



Introduction
The devastating environmental consequences of the end-Cretaceous bolide impact affected both marine and terrestrial organisms (e.g., Alvarez et al. Reference Alvarez, Alvarez, Asaro and Michel1980; Prinn and Fegley Reference Prinn and Fegley1987; Robertson et al. Reference Robertson, Lewis, Sheehan and Toon2013; Vellekoop et al. Reference Vellekoop, Sluijs, Smit, Schouten, Weijers, Sinninghe Damste and Brinkhuis2014; Tyrrell et al. Reference Tyrrell, Merico and Armstrong McKay2015; Artemieva and Morgan Reference Artemieva and Morgan2017; Brugger et al. Reference Brugger, Feulner and Petri2017). Over 60% of Cretaceous species became extinct, making the Cretaceous–Paleogene (K/Pg) event the most recent of the “big five” mass extinctions (Raup and Sepkoski Reference Raup and Sepkoski1982; Jablonski Reference Jablonski2005; Schulte et al. Reference Schulte, Alegret, Arenillas, Arz, Barton, Bown, Bralower, Christeson, Claeys, Cockell, Collins, Deutsch, Goldin, Goto, Grajales-Nishimura, Grieve, Gulick, Johnson, Kiessling, Koeberl, Kring, MacLeod, Matsui, Melosh, Montanari, Morgan, Neal, Nichols, Norris, Pierazzo, Ravizza, Rebolledo-Vieyra, Reimold, Robin, Salge, Speijer, Sweet, Urrutia-Fucugauchi, Vajda, Whalen and Willumsen2010). Although the K/Pg event affected biotas globally, the severity of the extinction and the pacing of the recovery were geographically heterogeneous for some groups of organisms (see next paragraph). Modern biodiversity is shaped by the surviving lineages of the K/Pg (e.g., Erwin Reference Erwin2002; Krug et al. Reference Krug, Jablonski and Valentine2017), establishing today's biogeographic patterns, in part, as the legacy of a globally heterogeneous extinction (e.g., Wolfe Reference Wolfe1987).
Southern Hemisphere records of calcareous nannoplankton, insect herbivory, and terrestrial palynomorphs indicate lower extinction and/or faster recoveries than Northern Hemisphere counterparts (Vajda et al. Reference Vajda, Raine and Hollis2001; Vajda and Raine Reference Vajda and Raine2003; Iglesias et al. Reference Iglesias, Wilf, Johnson, Cúneo and Matheos2007; Pole and Vajda Reference Pole and Vajda2009; Jiang et al. Reference Jiang, Bralower, Patzkowsky, Kump and Schueth2010; Barreda et al. Reference Barreda, Cúneo, Wilf, Currano, Scasso and Brinkhuis2012; Cantrill and Poole Reference Cantrill, Poole, Cantrill and Poole2012; Schueth et al. Reference Schueth, Bralower, Jiang and Patzkowsky2015; Donovan et al. Reference Donovan, Iglesias, Wilf, Labandeira and Cúneo2016). Furthermore, several plant and vertebrate groups that had been known only from Mesozoic localities in the Northern Hemisphere, have been reported in Cenozoic Southern Hemisphere deposits suggesting that they survived the K/Pg in southern latitudes (Case and Woodburne Reference Case and Woodburne1986; Pascual et al. Reference Pascual, Archer, Jaureguizar, Prado, Godthelp and Hand1992; Bonaparte et al. Reference Bonaparte, Van Valen and Kramartz1993; Goin et al. Reference Goin, Pascual, Tejedor, Gelfo, Woodburne, Case, Reguero, Bond, López, Cione, Sauthier, Balarino, Scasso, Medina and Ubaldón2006, Reference Goin, Tejedor, Chornogubsky, López, Gelfo and Woodburne2012; McLoughlin et al. Reference McLoughlin, Carpenter, Jordan and Hill2008, Reference McLoughlin, Carpenter and Pott2011; Gelfo et al. Reference Gelfo, Goin, Woodburne and Muizon2009; Sterli and de la Fuente Reference Sterli and de la Fuente2019). Proposed explanations for this geographic heterogeneity (Jiang et al. Reference Jiang, Bralower, Patzkowsky, Kump and Schueth2010; Donovan et al. Reference Donovan, Iglesias, Wilf, Labandeira and Cúneo2016, Reference Donovan, Iglesias, Wilf, Labandeira and Cúneo2018) have referenced increased distance from the Mexican impact site (Schulte et al. Reference Schulte, Alegret, Arenillas, Arz, Barton, Bown, Bralower, Christeson, Claeys, Cockell, Collins, Deutsch, Goldin, Goto, Grajales-Nishimura, Grieve, Gulick, Johnson, Kiessling, Koeberl, Kring, MacLeod, Matsui, Melosh, Montanari, Morgan, Neal, Nichols, Norris, Pierazzo, Ravizza, Rebolledo-Vieyra, Reimold, Robin, Salge, Speijer, Sweet, Urrutia-Fucugauchi, Vajda, Whalen and Willumsen2010), oceanic buffering of impact winter temperatures in the Southern Hemisphere (Bardeen et al. Reference Bardeen, Garcia, Toon and Conley2017; Tabor et al. Reference Tabor, Bardeen, Otto-Bliesner, Garcia and Toon2020), and a bolide impact angle that would have directed most ejecta and debris northward (Schultz and D'Hondt Reference Schultz and D'Hondt1996).
Because plants are the primary producers in terrestrial ecosystems and one of the most biodiverse groups of organisms, the paleobotanical record is critical for understanding extinction events on land (e.g., Nichols and Johnson Reference Nichols and Johnson2008). However, the vast majority of plant-bearing K/Pg continental sites is concentrated in the western interior of North America (NAM), which bears numerous examples of stratigraphically and temporally well constrained boundary-spanning palynofloras (e.g., Nichols et al. Reference Nichols, Jarzen, Orth and Oliver1986; Nichols and Fleming Reference Nichols and Fleming1990; Sweet et al. Reference Sweet, Braman and Lerbekmo1990, Reference Sweet, Braman and Lerbekmo1999; Sweet and Braman Reference Sweet and Braman2001; Nichols Reference Nichols2002; Bercovici et al. Reference Bercovici, Pearson, Nichols and Wood2009) and macrofloras (e.g., Wolfe and Upchurch Reference Wolfe and Upchurch1986; Johnson et al. Reference Johnson, Nichols, Attrep and Orth1989; Johnson and Hickey Reference Johnson and Hickey1990; Johnson Reference Johnson1992, Reference Johnson2002; Upchurch Reference Upchurch1995; Barclay et al. Reference Barclay, Johnson, Betterton and Dilcher2003; Nichols and Johnson Reference Nichols and Johnson2008). Southern Hemisphere K/Pg-spanning paleofloras are scarce and have been primarily limited to palynological records (e.g., Vajda-Santivanez Reference Vajda-Santivanez1999; Vajda et al. Reference Vajda, Raine and Hollis2001; Vajda and Raine Reference Vajda and Raine2003; Pole and Vajda Reference Pole and Vajda2009; Barreda et al. Reference Barreda, Cúneo, Wilf, Currano, Scasso and Brinkhuis2012; Cantrill and Poole Reference Cantrill, Poole, Cantrill and Poole2012; Scasso et al. Reference Scasso, Prámparo, Vellekoop, Franzosi, Castro and Damsté2020). Cretaceous–Paleogene macrofloras from New Zealand revealed a dramatic floral turnover (Pole and Vajda Reference Pole and Vajda2009) and a paleoclimatic cooling trend consistent with global records (Kennedy et al. Reference Kennedy, Spicer and Rees2002) across the K/Pg, but the severity of the macrofloral extinction was not estimated. Until now, no well-constrained and well-sampled macrofloral Maastrichtian and Danian sites from the same region have been available in the Southern Hemisphere.
Recent studies of the latest Cretaceous (Maastrichtian) Lefipán and early Paleocene (Danian) Salamanca and Peñas Coloradas Formations of Chubut, Argentine Patagonia (Fig. 1; and see “Materials” and “Analytical Methods”) showed lower palynological extinction, faster recovery of insect herbivory damage-type diversity, and remarkably diverse Danian macrofloras compared with most NAM sections of the same ages. Specifically, spore and pollen records showed <10% extinction across the K/Pg compared with the 30%–40% extinction in NAM palynofloras (Nichols and Fleming Reference Nichols and Fleming1990; Sweet and Braman Reference Sweet and Braman2001; Hotton Reference Hotton2002; Nichols Reference Nichols2002; Barreda et al. Reference Barreda, Cúneo, Wilf, Currano, Scasso and Brinkhuis2012). Notably, in early Danian Patagonian pollen records, an abundance spike of the conifer Classopollis represents the last record of a genus that is otherwise only known until the Late Cretaceous worldwide (Barreda et al. Reference Barreda, Cúneo, Wilf, Currano, Scasso and Brinkhuis2012). Insect feeding-damage types on angiosperm leaves recovered to pre-K/Pg diversity levels within about 4 Myr, compared with the estimated ca. 9 Myr for NAM (Donovan et al. Reference Donovan, Iglesias, Wilf, Labandeira and Cúneo2016, Reference Donovan, Iglesias, Wilf, Labandeira and Cúneo2018), and Danian macrofloras of the Salamanca Formation are much more diverse than most coeval NAM counterparts, suggesting a faster-paced Patagonian recovery (Iglesias et al. Reference Iglesias, Wilf, Johnson, Cúneo and Matheos2007). However, comparable Maastrichtian leaf floras from the same Patagonian region have not yet been evaluated using similar methods.

