


Background
In 1942, Conrad Waddington coined the term ‘epigenetics’ (literally ‘above genetics’) and defined it as ‘interactions of genes with their environment that bring the phenotype into being’ (Waddington, Reference Waddington1942). But it wasn't until 60 years later that evidence for the environment-epigenetic link was provided (Cooney et al., Reference Cooney, Dave and Wolff2002) and research on this issue has accelerated ever since. We now know that small molecular marks such as the methyl group (CH3) can modify DNA or its packaging proteins and influence the structure and function of chromosome regions, and that such changes can persist for many cell generations after an initial environmental-induced change (Bird, Reference Bird2007). During pre- and early post-natal development, it is thought that the epigenome, the sum of a genome's epigenetic marks, is particularly susceptible to environmental influence because of its high rate of change (Gluckman et al., Reference Gluckman, Hanson, Buklijas, Low and Beedle2009). However, it is far from clear how this susceptibility changes throughout the lifespan, although twins may epigenetically drift apart during the life course (Fraga et al., Reference Fraga, Ballestar, Paz, Ropero, Setien, Ballestar and Esteller2005).
Recruitment Procedures and Brief History of the Cohort and Samples Collected
PETS is a cohort of 250 women recruited in the second trimester of a twin pregnancy from three Melbourne hospitals. It was established primarily to investigate the involvement of epigenetics in the ‘Developmental Origins of Health and Disease’ (DOHaD) paradigm, in which early life environment influences risk of later chronic disease (Gluckman et al., Reference Gluckman, Hanson and Beedle2007). This field began with a review by David Barker, subtitled ‘The womb may be more important than the home’, in which he highlighted his findings of links between low placenta and birth weight and current blood pressure and risk of hypertension in 449 fifty-year-olds (Barker, Reference Barker1990).
On recruitment and at 24 and 36 weeks of pregnancy, questionnaires were administered to women on pre- and post-conceptional diet and lifestyle, stress during pregnancy, and obstetric issues including assisted reproduction (Saffery et al., Reference Saffery, Morley, Carlin, Joo, Ollikainen, Novakovic and Craig2011). In late pregnancy, serum and plasma was collected. A defining feature of PETS is that we optimized contact with mothers and with maternity units to maximize the chance of having PETS staff present at the births. We collected multiple tissue samples (placentae, cords, and cord blood) from vaginal and cesarean births and transported to the laboratory, mostly within 2 hours, to optimize storage for future analysis of DNA, RNA, and epigenetic marks. Human umbilical vein endothelial cells (HUVECs) were processed from cords; leucocytes (total white blood cells), mononuclear cells (CBMCs), granulocytes, serum, and plasma were processed from blood. Cheek swabs were collected from 91.4% of babies within 2 days of birth, including from the 25% of babies whose birth we missed. For the latter, we asked for consent to access neonatal ‘Guthrie’ dried blood spots, which enabled us to collect whole blood (leucocyte) equivalent from 98% of the cohort at, or close to, birth. We were the first to demonstrate the utility of using Guthrie cards to analyze DNA methylation (Wong et al., Reference Wong, Morley, Saffery and Craig2008), an important step for the study of human disease, allowing the distinction between cause and consequence of disease-associated epigenetic differences (Editorial, 2012). We have HUVECs, cords, and cheek cells from >85% of twins (Saffery et al., Reference Saffery, Morley, Carlin, Joo, Ollikainen, Novakovic and Craig2011; see Table 1). We reviewed mothers and twins again at 18 months of age and collected data on the nutrition and health of twins since birth. We took repeat cheek swabs and blood samples, where consent was provided, and anthropometric measurements including weight, length, head circumference, and skinfold thickness.
