology 7, 849–882.CrossRefGoogle Scholar
Ghosh, P, Pradhan, RC, Mishra, S, Patel, AS and Kar, A (2017) Physicochemical and nutritional characterization of jamun (Syzygium cumini). Current Research in Nutrition and Food Science 5, 25–35.CrossRefGoogle Scholar
Gorton, H, Williams, W and Vogelmann, T (1999) Chloroplast movement in Alocasia macrorrhiza. Physiologia Plantarum 106, 421–428.CrossRefGoogle Scholar
Jackson, JE (1980) Light interception and utilization by orchard systems. Horticultural Reviews 2, 208–267.CrossRefGoogle Scholar
Jones, JR, Lineback, DM and Levine, MJ (2006) Dietary reference intakes: implications for fibre labelling and consumption: a summary of the international life sciences institute. North America fibre workshop, June 1–2, 2004, Washington, DC. Nutrition Reviews 64, 31–38.CrossRefGoogle ScholarPubMed
Li, T, Heuvelink, E, Dueck, TA, Janse, J, Gort, G and Marcelis, LFM (2014) Enhancement of crop photosynthesis by diffuse light: quantifying the contributing factors. Annals of Botany 114, 145–156.CrossRefGoogle ScholarPubMed
Li, T, Kromdijk, J, Heuvelink, E, van Noort, FR, Kaiser, E and Marcelis, LFM (2016) Effects of diffuse light on radiation use efficiency of two anthurium cultivars depend on the response of stomatal conductance to dynamic light intensity. Frontiers in Plant Science 7, 56. doi: 10.3389/fpls.2016.00056Google ScholarPubMed
Maddonni, GA, Otegui, ME and Cirilo, AG (2001) Plant population density, row spacing and hybrid effects on maize canopy architecture and light attenuation. Field Crops Research 71, 183–193.CrossRefGoogle Scholar
Mishra, DS, Singh, A, Kumar, R, Singh, S, Singh, AK and Swamy, GSK (2014) Jamun. In Ghosh, SN (ed.), Tropical and Sub Tropical Fruit Crops: Crop Improvement and Varietal Wealth Part-2. New Delhi: Jaya Publishing House, pp. 375–390.Google Scholar
Niinemets, U (2007) Photosynthesis and resource distribution through plant canopies. Plant Cell and Environment 30, 1052–1071. doi: 10.1111/j.1365-3040.2007.01683.x.CrossRefGoogle ScholarPubMed
Norman, J and Arkebauer, T (1991) Predicting canopy light use efficiency from leaf characteristics. Agronomy Monographs 31, 125–143.Google Scholar
Norman, J and Miller, E (1971) Light intensity and sunfleck size distributions in plant canopies. Agronomy Journal 63, 743–748.CrossRefGoogle Scholar
Periyathambi, R (2007) Jamun - The Potential Untapped. In Kerala Calling, February 2007 pp 30–32. Public Relations Department, Govt. of. Kerala, Thiruvananthapuram (India).Google Scholar
Sarlikioti, V, de Visser, PH, Buck-Sorlin, GH and Marcelis, LFM (2011) How plant architecture affects light absorption and photosynthesis in tomato: towards an ideotype for plant architecture using a functional–structural plant model. Annals of Botany 108, 1065–1073.CrossRefGoogle ScholarPubMed
Singh, AK, Bajpai, A, Singh, A and Reddy, BMC (2007) Evaluation of variability in jamun using morphological and physico-chemical characterization. Indian Journal of Agricultural Sciences 77, 845–848.Google Scholar
Slinkard, K and Singleton, VL (1977) Total Phenol Analysis: Automation and Comparison with Manual Methods. American Journal of Enology and Viticulture 28, 49–55.CrossRefGoogle Scholar
Sultan, SE (2000) Phenotypic plasticity for plant development, function and life history. Trends in Plant Science 5, 537–542. doi: 10.1016/S1360-1385(00)01797-0.CrossRefGoogle ScholarPubMed
Urban, O, Janous, D, Acosta, M, Czerny, R, Markova, I, Navrátil, M, Pavelka, M, Pokorný, R, Šprtová, M, Zhang, R, Špunda, V, Grace, J and Marek, M (2007) Ecophysiological controls over the net ecosystem exchange of mountain spruce stand, comparison of the response in direct vs. diffuse solar radiation. Global Change Biology 13, 157–168.CrossRefGoogle Scholar
Warrier, PK, Nambiar, VPK and Ramankutty, C (eds) (1996) Indian Medicinal Plants: A Compendium of 500 Species, Vol. 5. Hyderabad: Orient Longman Ltd., pp. 225–228.Google Scholar
Warrington, IJ, Stanley, CJ, Tustin, DS, Hirst, PM and Cashmore, WM (1996) Light transmission, yield distribution, and fruit quality in six tree canopy forms of ‘Granny Smith’ apple. Journal of Tree Fruit Production 1, 27–54. doi: 10.1300/J072v01n0103CrossRefGoogle Scholar
Williams, W, Gorton, H and Witiak, S (2003) Chloroplast movements in the field. Plant, Cell and Environment 26, 2005–2014.CrossRefGoogle Scholar
Williams, M, Rastetter, EB, Pol, LV and Shaver, GR (2014) Arctic canopy photosynthetic efficiency enhanced under diffuse light, linked to a reduction in the fraction of the canopy in deep shade. New Phytologist 202, 1266–1276.CrossRefGoogle ScholarPubMed
Yue, X and Xu, Z (2008) Changes of anthocyanins, anthocyanidins, and antioxidant activity in bilberry extract during dry heating. Journal of Food Science 73, C494–C499.CrossRefGoogle ScholarPubMed