Figure 1. Setting of paleobotanical localities. A, Paleogeographic reconstruction of Patagonia at K/Pg time, redrawn from Scasso et al. (Reference Scasso, Aberhan, Ruiz, Weidemeyer, Medina and Kiessling2012). Light blue diagonal pattern: shallow platform; yellow horizontal pattern: coastal lowlands; dashed line: approximate boundary between paleoclimatic belts as inferred by Scasso et al. (Reference Scasso, Aberhan, Ruiz, Weidemeyer, Medina and Kiessling2012). Locations of panels B and C marked. B, Location of Scasso et al.'s (2012) San Ramón section (within star) of the Lefipán Fm., containing Maastrichtian macrofloras and Maastrichtian and Danian palynofloras. Modified after Barreda et al. (Reference Barreda, Cúneo, Wilf, Currano, Scasso and Brinkhuis2012). C, Locations of Danian paleobotanical sites: Palacio de los Loros (PL), Salamanca Fm.; Las Flores (LF), Peñas Coloradas Fm.; and Bosque Petrificado José Ormachea (OR, Salamanca Fm., used here for pollen data only). Modified after Clyde et al. (Reference Clyde, Wilf, Iglesias, Slingerland, Barnum, Bijl, Bralower, Brinkhuis, Comer, Huber, Ibañez-Mejia, Jicha, Krause, Schueth, Singer, Raigemborn, Schmitz, Sluijs and Zamaloa2014) and Comer et al. (Reference Comer, Slingerland, Krause, Iglesias, Clyde, Raigemborn and Wilf2015). D, Summary stratigraphy showing chronostratigraphic and absolute age constraints, placement of palynological (tick marks) and macrofloral sampling sites (black stars; compiled from Iglesias Reference Iglesias2007; Barreda et al. Reference Barreda, Cúneo, Wilf, Currano, Scasso and Brinkhuis2012; Clyde et al. Reference Clyde, Wilf, Iglesias, Slingerland, Barnum, Bijl, Bralower, Brinkhuis, Comer, Huber, Ibañez-Mejia, Jicha, Krause, Schueth, Singer, Raigemborn, Schmitz, Sluijs and Zamaloa2014; Comer et al. Reference Comer, Slingerland, Krause, Iglesias, Clyde, Raigemborn and Wilf2015; Donovan et al. Reference Donovan, Iglesias, Wilf, Labandeira and Cúneo2016, Reference Donovan, Iglesias, Wilf, Labandeira and Cúneo2018). Summary of absolute ages of the fossil sites (see text for details): Lefipán floras, 67–66 Ma, most likely 66.5–66.0 Ma; PL1 flora, 66.4–65.7 Ma (C29n); PL2 flora, 64.7–63.5 Ma (C28n); LF flora, 62.5–62.2 Ma (C27n). Range of stratigraphic section comprising approximate locations of LefW sampling horizons (as described in Vellekoop et al. Reference Vellekoop, Holwerda, Prámparo, Willmott, Schouten, Cúneo, Scasso and Brinkhuis2017) indicated by dashed line (see “Materials” for details). LefE, LefL, and LefW are shown on a single column for simplicity but represent different locations (see text for details). LefE and LefL come from a single laterally extensive horizon that varies in stratigraphic distance below the Turritella bed. The position shown is for LefL. BBF, Bajo Barreal Fm. (Color online.)
Gymnosperm macrofossils of the Lefipán and Salamanca Formations have been the subject of taxonomic studies (Zamuner et al. Reference Zamuner, Brea, Ganuza and Matheos2000; Brea et al. Reference Brea, Matheos, Zamuner and Ganuza2005; Quiroga et al. Reference Quiroga, Mathiasen, Iglesias, Mill and Premoli2015; Ruiz et al. Reference Ruiz, Brea, Raigemborn and Matheos2017; Wilf et al. Reference Wilf, Donovan, Cúneo and Gandolfo2017; Andruchow-Colombo et al. Reference Andruchow-Colombo, Escapa, Cúneo and Gandolfo2018, Reference Andruchow-Colombo, Escapa, Carpenter, Hill, Iglesias, Abarzua and Wilf2019; Escapa et al. Reference Escapa, Iglesias, Wilf, Catalano, Caraballo-Ortiz and Cúneo2018), indicating the survival of the Podocarpaceae and Araucariaceae conifer families. The presence of several derived angiosperm groups known from reproductive material in the Salamanca Formation suggests that they also represent K/Pg survivor families (Iglesias et al. Reference Iglesias, Wilf, Johnson, Cúneo and Matheos2007; Raigemborn et al. Reference Raigemborn, Brea, Zucol and Matheos2009; Jud et al. Reference Jud, Gandolfo, Iglesias and Wilf2017, Reference Jud, Gandolfo, Iglesias and Wilf2018a,Reference Jud, Iglesias, Wilf and Gandolfob; Supplementary Tables 1, 2). Angiosperm leaf fossils have sourced the aforementioned work on K/Pg insect damage (Donovan et al. Reference Donovan, Iglesias, Wilf, Labandeira and Cúneo2016, Reference Donovan, Iglesias, Wilf, Labandeira and Cúneo2018), yet the extensive leaf collections on which the insect herbivory was documented have not been rigorously compared to quantify macrofloral species extinction in Patagonia across the Cretaceous/Paleogene transition.
Fossil dicot leaves from the Maastrichtian and Danian of Patagonia (Fig. 1) together offer insight into macrofloral K/Pg diversity, and potential extinction and recovery patterns with well-sampled, stratigraphically and temporally constrained collections for the first time in the Southern Hemisphere (Iglesias et al. Reference Iglesias, Wilf, Johnson, Cúneo and Matheos2007; Scasso et al. Reference Scasso, Aberhan, Ruiz, Weidemeyer, Medina and Kiessling2012; Clyde et al. Reference Clyde, Wilf, Iglesias, Slingerland, Barnum, Bijl, Bralower, Brinkhuis, Comer, Huber, Ibañez-Mejia, Jicha, Krause, Schueth, Singer, Raigemborn, Schmitz, Sluijs and Zamaloa2014; Donovan et al. Reference Donovan, Iglesias, Wilf, Labandeira and Cúneo2016, Reference Donovan, Iglesias, Wilf, Labandeira and Cúneo2018). Through the analysis of more than 3500 leaf fossils from four localities, this study addresses terminal Cretaceous and early Paleocene macrofloral diversity in Patagonia, and the possible effects of the K/Pg extinction event on the floras of the region while considering the potential climatic, sampling, and ecological biases impacting the observations. Although the main goal of this contribution is to document the first temporally constrained K/Pg-spanning macrofloral assemblages occurring within the same region in the Southern Hemisphere, we compare our results with the well-sampled, boundary-spanning K/Pg NAM macrofloras from similar absolute paleolatitudes (ca. 50°N or 50°S) in the Williston Basin (WB) in North Dakota (Johnson Reference Johnson2002; Wilf and Johnson Reference Wilf and Johnson2004) to further explore potential biases in our observations and to examine potential geographic heterogeneity in macrofloral patterns among two widely separated regions.
Materials
The collections studied here are primarily the same as those from Chubut, Argentina, used in earlier work on Danian floral diversity (e.g., Iglesias Reference Iglesias2007; Iglesias et al. Reference Iglesias, Wilf, Johnson, Cúneo and Matheos2007) and K/Pg insect damage (Donovan et al. Reference Donovan, Iglesias, Wilf, Labandeira and Cúneo2016, Reference Donovan, Iglesias, Wilf, Labandeira and Cúneo2018), plus some additional material previously not cataloged (Supplementary Table 3). Iglesias et al. (Reference Iglesias, Wilf, Johnson, Cúneo and Matheos2007) documented Danian dicot-leaf diversity by morphotyping and assigning systematic affinities when possible (Iglesias Reference Iglesias2007; Iglesias et al. Reference Iglesias, Wilf, Johnson, Cúneo and Matheos2007; more recent updates shown in Supplementary Table 1). Donovan et al. (Reference Donovan, Iglesias, Wilf, Labandeira and Cúneo2016, Reference Donovan, Iglesias, Wilf, Labandeira and Cúneo2018) analyzed the insect feeding-damage diversity on the leaves from Maastrichtian and Danian collections and proposed a preliminary morphotype classification for the Maastrichtian leaves. The leaf collections from the Lefipán (Maastrichtian) and Salamanca and Peñas Coloradas (Danian) Formations were compiled over a series of field trips involving the four junior authors and others since 2003, and they are curated in the Paleobotanical Collection of the Museo Paleontológico Egidio Feruglio (MEF; repository acronym MPEF-Pb), Trelew, Argentina (Supplementary Table 3). All collections studied are unbiased, complete census collections of all identifiable material found, taken to the lab at MEF, and vetted and tallied. Although they are separated by ca. 400 km (Fig. 1), and the K/Pg boundary horizon itself is not preserved in the Lefipán or Salamanca Formations, these Maastrichtian and Danian collections represent broadly similar, marginal marine depositional settings and the only large, stratigraphically constrained leaf collections from their time periods sourced from a single region in the Southern Hemisphere. In addition to the summary presentation and citations that follow, further detailed accounts of collecting methods and sites and full descriptions of the leaf morphotypes are in separate preparation.