TABLE 1 PETS Numbers and Samples Indicating the Proportion of Questionnaire Data and Biological Specimens Collected, Includes Frozen Whole Blood and Dried Neonatal Blood Spots

1Twins/whole blood
Assessment of Chorionicity and Zygosity
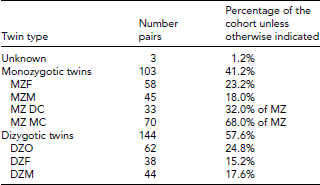
Knowledge of chorionicity is paramount to determine perinatal risks of mortality and morbidity and also correlates with post-natal health outcomes. Monochorionic (MC) twins have poorer outcomes than dichorionic (DC) twins, mostly due to in utero twin–twin transfusion syndrome, the pathological extreme of blood sharing due to vascular connections within the same placenta, which occurs in most MC twin pairs (Hack et al., Reference Hack, Derks, Elias, Franx, Roos, Voerman and Visser2008; Hatkar & Bhide, Reference Hatkar and Bhide1999; Leduc et al., Reference Leduc, Takser and Rinfret2005; Rao et al., Reference Rao, Sairam and Shehata2004). We recorded two measures of chorionicity: from first trimester ultrasound images (Carroll et al., Reference Carroll, Soothill, Abdel-Fattah, Porter, Montague and Kyle2002; D'Alton & Dudley, Reference D'Alton and Dudley1989) and from physical examination of the inter-placental membranes at birth. The latter was performed by trained PETS staff by a relatively simple procedure. DC placentae were separated by a thick layer of 3–4 visible membranes; MC placentae by a thin, almost transparent layer of 1–2 visible membranes. To determine zygosity, MC twins were assumed to be monozygotic (MZ), different-sex twins as dizygotic (DZ), and in same-sex DC twins, zygosity was determined by a 12-marker micro-satellite test (Becker et al., Reference Becker, Busjahn, Faulhaber, Bahring, Robertson, Schuster and Luft1997) using DNA from cord and/or buccal samples. DZ twins generally differed in size at 4–19/24 (median 13/24) alleles. Twins in which initial zygosity results indicated 1–2/24 allele differences or >2 sizes at individual alleles were all found to be MZ on retesting, indicating a low frequency of minor errors using this method. All mothers were sent a letter with zygosity conclusions. Three of the 70 mothers of MC twins contacted us asking to conduct a zygosity test due to the perceived discordance of their twins. In each case, the twins were confirmed as MZ. One mother wrote at 18 months: ‘We were very surprised when [the Research Nurse] rang at 7 months of age and said they were identical. Since then they have become more similar and look identical’. It is also worth noting that in rare cases, DZ twins can also be MC (Umstad et al., Reference Umstad, Short, Wilson and Craig2012) although we did not assess this. A breakdown of PETS twins by zygosity, chorionicity, and sex is provided in Table 2. In summary, approximately two-fifths are MZ and approximately one-third of these are DC.
TABLE 2 Breakdown of Numbers of Twins in the PETS Cohort at Birth
Notes
Unknown = zygosity unknown; MZF = monozygotic female twins; MZM = monozygotic male twins; MZ DC = monozygotic dichorionic twins; MZ MC = monozygotic monochorionic twins; DZO = opposite-sex dizygotic twins; DZF = dizygotic female twins; DZM = dizygotic male twins.
Participant Retention
In addition to the above mentioned procedures to maximize participation between recruitment and delivery, we minimized attrition rates between birth and 18 months (Table 2) by sending twins annual birthday cards, by contacting mothers well in advance, and by sending out annual newsletters. The newsletters contained information on twins and twinning (‘myths about multiples’), lay summaries of results, press cuttings, and each featured a specific twin pair (‘twins’ tales’). Some tracing, including searching the Internet and electoral registries, was also attempted for the families we had lost contact with, with limited success. Since following up twins at 18 months of age, we have contacted all families to inform them of a local twins’ festival, with the added bonus of being able to update contact information. Future work will focus on obtaining email addresses from all participants and establishing a PETS Web/social media page.