Lefipán Formation
The siliciclastic Lefipán Formation conformably overlies the Campanian–Maastrichtian fluvio-estuarine Paso del Sapo Formation, and it is unconformably overlain by the Eocene Barda Colorada Ignimbrite (Spalletti Reference Spalletti1996; Aragón and Mazzoni Reference Aragón and Mazzoni1997; Scasso et al. Reference Scasso, Aberhan, Ruiz, Weidemeyer, Medina and Kiessling2012; Aragón et al. Reference Aragón, Castro, Diaz-Alvarado, Pinotti, D'Eramo, Demartis, Coniglio, Hernando and Rodriguez2018) and other Paleogene volcanic units (Aragón et al. Reference Aragón, Castro, Diaz-Alvarado, Pinotti, D'Eramo, Demartis, Coniglio, Hernando and Rodriguez2018). Deposited in the shallow, paleo-Atlantic Paso del Sapo embayment (Fig. 1A,B), the Lefipán Formation records the latest Cretaceous (?Campanian–Maastrichtian) to early Paleocene (Danian) infilling of the Jurassic–early Paleocene Cañadón Asfalto Basin (Fig. 1A) (Spalletti Reference Spalletti1996; Scasso et al. Reference Scasso, Aberhan, Ruiz, Weidemeyer, Medina and Kiessling2012; Figari et al. Reference Figari, Scasso, Cúneo and Escapa2015). Leaf cuticular pCO2 estimates (Martínez et al. Reference Martínez, Gandolfo and Cúneo2018), TEX86 proxy records (Scasso et al. Reference Scasso, Aberhan, Ruiz, Weidemeyer, Medina and Kiessling2012; Vellekoop et al. Reference Vellekoop, Holwerda, Prámparo, Willmott, Schouten, Cúneo, Scasso and Brinkhuis2017), and paleobotanical evidence (Baldoni Reference Baldoni1992; Baldoni et al. Reference Baldoni, Askin and Baldoni1993; Barreda et al. Reference Barreda, Cúneo, Wilf, Currano, Scasso and Brinkhuis2012; Wilf et al. Reference Wilf, Donovan, Cúneo and Gandolfo2017) indicate that the upper Maastrichtian Lefipán Formation was deposited in mesothermal (or warmer), frost-free climates (Fig. 1A) along a coastline fringed by patchy woodlands and mangroves; macrofossil evidence suggests areas of rain forest as well (Wilf et al. Reference Wilf, Donovan, Cúneo and Gandolfo2017; Escapa et al. Reference Escapa, Iglesias, Wilf, Catalano, Caraballo-Ortiz and Cúneo2018).
The San Ramón Section (SRS; Fig. 1B,D) studied here is the most complete of the Lefipán Formation (Scasso et al. Reference Scasso, Aberhan, Ruiz, Weidemeyer, Medina and Kiessling2012), comprising marginal, tidal-flat deposits to fully marine sediments within a tidally dominated deltaic setting that ranges from ?Campanian–Maastrichtian to Danian in age (Legarreta and Uliana Reference Legarreta and Uliana1994; Spalletti Reference Spalletti1996; Barreda et al. Reference Barreda, Cúneo, Wilf, Currano, Scasso and Brinkhuis2012; Scasso et al. Reference Scasso, Aberhan, Ruiz, Weidemeyer, Medina and Kiessling2012; Vellekoop et al. Reference Vellekoop, Holwerda, Prámparo, Willmott, Schouten, Cúneo, Scasso and Brinkhuis2017; Butler et al. Reference Butler, Horton, Echaurren, Folguera and Fuentes2019). The K/Pg impact layer recognized at other sites around the world (e.g., Alvarez et al. Reference Alvarez, Alvarez, Asaro and Michel1980, Reference Alvarez, Asaro and Montanari1990; Orth et al. Reference Orth, Gilmore, Knight, Pillmore, Tschudy and Fassett1981; Brooks et al. Reference Brooks, Strong, Lee, Orth, Gilmore, Ryan and Holzbecher1986; Lerbekmo and St. Louis Reference Lerbekmo and St. Louis1986; Schulte et al. Reference Schulte, Alegret, Arenillas, Arz, Barton, Bown, Bralower, Christeson, Claeys, Cockell, Collins, Deutsch, Goldin, Goto, Grajales-Nishimura, Grieve, Gulick, Johnson, Kiessling, Koeberl, Kring, MacLeod, Matsui, Melosh, Montanari, Morgan, Neal, Nichols, Norris, Pierazzo, Ravizza, Rebolledo-Vieyra, Reimold, Robin, Salge, Speijer, Sweet, Urrutia-Fucugauchi, Vajda, Whalen and Willumsen2010) is not preserved in the SRS, probably due to bioturbation or erosion at that particular level (Scasso et al. Reference Scasso, Aberhan, Ruiz, Weidemeyer, Medina and Kiessling2012). However, the K/Pg in the Lefipán Formation is constrained to about 4 m of section based on biostratigraphic data, including the marker Turritella malaspina lag bed as the first Danian deposit (Fig. 1D) (Medina and Camacho Reference Medina and Camacho1990; Scasso et al. Reference Scasso, Aberhan, Ruiz, Weidemeyer, Medina and Kiessling2012), dinoflagellate index taxa (Barreda et al. Reference Barreda, Cúneo, Wilf, Currano, Scasso and Brinkhuis2012; Vellekoop et al. Reference Vellekoop, Holwerda, Prámparo, Willmott, Schouten, Cúneo, Scasso and Brinkhuis2017), age-diagnostic continental palynomorphs (Barreda et al. Reference Barreda, Cúneo, Wilf, Currano, Scasso and Brinkhuis2012), and a sudden and significant turnover of invertebrate faunas (Aberhan and Kiessling Reference Aberhan and Kiessling2014) and microfloras with the extinction of ca. 10% of total and 50% of angiosperm palynotaxa (Fig. 1D) (Barreda et al. Reference Barreda, Cúneo, Wilf, Currano, Scasso and Brinkhuis2012).
Lefipán Formation specimens analyzed here were collected in the SRS (Fig. 1A,C,D) in sandstone–mudstone tidal-flat deposits (Scasso et al. Reference Scasso, Aberhan, Ruiz, Weidemeyer, Medina and Kiessling2012). Localities LefW, LefE, and LefL, as detailed in Donovan et al. (Reference Donovan, Iglesias, Wilf, Labandeira and Cúneo2016, Reference Donovan, Iglesias, Wilf, Labandeira and Cúneo2018) and Wilf et al. (Reference Wilf, Donovan, Cúneo and Gandolfo2017), yielded a total of 1062 dicot-leaf specimens from unbiased collections biostratigraphically constrained to the 67–66 Ma range and most likely in the 66.5–66.0 range based on dinoflagellate markers (Fig. 1D) (Barreda et al. Reference Barreda, Cúneo, Wilf, Currano, Scasso and Brinkhuis2012; Scasso et al. Reference Scasso, Aberhan, Ruiz, Weidemeyer, Medina and Kiessling2012; Vellekoop et al. Reference Vellekoop, Holwerda, Prámparo, Willmott, Schouten, Cúneo, Scasso and Brinkhuis2017). All identifiable material was collected at these sites and taken back to the MEF, where it was vetted and censused (no identifiable fossils were discarded in the field). The LefW macrofloral site corresponds to the same plant-bearing beds discussed in Scasso et al. (Reference Scasso, Aberhan, Ruiz, Weidemeyer, Medina and Kiessling2012) and Vellekoop et al. (Reference Vellekoop, Holwerda, Prámparo, Willmott, Schouten, Cúneo, Scasso and Brinkhuis2017). The LefW collection (Supplementary Table 3) includes 278 dicot-leaf specimens spanning about 20 m of stratigraphic section from four collecting horizons, the youngest of which is located 5 m below the Danian Turritella marker bed (Scasso et al. Reference Scasso, Aberhan, Ruiz, Weidemeyer, Medina and Kiessling2012; Donovan et al. Reference Donovan, Iglesias, Wilf, Labandeira and Cúneo2016; Vellekoop et al. Reference Vellekoop, Holwerda, Prámparo, Willmott, Schouten, Cúneo, Scasso and Brinkhuis2017). Approximately 1000 m map distance east of LefW is the single fossiliferous horizon yielding the macrofloras of the LefE and LefL collections. Locality LefE, 21.5 m below the Turritella marker bed, is closest to LefW. The LefE quarry extends about 40 m along the horizon (Wilf et al. Reference Wilf, Donovan, Cúneo and Gandolfo2017), from where 614 dicot-leaf fossils were collected. LefL is located approximately 500 m (map distance) east of LefE, in the same bed, at 24 m locally below the Turritella bed (Donovan et al. Reference Donovan, Iglesias, Wilf, Labandeira and Cúneo2016) and yielded 170 dicot-leaf specimens. The single fossiliferous horizon containing LefE and LefL cannot be correlated precisely to those in LefW due to cover and erosion in the intervening landscape, but their ages are probably very similar based on stratigraphic position.