Major Accomplishments
Our research to date has focused on the following questions: (1) To what extent does the intrauterine experience render MZ and DZ twins epigenetically discordant? (2) If such differences exist, what are the relative influences of genetic, shared, and non-shared intrauterine environment and which genomic regions are more epigenetically plastic? (3) Do twins drift apart epigenetically during this period and beyond? Below we summarize our research publications to date.
DNA Methylation Analysis of Multiple Tissues From Newborn Twins Reveals Both Genetic and Intrauterine Components to Variation in the Human Neonatal Epigenome
In our first PETS publication (Ollikainen et al., Reference Ollikainen, Smith, Joo, Ng, Andronikos, Novakovic and Craig2010), we focused on measuring the epigenetic mark of DNA methylation within the imprinted insulin-like growth factor 2 (IGF2) locus, important for fetal growth and previously shown to be influenced by shared intrauterine environment (Heijmans et al., Reference Heijmans, Tobi, Stein, Putter, Blauw, Susser and Lumey2008; Steegers-Theunissen et al., Reference Steegers-Theunissen, Obermann-Borst, Kremer, Lindemans, Siebel, Steegers and Heijmans2009; Tobi et al., Reference Tobi, Lumey, Talens, Kremer, Putter, Stein and Heijmans2009). We examined DNA methylation of four differentially methylated regions associated with IGF2 locus in multiple birth tissues (CBMCs, granulocytes, HUVECs, buccal epithelial cells, and placenta) derived from 56 MZ and 35 DZ pairs using the SEQUENOM EpiTYPER platform. In this article, we were the first to demonstrate methylation discordance present at birth within twin pairs. Furthermore, DZ pairs showing greater discordance than MZ pairs. These data highlight the contribution of non-shared intrauterine environment and genetic factors to the establishment and maintenance of foetal epigenome. We also found a wide variation in within-pair discordance between tissues. Our findings support the role of epigenetics in the DOHaD paradigm and suggest that multiple tissues need to be examined in such studies.
Expression Discordance of MZ Twins at Birth: Effect of Intrauterine Environment and a Possible Mechanism for Fetal Programming
As epigenetic marks govern gene expression, we used gene expression discordance as a proxy measure of epigenetic discordance in MZ twins at birth in two cell types: CBMCs and HUVECs (Gordon et al., Reference Gordon, Joo, Andronikos, Ollikainen, Wallace, Umstad and Craig2011). We found strong evidence of expression discordance at birth in both cell types and, assuming that MZ twins start life as epigenetically identical, we conclude that a combination of stochastic and non-shared intrauterine environmental influences shape the epigenome in utero. To support the role of the latter, we found that within-pair differences in expression were most pronounced in genes involved in response to the external environment (Gordon et al., Reference Gordon, Joo, Andronikos, Ollikainen, Wallace, Umstad and Craig2011). Focusing on birth weight, previously identified as a predisposing factor for cardiovascular, metabolic, and other complex diseases, and using a statistical model that estimated association based on within-pair variation of expression and birth weight, we found some association between birth weight and expression of genes involved in metabolism and cardiovascular function. Of interest, expression of LEPROTL1 (leptin receptor overlapping transcript-like 1) was significantly negatively correlated with birth weight in both cell types. This gene and its leptin-regulating homologue Leprot have been associated with growth in mice (Touvier et al., Reference Touvier, Conte-Auriol, Briand, Cudejko, Paumelle, Caron and Bailleu2009). Leptin plays a key role in regulating energy intake and energy expenditure, including appetite and metabolism, and is one of the most important adipose-derived hormones. There is also evidence that leptin resistance may be programmed pre-natally (Struwe et al., Reference Struwe, Berzl, Schild, Blessing, Drexel, Hauck and Dötsch2010). This data provides a plausible biological mechanism for the link between low birth weight, obesity, and other metabolic and related diseases in later life.
Neonatal DNA Methylation Profile in Human Twins is Specified By a Complex Interplay Between Intrauterine Environmental and Genetic Factors, Subject to Tissue-Specific Influence
Following on from our genome-scale study of gene expression, we turned our attention to DNA methylation, using the Illumina Infinium HumanMethylation27 (HM27) BeadChip array platform (Bibikova et al., Reference Bibikova, Le, Barnes, Saedinia-Melnyk, Zhou, Shen and Gunderson2009; Rajendram et al., Reference Rajendram, Ferreira, Grafodatskaya, Choufani, Chiang, Pu and Weksberg2011; Weisenberger et al., Reference Weisenberger, Van Den Berg, Pan, Berman and Laird2008). This platform interrogates >27,000 CpG dinucleotides primarily associated with 14,475 transcription start sites and has a high technical reproducibility. Using methylation of each CpG site as a phenotype, we examined discordance levels in three neonatal tissues (CBMCs, HUVECs, placenta) from 22 MZ and 12 DZ twin pairs (Gordon et al., Reference Gordon, Joo, Powell, Ollikainen, Novakovic, Li and Saffery2012). We found that MZ and DZ twins exhibit a wide range of within-pair differences at birth. We also found that the range of discordance overlaps between MZ twins, DZ twins, and unrelated, but that the median discordance increases across these groups. Variance component decomposition analysis revealed a low mean heritability and negligible levels of mean shared environment across all tissues, although a wide range of values was found throughout the genome. The residual, large (~90%) component of variation of neonatal DNA methylation represents the combined effects of non-shared intrauterine environment, stochastic factors, and measurement error. We suggest that although genetic factors can influence epigenetic marks such as DNA methylation, that intrauterine environment and/or stochastic influences can dominate in some individuals. As with our study of gene expression, methylation discordance was regressed on birth weight discordance and revealed a general association between methylation of genes involved in lipid metabolism and biosynthesis. Taken together, these data adds further support for the involvement of epigenetic mechanisms within the DOHaD paradigm. In collaboration with other groups studying DNA methylation in older twins using the same array platform, we also found no evidence for epigenetic drift between birth and 18 months of age (increasing differences in epigenetic profile as a function of age) and we suggest that to better understand epigenetic changes as twins age, longitudinal studies that take into account specific environments will need to be conducted.
Maternal Vitamin D Predominates Over Genetic Factors in Determining Neonatal Circulating Vitamin D Concentrations
Vitamin D is a hormone with roles in several molecular processes including calcium homeostasis, cell proliferation, cell-cycle control, and immune function (Holick, Reference Holick2007). Humans acquire vitamin D through exposure to sunlight, with other sources including diet and dietary supplements (IOM, 2011). Vitamin D metabolism is regulated by several enzymes, including CYP24A1, which catabolises active vitamin D into less active metabolites (Jones et al., Reference Jones, Prosser and Kaufmann2012). Vitamin D also plays a role in modulating the maternal immune response and the transport of other nutrients across the placenta, including calcium (Shin et al., Reference Shin, Choi, Longtine and Nelson2010). We previously found that the CYP24A1 gene promoter is methylated in the human placenta, but not in other human tissues (Novakovic et al., Reference Novakovic, Sibson, Ng, Manuelpillai, Rakyan, Down and Saffery2009). We proposed that silencing of CYP24A1 might facilitate the transfer of circulating vitamin D (25-OH-D) to the fetal circulation. In this study, the twin model was used to measure the contribution of genetic and environmental factors to CYP24A1 promoter methylation and neonatal circulating vitamin D levels. A total of 86 mother-twin trios (32 MZ and 54 DZ pairs) were used in this analysis. Measurements included maternal serum vitamin D at 28 weeks’ gestation, cord blood 25-OH-D at term, and placental CYP24A1 promoter methylation level, which was quantified using the SEQUENOM EpiTYPER platform. Our results indicate that CYP24A1 promoter methylation is controlled by both genetic and environmental/stochastic influences. However, there was no association between CYP24A1 methylation level and maternal/neonatal circulating vitamin D levels (Novakovic et al., in press). Furthermore, we found that maternal 25-OH-D concentration is the major determinant of neonatal 25-OH-D concentrations, with the underlying genetics not playing a significant role (Novakovic et al., in press).