Salamanca and Peñas Coloradas Formations
The Danian Salamanca and Peñas Coloradas Formations are the oldest Cenozoic sedimentary units in the north-central San Jorge Basin (Fig. 1A) (Sylwan Reference Sylwan2001; Clyde et al. Reference Clyde, Wilf, Iglesias, Slingerland, Barnum, Bijl, Bralower, Brinkhuis, Comer, Huber, Ibañez-Mejia, Jicha, Krause, Schueth, Singer, Raigemborn, Schmitz, Sluijs and Zamaloa2014; Comer et al. Reference Comer, Slingerland, Krause, Iglesias, Clyde, Raigemborn and Wilf2015). In our study area (Fig. 1A,C), tidally influenced, marginal to estuarine deposits of the early Danian Salamanca Formation (Legarreta and Uliana Reference Legarreta and Uliana1994; Iglesias Reference Iglesias2007; Comer et al. Reference Comer, Slingerland, Krause, Iglesias, Clyde, Raigemborn and Wilf2015) unconformably overlie fluvial and pyroclastic deposits of the Late Cretaceous (Campanian) Bajo Barreal Formation or (where preserved) the Maastrichtian La Angostura Basalt (Fig. 1D) (Iglesias Reference Iglesias2007; Clyde et al. Reference Clyde, Wilf, Iglesias, Slingerland, Barnum, Bijl, Bralower, Brinkhuis, Comer, Huber, Ibañez-Mejia, Jicha, Krause, Schueth, Singer, Raigemborn, Schmitz, Sluijs and Zamaloa2014; Comer et al. Reference Comer, Slingerland, Krause, Iglesias, Clyde, Raigemborn and Wilf2015), and they are unconformably overlain by the fluvio-volcanic deposits of the late Danian Peñas Coloradas Formation of the continental Rio Chico Group (Fig. 1C,D) (Iglesias Reference Iglesias2007; Raigenborn et al. Reference Raigemborn, Krause, Bellosi and Matheos2010; Clyde et al. Reference Clyde, Wilf, Iglesias, Slingerland, Barnum, Bijl, Bralower, Brinkhuis, Comer, Huber, Ibañez-Mejia, Jicha, Krause, Schueth, Singer, Raigemborn, Schmitz, Sluijs and Zamaloa2014; Comer et al. Reference Comer, Slingerland, Krause, Iglesias, Clyde, Raigemborn and Wilf2015; Krause et al. Reference Krause, Clyde, Ibañez-Mejia, Schmitz, Barnum, Bellosi and Wilf2017). At the Las Flores (LF; Fig 1C) Peñas Coloradas Formation locality, deposits overlie the Maastrichtian Colhué Huapi Formation and La Angostura Basalt (Casal et al. Reference Casal, Allard and Foix2015; Comer et al. Reference Comer, Slingerland, Krause, Iglesias, Clyde, Raigemborn and Wilf2015).
Fossil occurrences from the Salamanca and overlying Peñas Coloradas Formations are consistent with humid, mesothermal, frost-free climates (Bonaparte et al. Reference Bonaparte, Van Valen and Kramartz1993; Brea et al. Reference Brea, Matheos, Zamuner and Ganuza2005; Iglesias et al. Reference Iglesias, Wilf, Johnson, Cúneo and Matheos2007; Palazzesi and Barreda Reference Palazzesi and Barreda2007; Raigemborn et al. Reference Raigemborn, Brea, Zucol and Matheos2009; Futey et al. Reference Futey, Gandolfo, Zamaloa, Cúneo and Cladera2012; Clyde et al. Reference Clyde, Wilf, Iglesias, Slingerland, Barnum, Bijl, Bralower, Brinkhuis, Comer, Huber, Ibañez-Mejia, Jicha, Krause, Schueth, Singer, Raigemborn, Schmitz, Sluijs and Zamaloa2014; Ruiz et al. Reference Ruiz, Brea, Raigemborn and Matheos2017, Reference Ruiz, Raigemborn, Brea and Pujana2020). Palynofloras of the Salamanca Formation and related strata over larger areas reflect a range of environments, from mangrove swamps to lowland and upland forests (Petriella and Archangelsky Reference Petriella and Archangelsky1975; Zamaloa and Andreis Reference Zamaloa and Andreis1995; Volkheimer et al. Reference Volkheimer, Scafati and Melendi2007). In the study area (Fig. 1C,D), local pollen records show diverse, angiosperm-dominated lowland forests (Petriella and Archangelsky Reference Petriella and Archangelsky1975; Clyde et al. Reference Clyde, Wilf, Iglesias, Slingerland, Barnum, Bijl, Bralower, Brinkhuis, Comer, Huber, Ibañez-Mejia, Jicha, Krause, Schueth, Singer, Raigemborn, Schmitz, Sluijs and Zamaloa2014).
The unbiased, lab-vetted (the same collecting and tallying methods used as for the Lefipán floras) early Paleocene leaf collections analyzed in this study came from two early Danian Salamanca Formation localities in the Palacio de los Loros sampling area (PL1 and PL2) and one late Danian locality in the Peñas Coloradas Formation, the LF plant-fossil site (Fig. 1B) (Iglesias Reference Iglesias2007; Iglesias et al. Reference Iglesias, Wilf, Johnson, Cúneo and Matheos2007; Clyde et al. Reference Clyde, Wilf, Iglesias, Slingerland, Barnum, Bijl, Bralower, Brinkhuis, Comer, Huber, Ibañez-Mejia, Jicha, Krause, Schueth, Singer, Raigemborn, Schmitz, Sluijs and Zamaloa2014). Biostratigraphic data from foraminifera, dinoflagellates, and calcareous nannoplankton indicate the early Danian age for the local Salamanca Formation, with a maximum absolute age of 67.31 ± 0.55 Ma from 40Ar/39Ar dating of the La Angostura Basalt flow underlying the formation in the easternmost exposures of the study area (Fig. 1D) (Clyde et al. Reference Clyde, Wilf, Iglesias, Slingerland, Barnum, Bijl, Bralower, Brinkhuis, Comer, Huber, Ibañez-Mejia, Jicha, Krause, Schueth, Singer, Raigemborn, Schmitz, Sluijs and Zamaloa2014). The U-Pb age of a tuff layer in the Peñas Coloradas Formation at Palacio de los Loros is 61.984 ± 0.041 Ma, placing it in the late Danian (Fig. 1D) (Clyde et al. Reference Clyde, Wilf, Iglesias, Slingerland, Barnum, Bijl, Bralower, Brinkhuis, Comer, Huber, Ibañez-Mejia, Jicha, Krause, Schueth, Singer, Raigemborn, Schmitz, Sluijs and Zamaloa2014). Combined analysis of paleomagnetic stratigraphy, biostratigraphy, and U-Pb and 40Ar-39Ar ages place the paleobotanical sites PL1, PL2, and LF within geomagnetic polarity chrons C29n (65.58–64.88 Ma), C28n (64.67–63.49 Ma), and C27n (62.52–62.22 Ma), respectively (Fig. 1D) (Clyde et al. Reference Clyde, Wilf, Iglesias, Slingerland, Barnum, Bijl, Bralower, Brinkhuis, Comer, Huber, Ibañez-Mejia, Jicha, Krause, Schueth, Singer, Raigemborn, Schmitz, Sluijs and Zamaloa2014; Comer et al. Reference Comer, Slingerland, Krause, Iglesias, Clyde, Raigemborn and Wilf2015), using the 2012 Geologic Time Scale (Gradstein et al. Reference Gradstein, Ogg, Schmitz and Ogg2012).
Between the early Danian Salamanca and late Danian Peñas Coloradas Formations, there is a sharp change in the depositional environment. The strata at PL1 and PL2 were deposited in marginal estuarine settings, whereas LF was deposited in a nearly fully terrestrial fluvio-volcanic environment (Comer et al. Reference Comer, Slingerland, Krause, Iglesias, Clyde, Raigemborn and Wilf2015). Locality PL1 (Fig. 1A,C,D) produced 1089 dicot-leaf specimens, preserved in sand–siltstone sediments interpreted as lateral accretion beds of abandoned tidal channel fill (facies Sab in Comer et al. Reference Comer, Slingerland, Krause, Iglesias, Clyde, Raigemborn and Wilf2015). The younger locality PL2 yielded 1132 dicot-leaf specimens, collected from silty claystones interpreted as the transitional facies of tidal flats prograding over a coastal plain (facies SCt in Comer et al. Reference Comer, Slingerland, Krause, Iglesias, Clyde, Raigemborn and Wilf2015). Sediments in locality LF are the coarsest-grained of the Danian macrofloral sites, from where 564 dicot-leaf specimens were recovered from poorly sorted litharenites interpreted as channel fills (facies LF1 in Comer et al. Reference Comer, Slingerland, Krause, Iglesias, Clyde, Raigemborn and Wilf2015). Based on sedimentology and preservational quality, the potential order of transport distances for fossil leaves is ranked highest to lowest as LF, PL1, and PL2. Donovan et al. (Reference Donovan, Iglesias, Wilf, Labandeira and Cúneo2018) noted that preservation quality in Salamanca Formation locality PL1 is similar to that of the Lefipán Formation, indicating minimal preservation bias between the oldest Paleogene locality and the Maastrichtian collections in this study.