Current Work
A number of projects are underway and we feature two below.
Association of Maternal Environmental and Nutrient Supply Line Factors With DNA Methylation at the Imprinted IGF2/H19 Locus in Multiple Tissues of Newborn Twins
We investigated the relationship between multiple shared gestational factors and DNA methylation within regions influencing DNA methylation within the IGF2 locus in the same five cell types and using the SEQUENOM EpiTYPER platform as detailed above. Using multiple linear regression, our preliminary findings showed a strong relationship between non-shared environments such as the site of cord insertion, and shared factors such as a mother with gestational diabetes and levels of DNA methylation at this locus in most or all of the regions studied, across most of the all five cell types. However, some environments showed strong associations with DNA methylation within a subset of tissues and genomic regions. These data support our earlier conclusions that non-shared environment is an important influence on neonatal epigenetics and that multiple tissues should be investigated in such studies.
Longitudinal Genome-Scale Analysis of DNA Methylation in Twins From Birth to 18 Months of Age Reveals Rapid Epigenetic Change and Pair-Specific Effects of Epigenetic Drift and Convergence
Following on from our recent genome-scale study of neonatal DNA methylation using the HM27 array, we have spearheaded the first longitudinal, genome-scale study of DNA methylation in young twins. This time, we used the successor to the HM27 platform, the HM450 array (>450,000 CpG probes) to examine DNA methylation changes in DNA from buccal cells from birth and 18 months of age. We found that approximately one-third of CpGs changed over this time at a genome-wide level of significance (adjusted p < .05), indicating that the epigenome is changing rapidly in early postnatal life. Interestingly, we found evidence for epigenetic drift in some pairs, but the opposite, epigenetic convergence in others, most likely reflecting pair-specific influence on DNA methylation dynamics.
Future Directions
To our knowledge, PETS is the first antenatally recruited longitudinal study of twins that has attempted to bridge the gap between environment and genetic/epigenetic factors contributing to phenotypic variability in humans in early life. The unique and labor-intensive aspect of our approach lies in the collection of a diverse range of biospecimens of both maternal and fetal origin. Although originally established to examine the relative contribution of environmental and genetic determinants on the epigenetic profile of humans at birth and their relative contribution to stability of the early life epigenetic profile, it is clear that the utility of the PETS cohort extends beyond this original focus.
Beyond Epigenetics at Birth
The combined strength imparted by the availability of extensive environmental and biological samples at birth provides tremendous opportunities to examine factors that regulate other aspects of cellular function and disease. This includes fundamental questions related to the early life determinants of such things as telomere length or mitochondrial copy number across different tissues at birth and their rate of change in early life.
Emergence of Phenotypic Variability With Time
One of the major challenges associated with linking early life environmental determinants and epigenetic variability with specific health outcomes is the general lack of phenotypic discordance for such outcomes at birth. We have previously dealt with this challenge by focusing on continuous outcome variables of within-pair difference measurable at birth, including birth weight, circulating vitamin D, and plasma folic acid, each of which are implicated in the later manifestation of adverse health outcomes. However, as the PETS twins begin to age, the incidence of relatively common phenotypes such as allergy and atopy (estimated heritability 0.5–0.8; Jackola, Reference Jackola2007; Jackola et al., Reference Jackola, Miller, Liebeler and Blumenthal2007; Liebeler et al., Reference Liebeler, Basu and Jackola2007; Liu et al., Reference Liu, Zhang, Tsai, Hong, Wang, Fang and Wang2009) and early risk factors for cardiovascular disease (estimated heritability 0.4–0.8; Jermendy et al., Reference Jermendy, Horvath, Littvay, Steinbach, Jermendy, Tarnoki and Osztovits2011; Middelberg et al., Reference Middelberg, Martin and Whitfield2007; Osztovits et al., Reference Osztovits, Horvath, Littvay, Steinbach, Jermendy, Tarnoki and Jermendy2011) will begin to emerge in sufficient numbers to allow an exploration of the interrelationship between environmental, genetic, and epigenetic variation (including stability over time), to phenotype outcome. Given previous heritability estimates, we are particularly interested in exploring the relationship between epigenetic and phenotypic variability using a phenotypically discordant MZ twin model, now widely regarded as an extremely valuable approach for the dissection of factors contributing to phenotypic variation associated with complex disorders. Additional sampling at later time points will also allow an examination of the relationship between genes and pre- and post-natal environment to such areas as microbial diversity with or without a further association to phenotype.