Analytical Methods
Macrofloral turnover across the K/Pg transition was addressed through analyses of (1) change in species composition, (2) morphospace shifts among assemblages, and (3) the potential biasing effect of the ca. 400 km separation between the Cretaceous and Paleogene collection sites using crosschecks from local pollen records in both formations (Fig. 1D). Preliminary Lefipán Formation morphotypes (Donovan et al. Reference Donovan, Iglesias, Wilf, Labandeira and Cúneo2016) were significantly revised and updated based on leaf architectural characters (Ellis et al. Reference Ellis, Daly, Hickey, Johnson, Mitchell, Wilf and Wing2009). Danian morphotypes of the Salamanca and Peñas Coloradas Formations are based on the morphotype set of the PL1, PL2, and LF macrofloral localities (Iglesias Reference Iglesias2007; Iglesias et al. Reference Iglesias, Wilf, Johnson, Cúneo and Matheos2007) and an additional set published in the supplementary information of Donovan et al. (2016; Supplementary Table 4). A total of 58 Maastrichtian and 43 Danian morphotypes were established and used in the subsequent analyses (Supplementary Table 4). Each morphotype is here interpreted as likely to have been produced by a single species with morphologically distinct leaves and will be referred to as a species throughout this study. An updated specimen-level collections inventory of all leaf morphotypes as used here is archived in Supplementary Table 3.
Extinction and Turnover
All Maastrichtian and Danian collections were grouped into two respective assemblages for macrofloral extinction estimates and morphospace analyses due to their self-similar floral compositions and morphospace occupation, based on preliminary analyses. Each of the Maastrichtian morphotypes was compared with the Danian morphotypes in search of K/Pg survivor pairs based on a detailed comparison of their leaf architecture (per Johnson et al. Reference Johnson, Nichols, Attrep and Orth1989; Ash et al. Reference Ash, Ellis, Hickey, Johnson, Wilf and Wing1999; Ellis et al. Reference Ellis, Daly, Hickey, Johnson, Mitchell, Wilf and Wing2009). Maastrichtian and Danian morphotypes that shared all morphological characters or that fell within each other's ranges of variation were considered survivor pairs.
Once the survivor pairs were established, they were each considered as a single species, along with all the other leaf species, in a principal components analysis (PCA) applied to a matrix of species abundances per sample, with the objective of testing compositional heterogeneity among the considered Maastrichtian and Danian macrofloral collections. In this analysis, Maastrichtian collections LefW and LefE+LefL were considered as two samples, because LefW was collected along several horizons within the same small canyon (see “Materials”), and LefE and LefL were collected along the same fossiliferous horizon.
For rarefied (Tipper Reference Tipper1979) species richness analysis across the K/Pg, the Maastrichtian LefE and LefL sites were grouped into a single sample, from which LefW was excluded because it includes specimens from multiple horizons with uncertain stratigraphic relationships to horizon LefE+LefL. The Salamanca (PL1 and PL2) and Peñas Coloradas (LF) localities (Fig. 1A,C,D) (Iglesias Reference Iglesias2007; Iglesias et al. Reference Iglesias, Wilf, Johnson, Cúneo and Matheos2007) were considered for rarefaction individually because they span three Danian time intervals (Fig. 1D) (see “Materials”; Clyde et al. Reference Clyde, Wilf, Iglesias, Slingerland, Barnum, Bijl, Bralower, Brinkhuis, Comer, Huber, Ibañez-Mejia, Jicha, Krause, Schueth, Singer, Raigemborn, Schmitz, Sluijs and Zamaloa2014; Comer et al. Reference Comer, Slingerland, Krause, Iglesias, Clyde, Raigemborn and Wilf2015). Each rarefaction curve was plotted with a 95% confidence interval using the R package iNext (Hsieh et al. Reference Hsieh, Ma and Chao2016).
As a baseline for comparison of rarefied species richness between Patagonian and North American K/Pg assemblages, three representative dicot-leaf samples surpassing 350 specimens each (per Burnham Reference Burnham1993) from meter-binned collections of the Maastrichtian Hell Creek and Danian Fort Union Formations in the WB of southwestern North Dakota were selected from the dataset of Wilf and Johnson (Reference Wilf and Johnson2004; see also Johnson Reference Johnson2002). The Dean Street level found 15 m below the K/Pg boundary (Wilf and Johnson Reference Wilf and Johnson2004) represents the most diverse, latest Maastrichtian HCIII zone (Johnson Reference Johnson2002). Two representative Fort Union Formation collection horizons were based on specimen counts >350 and magnetostratigraphic constraints. Horizon +7 m is the most specimen-rich representative of early Danian Fort Union Formation fossil horizons, corresponding to geomagnetic polarity chron C29r (which is unrecorded in Patagonia). The +38 m horizon is constrained to chron C29n (Hicks et al. Reference Hicks, Johnson, Obradovich, Tauxe and Clark2002; Johnson Reference Johnson2002), serving as a temporal analog to the PL1 locality in Patagonia (Fig. 1D) (Clyde et al. Reference Clyde, Wilf, Iglesias, Slingerland, Barnum, Bijl, Bralower, Brinkhuis, Comer, Huber, Ibañez-Mejia, Jicha, Krause, Schueth, Singer, Raigemborn, Schmitz, Sluijs and Zamaloa2014). While acknowledging work on diverse Paleocene floras from the more southerly Denver and San Juan basins (Johnson and Ellis Reference Johnson and Ellis2002; Flynn and Peppe Reference Flynn and Peppe2019; Lyson et al. Reference Lyson, Miller, Bercovici, Weissenburger, Fuentes, Clyde, Hagadorn, Butrim, Johnson, Fleming, Barclay, Maccracken, Lloyd, Wilson, Krause and Chester2019), we focus on the floras of the WB for comparison, because they come from similar absolute paleolatitudes to our samples (i.e., ca. 51.3°S for the Danian Patagonian floras, ca. 48.4°S for the Cretaceous Lefipán floras, and ca. 49.4°N for the WB floras, versus ca. 42.5°N for the Denver Basin, following Iglesias et al. Reference Iglesias, Wilf, Johnson, Cúneo and Matheos2007: fig. 3), contain both Cretaceous and Paleocene floras that are well sampled in a single area, and remain the best-sampled and described boundary-spanning K/Pg macrofloras of NAM.
Morphospace Analysis
Species richness and morphological diversity are not always coupled, and this disconnection may be driven by underlying selective pressures (Foote Reference Foote1993; Roy and Foote Reference Roy and Foote1997). Morphospace analysis is here used as a quantitative tool to measure morphological diversity and phenotypic relationships among angiosperm leaf species and their change through time, by summarizing morphological characters in a mathematical space where phenotypic dissimilarities are observed as graphical distances (Mitteroecker and Huttegger Reference Mitteroecker and Huttegger2009). Leaf architecture is related both to systematic affinities and environmental parameters (e.g., Wolfe Reference Wolfe1995; Wilf Reference Wilf1997; Little et al. Reference Little, Kembel and Wilf2010; Givnish and Kriebel Reference Givnish and Kriebel2017), making leaves ideal candidates to simultaneously test species and morphological turnover across the K/Pg. Through morphospace analysis, the multivariate relationships among characters can shed light on selective morphological extinction patterns, if present, and the relationships between morphological disparity and species richness.
Based on the Manual of Leaf Architecture (Ellis et al. Reference Ellis, Daly, Hickey, Johnson, Mitchell, Wilf and Wing2009), forty-six discrete shape, size, margin, and venation characters (Supplementary Table 5) were scored for each of the Maastrichtian and Danian morphotypes (Supplementary Table 7). Features that were not preserved (or observable) in at least three-fourths of the specimens, such as fourth- and higher-order venation, were excluded to reduce noise in the dataset. After observation of all available material, character states for each morphotype were usually based on one or two (if a single specimen did not preserve all morphological characters) selected exemplar specimens. If a morphotype showed significant variation in one or more characters after observation of all available samples, end members were separated in the morphological matrix designated by letters following the morphotype name (e.g., SA9A, SA9B; Supplementary Table 7) to ensure that all morphological diversity was captured in the subsequent ordination. Maximum size was based on the largest specimen of each species in the collection. Missing characters were coded as “NA.”
The morphological matrix (Supplementary Table 7) was ordinated using principal coordinates analysis (PCoA) with the R Package ape (Paradis and Schliep Reference Paradis and Schliep2018). Because it compares each morphotype on a character-by-character basis, PCoA is considered the best-performing method for datasets with uneven preservation and missing characters (Foote Reference Foote1994; Roy and Foote Reference Roy and Foote1997). The percentages of missing data for each character and each morphotype are reported in Supplementary Table 8. To understand how species are distributed in space and to explore possible character-related patterns, the same scores obtained in the PCoA ordination were analyzed by graphically superimposing character states on plotted scores including axes PCoA 1–PCoA 3. The process was repeated for each character.
To obtain a quantitative measure of morphospace occupation for Maastrichtian and Danian assemblages and to verify consistency in the results, four disparity measures were calculated separately for each Maastrichtian (LefW, LefE, and LefL) and Danian (PL1, PL2, and LF) leaf assemblage and for grouped Maastrichtian (LefW+LefE+LefL) and Danian (PL1+PL2+LF) assemblages. Based on the first four axes of the PCoA, (1) a hypercuboid volume and (2) a sum of ranges were estimated (Foote Reference Foote1994; Wills et al. Reference Wills, Briggs and Fortey1994). Based on the Euclidean pairwise distance matrices, the (3) average pairwise distance and (4) maximum pairwise distance were extracted. The hypercuboid volume was estimated by multiplying the ranges of the first four axes (Wills et al. Reference Wills, Briggs and Fortey1994). Following Gerber's (Reference Gerber2019) proposed measures of disparity for discrete character spaces, the (3) average and (4) maximum pairwise distances of the Euclidean distance matrix are reported here (Supplementary Table 6) for each local assemblage and the grouped Maastrichtian and Danian assemblages.