A Note on Maternal Observational Data
In the 18 months questionnaire, we asked brief questions on developmental milestones — hearing, speech, diet, hospitalization, health, and vision — to roughly gauge the incidence of these phenotypes to inform future studies. We also asked an open, unstructured question: ‘Is there anything else you would like to tell us about your twins? For example, other differences between your twins?’ With a 93% response rate, we discovered that parents of twins had much to say about this topic. Although this question wasn't structured for accurate qualitative analysis, it is worth noting that the most frequently reported discordance related to temperament (80% of pairs), physical activity (70%), speech and language (40%), size (31%), and eating (25%). Interestingly, 11 pairs (5%) were reported to swap opposing temperaments one or more times. In the structured health questionnaire, 24% of MZ pairs and 22% of DZ pairs were reported by mothers to be discordant for any infection at any time during their first 18 months.
Limitations and Lessons
The establishment of a longitudinal birth cohort of twins with sufficient depth of data and biospecimen collection to show broad utility in epigenetic epidemiology is a formidable challenge. Balancing issues of utility with feasibility inherent in most cohorts are amplified when the aim is to attend all deliveries in order to collect multiple birth biospecimens suitable for future molecular and other biomarker analysis. Not surprisingly, the PETS biorepository is not 100% complete in this regard, with relatively few complete sets of biospecimens, particularly with matched sets of blood fractions collected longitudinally from both twins within a pair. In addition, the collection of cord blood at birth was done after delivery of the final placenta, often resulting in clotting of blood in one of a pair of placentas, thereby limiting sample volume. Other potential modifications to biospecimen collection that would have boosted utility and not greatly impacted on feasibility could have included the addition of longitudinal maternal and childhood stool and/or rectal or oral swabs for microbiome measurements, collection of hair and/or urine and/or fingernails for measuring environmental exposures (e.g., cotinine, pesticides). Ideally, longitudinal blood and buccal cell samples would have been collected from infants at multiple time points between the current birth and 18 month samples. Balancing future utility with burdens on twins’ families and budget is a difficult task but one that we aim to improve.
Acknowledgments
We thank all of the families who participated in the PETS study; our tireless research assistants, Sarah Healy, Tina Vaiano, Nicole Brooks, Jennifer Foord, Sheila Holland, Ann Krastev, and Joanne Mockler; Anna Czajko, Mandy Parkinson-Bates, Hong Kiat Ng, Nur Khairunnisa Abdul Aziz, and Noha Ahmed Nasef for technical assistance; bioinformaticians Gordon Smyth, Alicia Oshlack, Katrina Bell, and Katherine Smith; phlebotomists Bernie McCudden, Judith Spottswood, Erin Hill, and staff from Royal Childrens Hospital Pathology Collection; and Geraldine McIlroy and Hien Nguyen for their administrative assistance. This work was supported by grants from the Australian National Health and Medical Research Council (grant numbers 437015 and 607358 to JC, and RS), the Bonnie Babes Foundation (grant number BBF20704 to EJ), the Sigrid Juselius Foundation (to MO), the Academy of Finland (to MO), the Finnish Cultural Foundation (to MO), the Financial Markets Foundation for Children (grant no. 032-2007), and by the Victorian Government's Operational Infrastructure Support Program.