Pollen Analysis and Regional Floral Heterogeneity
Palynological records from the same formations and sections offer the closest possible points of comparison to macrofloras, and they provide, for the most part with taxonomic resolution above the species level, information about changes in regional floral composition (Behrensmeyer et al. Reference Behrensmeyer, Kidwell and Gastaldo2000; Nichols and Johnson Reference Nichols and Johnson2008). To evaluate the effects of the approximately 400 km distance separating the Maastrichtian and Danian localities (Fig. 1A) as a spatial bias affecting macrofloral composition, published palynological records for the same two sampling areas and stratigraphic framework (Fig. 1D) (Barreda et al. Reference Barreda, Cúneo, Wilf, Currano, Scasso and Brinkhuis2012; Clyde et al. Reference Clyde, Wilf, Iglesias, Slingerland, Barnum, Bijl, Bralower, Brinkhuis, Comer, Huber, Ibañez-Mejia, Jicha, Krause, Schueth, Singer, Raigemborn, Schmitz, Sluijs and Zamaloa2014) were compared at taxon-by-taxon and whole-assemblage levels. The Lefipán Formation palynological records from the SRS span the boundary (Fig. 1D) (unlike the Lefipán macrofloras; Barreda et al. Reference Barreda, Cúneo, Wilf, Currano, Scasso and Brinkhuis2012); thus, both their Maastrichtian and Danian components were compared with the exclusively Danian records of the Salamanca Formation at Palacio de los Loros and adjacent areas sampled at high stratigraphic resolution (Clyde et al. Reference Clyde, Wilf, Iglesias, Slingerland, Barnum, Bijl, Bralower, Brinkhuis, Comer, Huber, Ibañez-Mejia, Jicha, Krause, Schueth, Singer, Raigemborn, Schmitz, Sluijs and Zamaloa2014). We note that several other palynological studies exist of both formations based on other sections (e.g., Archangelsky Reference Archangelsky1973; Petriella and Archangelsky Reference Petriella and Archangelsky1975; Medina and Camacho Reference Medina and Camacho1990; Baldoni et al. Reference Baldoni, Askin and Baldoni1993; Casal et al. Reference Casal, Allard and Foix2015), but our goal is to compare palynological data from the same well-constrained sections as the macrofloras addressed here. The fine temporal correlations between the Lefipán and Salamanca Formations are not yet established.
V. Barreda (Museo Argentino de Ciencias Naturales, Buenos Aires) kindly reviewed the illustrations and species lists in Clyde et al. (Reference Clyde, Wilf, Iglesias, Slingerland, Barnum, Bijl, Bralower, Brinkhuis, Comer, Huber, Ibañez-Mejia, Jicha, Krause, Schueth, Singer, Raigemborn, Schmitz, Sluijs and Zamaloa2014) to establish equivalencies between palynotaxa (e.g., “Proteaceae sp. A” and “Proteaceae sp. B”) among the Lefipán and Salamanca sections, and her vetted species data were used in subsequent analyses (Supplementary Table 2). Sample-level compositional comparisons were based on a presence–absence matrix of palynomorph species and morphotypes (Supplementary Table 2) and processed in the R vegan package (Oksanen et al. Reference Oksanen, Blanchet, Friendly, Kindt, Legendre, McGlinn, Minchin, O'Hara, Simpson, Solymos, Stevens, Szoecs and Wagner2017).
The presence–absence matrix of palynomorph data was transformed using the Beals smoothing method (McCune Reference McCune1994; Münzbergová and Herben Reference Münzbergová and Herben2004), which accounts for unevenness in ecological sampling by replacing species’ presence data with a probability of occurrence based on co-occurrences in the sample pool. This method was used to reduce the underlying compositional heterogeneity caused by the inherent sampling bias of the fossil record. The samples were then clustered based on their Bray-Curtis pairwise dissimilarity matrix calculated from the Beals-smoothed dataset and linked using Ward's method algorithm (Oksanen et al. Reference Oksanen, Blanchet, Friendly, Kindt, Legendre, McGlinn, Minchin, O'Hara, Simpson, Solymos, Stevens, Szoecs and Wagner2017).
Results
Extinction and Turnover
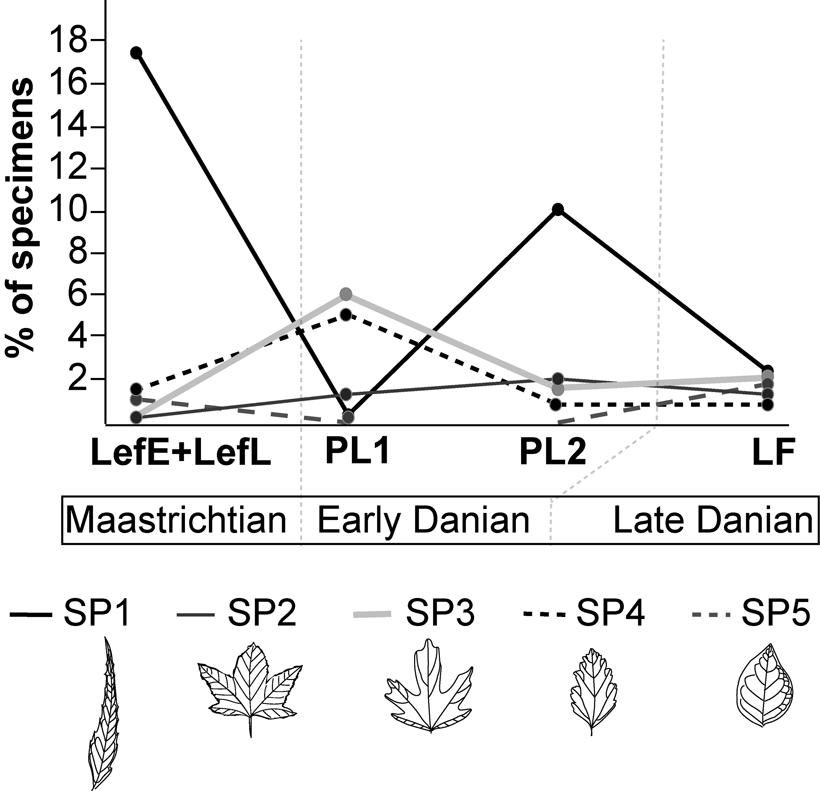
We recognize only five Maastrichtian leaf species as having a corresponding survivor in any Danian assemblage (Figs. 2, 3), indicating a raw dicot macrofloral extinction of 92.2%. Using only the oldest Danian assemblage (PL1 locality) yields a 93.3% extinction. Excluding species represented by a single specimen (singletons) reduces the extinction slightly, to 90.6%. The K/Pg survivor pairs based on Maastrichtian (prefix LEF) and Danian (prefix SA) morphotypes (Supplementary Tables 1,2) are referred to as Survivor Pair (SP) 1 (LEF57-SA20; Fig. 2A–E), SP2 (LEF64-SA35; Fig. 2F–I), SP3 (LEF6-SA19; Fig. 3A–C), SP4 (LEF18-SA08; Fig. 3D–G), and SP5 (LEF55-SA78; Fig. 3H–J).

Figure 2. K/Pg survivor pairs (SPs) from the Maastrichtian Lefipán (blue circles) and Danian (unmarked) Salamanca and Peñas Coloradas Formations (see also Fig. 3). A–E, SP1, Cunoniaceae, including articulated compound leaf. Note ovate and asymmetrical blades of variable leaflets, curved primary vein, craspedodromous secondary venation, mixed percurrent tertiaries perpendicular to secondaries, and small triangular teeth with long-rounded sinuses. A, MPEF-Pb-4416 from locality LefE; B, MPEF-Pb-4349, LefE; C, MPEF-Pb-9154 from locality PL2; D, MPEF-Pb-3691, PL1; E, MPEF-Pb-3694, PL1. F–I, SP2. Note well-developed palmate lobing, toothed margin, straight secondaries with opposite insertion, compound agrophic veins, and alternate percurrent tertiaries. F, MPEF-Pb-3701a, LefL; G, MPEF-Pb-2031, PL1; H, MPEF-Pb-3701b detail of alternate percurrent venation; I, MPEF-Pb-2031 (also H) detail of alternate percurrent venation. (Color online.)

Figure 3. K/Pg survivor pairs (SPs; continued from Fig. 2) from the Maastrichtian Lefipán (blue circles) and Danian (unmarked) Salamanca and Peñas Coloradas Fms. A–C, SP3, “Sterculia” acuminataloba. Note the palmately lobed form with entire margins, three strong primary veins, central primary deflected by strong basal secondaries, interior secondaries, intersecondary veins, agrophic veins, irregularly branching brochidodromous secondaries with spacing decreasing apically, and percurrent tertiaries nearly perpendicular to the primary vein. A, MPEF-Pb-4662a from locality LefW; B, MPEF-Pb-3692, PL1; C, MPEF-Pb-3695, PL2. D–G, SP4, Rosaceae similar to Crataegus spp. Note ovate shape, craspedodromous secondaries, pinnate lobing with toothed lobes, large triangular teeth, opposite to subopposite secondaries. D, MPEF-Pb-4487, LefE; E, MPEF-Pb-4482, LefE; F, MPEF-Pb-3693, PL1; G, MPEF-Pb-4030, PL1. H–J, SP5. Note ovate-elliptic leaf shape, opposite to subopposite concave-upward brochidodromous secondaries, intersecondary veins perpendicular to primary, widely spaced irregularly angled tertiaries ranging from opposite to alternate percurrent, and fimbrial vein running along entire margin. H, MPEF-Pb-4870, LefL; I, MPEF-Pb-4835, LefE; J, MPEF-Pb-3019, LF. (Color online.)
We interpret SP1 (Fig. 2A–E) to have a botanical affinity with the family Cunoniaceae based on its architectural characters including compound leaves (Fig. 2), supported by the well-documented presence (and earliest global macrofossil occurrence) of this family from abundant co-occurring Lacinipetalum spectabilis flowers with in situ pollen (schizomerioid Cunoniaceae; Jud et al. 2018) from PL2 and other Salamanca Formation localities and fossil wood in the Peñas Coloradas Formation (Raigemborn et al. Reference Raigemborn, Brea, Zucol and Matheos2009). The affinities of SP2 (Fig. 2F–I) and SP5 (Fig. 3H–J) remain unknown. Following Iglesias et al. (Reference Iglesias, Wilf, Johnson, Cúneo and Matheos2007), SP3 (Fig. 3A–C) is equivalent to some specimens designated as “Sterculia” acuminataloba Berry Reference Berry1937 of the family Malvaceae, and SP4 (Fig. 3D–G) belongs to the family Rosaceae based on diagnostic characters including serrated margins with compound teeth and craspedodromous secondary venation. The survivorship of the family Rosaceae is a new contribution of this study, along with Cretaceous macrofloral evidence to support the K/Pg survival of Malvaceae and Cunoniaceae, previously known from pollen records in these sections (Barreda et al. Reference Barreda, Cúneo, Wilf, Currano, Scasso and Brinkhuis2012; Jud et al. Reference Jud, Gandolfo, Iglesias and Wilf2018a,Reference Jud, Iglesias, Wilf and Gandolfob).
Laurophyll morphotypes observed in this study provide evidence for the K/Pg survival of the Lauraceae in Patagonia, although we cannot assign any Maastrichtian laurophyll to a definite Danian species-level equivalent because they lack cuticular preservation for further analysis (e.g., Carpenter et al. Reference Carpenter, Iglesias and Wilf2018). Based on a conservative “lumping” approach, we established four Maastrichtian laurophyll morphotypes based on their having typical lauraceous leaf architectural features (after Hickey and Wolfe Reference Hickey and Wolfe1975). Lefipán Formation morphotypes LEF08, LEF24, LEF26, and LEF32 (Supplementary Table 3), coupled with Danian Laurophyllum piatnitzkyi Berry and Laurophyllum chubutensis Berry of the Salamanca and Peñas Coloradas Formations (Iglesias et al. Reference Iglesias, Wilf, Johnson, Cúneo and Matheos2007), add Lauraceae to the list of Patagonian K/Pg macrofloral survivor families. Laurophyllum piatnitzkyi is notable as the most abundant species in the Danian assemblages, accounting for ca. 17% of total leaf specimens (Iglesias et al. Reference Iglesias, Wilf, Johnson, Cúneo and Matheos2007).
The loss of common taxa followed by the emergence of previously rare taxa is a commonly observed pattern across mass extinction intervals (e.g., Erwin Reference Erwin2002; Johnson Reference Johnson2002; Jablonski Reference Jablonski2005). A comparable pattern occurs in our survivor pairs (Fig. 4). SP1 of the Cunoniaceae (Fig. 2A–E) is a major component of Maastrichtian floras, but its abundance is greatly reduced at earliest Danian PL1 before partially recovering in PL2 (Fig. 4). In contrast, Malvaceae SP3 (Fig. 3A–C) and Rosaceae SP4 (Fig. 3D–G), both minor components in Maastrichtian assemblages, surpass 5% of assemblage composition at early Danian PL1 and are reduced back to minor components at PL2 and LF (Fig. 4). Leaf morphotypes SP2 and SP5, of unknown affinities, remain minor components throughout.
Rarefied dicot-leaf richness dropped in Patagonia by almost 40% across the K/Pg and remained comparably low from the early to late Danian (Fig. 5). Patagonian Danian floras are remarkably homogeneous across time and facies, from early to late Danian and from marginal marine to pyroclastic–fluvial environments, in comparison with the relatively more heterogeneous Lefipán floras deposited in similar facies and spanning a narrower time interval (Fig. 6). A sharp drop in rarefied species richness was also observed in the North American K/Pg floras of the WB in North Dakota (e.g., Johnson Reference Johnson2002; Wilf and Johnson Reference Wilf and Johnson2004). However, our rarefaction also shows that Patagonian floras are significantly more diverse than the North Dakota floras not only during the early Danian (Iglesias et al. Reference Iglesias, Wilf, Johnson, Cúneo and Matheos2007) but also the Maastrichtian (Fig. 5).

Figure 5. Rarefied leaf species richness with 95% confidence intervals for Patagonian sites (solid lines, this study) and Williston Basin, North Dakota (dashed lines, denoted with meters below or above K/Pg impact layer; data from Wilf and Johnson Reference Wilf and Johnson2004). Data include all Maastrichtian (black) and Danian (gray) dicot-leaf samples, including singleton species (see “Methods” for details).

Figure 6. Compositional differences between Maastrichtian (K) and Danian (Pg) assemblages based on a species by abundances principal components analysis (PCA), showing increased similarity of floral composition from the Maastrichtian (Lef) to the Danian (PL, LF) macrofloras, even though the Danian assemblages represent a longer time interval and a much greater array of facies types. PCA 1 variance explained = 72.7%, PCA 2 variance explained = 11.5%.
Morphospace Analysis
Pre- and post-K/Pg leaf assemblages significantly overlap in morphospace, but there is an increase in morphospace occupation between the terminal Cretaceous and early Paleogene (Fig. 7). This result shows no significant loss of characters and higher morphological disparity in post- compared with pre-K/Pg assemblages, despite lower species richness in the Danian. Morphospace occupation across the K/Pg is characterized by three notable shifts. First, morphospace volume expands, driven by Danian lobed species displaying blade incision characters absent in the Maastrichtian. Examples include lobed species with toothed margins such as SA35 (see circled groups in Fig. 8A). Second, within the morphospace area shared by Maastrichtian–Danian assemblages (Fig. 8), more Danian species have lobed blades, toothed margins, and craspedodromous venation.

Figure 7. Percentage increase in leaf morphospace occupation from Maastrichtian to Danian in Patagonia based on four measures of disparity (see “Methods” for details). A, Morphospace volume increase based on all Maastrichtian and Danian species. B, Morphospace area change partitioned by most common leaf size classes.

Figure 8. Morphospace occupation of Maastrichtian and Danian assemblages from principal coordinates analysis (PCoA). The expanded Danian morphospace (see Fig. 7) indicates higher morphological diversity in post-K/Pg floras despite lower species richness. A, Axis 2 vs. axis 1. Characteristically lobed outliers driving Danian morphospace expansion are recognized as two groups as circled; labeled exemplar species are: 1, SA35; 2, SA39; 3, SA55; 4, SA19A; 5, SA19B. B, Axis 3 vs. axis 1. Morphological end members along PCoA 1, both Danian, labeled: 1, SA35; 2, SA50.
Third, extremes in morphological variation, seen as end members along the PCoA axes, are all represented by Danian species, signaling higher morphological disparity in post-K/Pg assemblages (Fig. 8). For example, with the lowest score along axis PCoA 1, Danian species SA35 (Fig. 8A, lower left) is lobed with craspedodromous secondary veins and toothed margins, whereas SA50 (Fig. 8B, upper right), with the highest score for axis PCoA 1, is unlobed with entire margins and brochidodromous venation. Intermediate morphotypes along PCoA 1 display combinations of entire and toothed-margined characters and additional, less common venation types. Maastrichtian morphotypes cluster toward the higher scores of PCoA 1 and most share morphological characteristics with SA50, such as entire margins, no lobes, and brochidodromous secondaries (Fig. 8).
When we analyzed morphospace occupation by character states (Supplementary Table 5), the most significant observable variation was found in characters of maximum blade size, lobation, and margin types. Maximum leaf size ranges from nanophyll to megaphyll, with representatives of both Maastrichtian and Danian species in each size class. The representation of both Maastrichtian and Danian leaves within the same size class range could indicate isotaphonomic leaf assemblages. However, for purposes of morphospace analysis, leaves in the smallest (nanophyll) and largest (macrophyll and megaphyll) size classes were excluded, because they included too few specimens to produce an interpretable morphospace occupation pattern. Individually plotting the ordination scores by maximum leaf size classes of each species (Fig. 9), we found that shifts in morphospace occupation across the K/Pg become more significant as maximum leaf size increases (Figs. 7, 9).

Figure 9. Morphospace occupation (axis 2 vs. axis 1) partitioned by maximum leaf size per species using the same PCoA scores for each species shown in Fig. 8. Microphyll is the smallest size class present; mesophyll is the largest. A, Microphylls, selected end-member morphotypes with icons are 1, LEF16; 2, LEF20; 3, LEF46; 4, LEF34. B, Notophylls, examples are 1, LEF64; 2, LEF01; 3, SA50; 4, LEF05. C, Mesophylls, examples are 1, SA35; 2, SA74; 3, LEF12; 4, SA73; 5, LEF07; 6, LEF26; 7, SA55.
Within each size class, there appear to be distinct morphological trends (Fig. 9). In the smallest size classes, microphylls and notophylls, Maastrichtian leaves occupy a larger morphospace than Danian species, as shown by the four end members along axes 1 and 2 (Fig. 9A, labeled 1–4; Fig. 9B, labeled 1, 2, and 4). This result is supported by a reduction in morphospace volume (Fig. 7B), indicating that leaves in these size classes are more morphologically diverse in Maastrichtian assemblages. In contrast, the larger mesophyll-sized leaves (Fig. 9C) expand in morphospace occupation after the K/Pg (Fig. 7B). Danian mesophylls span a wide range of morphologically diverse lobed, unlobed, entire-margined, and toothed morphotypes. Mesophyll Maastrichtian morphotypes are generally entire-margined and share self-similar secondary venation patterns (e.g., labels 5 and 6, Fig. 9C).
We found distinct morphospaces occupied by lobed versus unlobed species, whether we included all species (Fig. 10A), Maastrichtian-only (Fig. 10B), or Danian-only (Fig. 10C) assemblages. Lobed leaves are consistently more morphologically disparate than unlobed types (Fig. 10B,C), a notable result considering that presence of lobes is only one of the more than 40 characters weighted equally in the ordination. Lobation in Danian species is generally associated with non-entire margins and craspedodromous venation, a relatively uncommon combination in Maastrichtian assemblages.

Figure 10. Principal coordinates analysis (PCoA axis 2 vs. axis 1) coded by time period, lobed vs. unlobed species (A–C), and toothed vs. entire-margined species (D–F). Species ordination scores are the same as and extracted from the analysis shown in Fig. 8. A–C, Lobed species lead the morphospace increase from Maastrichtian to Danian assemblages. D–F, Toothed and entire-margined species occupy distinct areas of morphospace in all assemblages and increase in separation in Danian assemblages, indicating a loss of intermediate morphospace across the extinction interval. First column shows morphospace occupation by all species, colored by respective character. Second column shows morphospace distribution of Maastrichtian species only. Exemplars in B, 1, LEF64; 2, LEF66; 3, LEF46; 4, LEF05; and E, 1, LEF64; 2, LEF53; 3, LEF7; 4, LEF46. Third column shows morphospace distribution of Danian species only. Exemplars in C, 1, SA35; 2, SA48; 3, SA50; 4, SA39; and F, 1, SA35; 2, SA50.
Regarding margin characters, in both Maastrichtian and Danian assemblages, toothed and entire-margined morphotypes occupy distinct morphospaces (Fig. 10D), presumably in large part a result of the numerous characters in the matrix that pertain to toothed margins that are coded as zeroes for entire-margined morphotypes. However, there is some overlap among toothed and entire-margined types in Maastrichtian assemblages, characterized by species with margins scored as crenate and a few entire-margined examples (Fig. 10E). Interestingly, there is no such overlap in Danian assemblages (Fig. 10F), graphically showing the loss of crenate margins from pre- to post-K/Pg assemblages and indicating one of the few morphological losses.
Pollen Analysis and Floral Heterogeneity among Sections
Comparison of published pollen records from the same sections as the macrofloral localities (Fig. 1D) shows that the Maastrichtian–Danian Lefipán and Danian-only Salamanca Formation samples (Fig. 1A) share most of the same pollen species and thus the same higher taxa (families and possibly genera), whether or not the Maastrichtian pollen data are included. First, in the closest stratigraphic comparison with the leaf data, approximately 60% of palynospecies in the Danian Salamanca Formation are shared with the Maastrichtian part of the Lefipán Formation (Supplementary Table 2). Second, the Salamanca Formation also shares about 75% of palynospecies with the Danian strata of the Lefipán (Barreda et al. Reference Barreda, Cúneo, Wilf, Currano, Scasso and Brinkhuis2012; Supplementary Table 2). Thus, the same higher plant taxa are inferred to be present in the Danian microfloras of both formations (Fig. 1), despite the high species-level turnover indicated by leaves. Third, the percentage of shared palynotaxa among the Maastrichtian Lefipán and the Danian Salamanca (i.e., the pollen analog to the macrofloras studied here; Fig. 1) is similar to that among Maastrichtian and Danian Lefipán Formation samples within the SRS.
These palynological results all suggest that geographic separation does not significantly influence the composition of higher taxa between the two localities, supporting the comparability of the Maastrichtian and Danian macrofloras undertaken here. Considering all the Danian palynotaxa (Lefipán and Salamanca), 80% are Cretaceous–Paleogene survivors based on the Cretaceous Lefipán as the source of survivors for both formations, demonstrating the persistence of taxa including the gymnosperm families Podocarpaceae and Classopollis sp. (Cheirolepidiaceae) and the angiosperm families Arecaceae, Liliaceae, Proteaceae, Symplocaceae, and Gunneraceae, as well as various bryophytes and ferns (Supplementary Tables 1, 2). Cluster analysis of the Beals smoothed compositional matrix (Fig. 11) shows that despite the high percentage of palynomorph species shared between the two formations, compositional differences associated with age and formation can be recognized.

Figure 11. Cluster analyses of palynospecies occurrences with Beals smoothing (see “Methods” and Fig. 1) from the Maastrichtian (samples M1–3) and Danian (D1–11) Lefipán Fm. (Barreda et al. Reference Barreda, Cúneo, Wilf, Currano, Scasso and Brinkhuis2012) and Danian-only Salamanca Fm. (remaining samples; Fig. 1) (Clyde et al. Reference Clyde, Wilf, Iglesias, Slingerland, Barnum, Bijl, Bralower, Brinkhuis, Comer, Huber, Ibañez-Mejia, Jicha, Krause, Schueth, Singer, Raigemborn, Schmitz, Sluijs and Zamaloa2014). A, All species. B, Gymnosperms and angiosperms only. C, Angiosperms only.
Discussion
A high angiosperm macrofloral extinction percentage, drop in rarefied species richness, depauperate and homogeneous early Paleocene floras (Fig. 6) despite spanning a range of continental to marginal facies (e.g., Comer et al. Reference Comer, Slingerland, Krause, Iglesias, Clyde, Raigemborn and Wilf2015), and significant extinction of specialized plant–insect associations in Patagonia (Donovan et al. Reference Donovan, Iglesias, Wilf, Labandeira and Cúneo2016) all provide evidence of a significant K/Pg floral extinction event as seen in NAM (Fig. 5) (Wolfe and Upchurch Reference Wolfe and Upchurch1986; Johnson and Hickey Reference Johnson and Hickey1990; Johnson Reference Johnson2002; Wilf and Johnson Reference Wilf and Johnson2004). Our results support a disruption in plant communities of global extent across the K/Pg (Vajda et al. Reference Vajda, Raine and Hollis2001; Nichols and Johnson Reference Nichols and Johnson2008; Barreda et al. Reference Barreda, Cúneo, Wilf, Currano, Scasso and Brinkhuis2012; Cantrill and Poole Reference Cantrill, Poole, Cantrill and Poole2012; Vajda and Bercovici Reference Vajda and Bercovici2014; Donovan et al. Reference Donovan, Iglesias, Wilf, Labandeira and Cúneo2016).
Lauraceae have recently been proposed as the principal component of K/Pg recovery macrofloras alongside the classic fern spike in the North American Ratón Basin (Berry Reference Berry2019), although Lauraceae are not reported from the well-sampled early Danian localities in North Dakota (Johnson Reference Johnson2002). The dominance of L. piatnitzkyi in early Paleocene Patagonian macrofloras, and of several other leaf morphotypes assigned to the family (Iglesias Reference Iglesias2007; Iglesias et al. Reference Iglesias, Wilf, Johnson, Cúneo and Matheos2007) would be consistent with the presence of Lauraceae-dominated K/Pg recovery floras in regions of both North and South America.
Despite the similarities between Patagonian and North American records, the subtle differences between them suggest that extinction could have been heterogeneous between the two regions. Morphological turnover shows interesting differences between the WB and Patagonia, even though the WB floras have not been analyzed in morphospace. Maastrichtian assemblages of North Dakota are characterized by the abundance of leaves with a “Cretaceous look” or unusual lobation (Johnson Reference Johnson2002: p. 371), a feature that is drastically reduced in the early Paleocene (Johnson and Hickey Reference Johnson and Hickey1990; Johnson Reference Johnson2002). In contrast, the increase in proportion of lobed and toothed species in the early Paleocene of Patagonia (Fig. 8) (Iglesias et al. Reference Iglesias, Wilf, Johnson, Cúneo and Matheos2007) leads to an expansion in morphospace occupation by post-K/Pg assemblages. Despite our estimate of almost complete species-level macrofloral turnover in Patagonia, the continuity and expansion of leaf morphologies (Fig. 8) and, presumably, their underlying ecological and phylogenetic diversity across the K/Pg could, in part, reflect low turnover at higher taxonomic levels observed in both the microfloral and macrofloral records of Patagonia (Barreda et al. Reference Barreda, Cúneo, Wilf, Currano, Scasso and Brinkhuis2012; Supplementary Table 1).
Both WB and Patagonian species richness suffer a sharp drop between Maastrichtian and Danian assemblages (Fig. 5). The K/Pg drop in rarefied species richness, though severe in both areas, is much sharper in the WB (ca. 75%) than in Patagonia (ca. 45%; Fig. 5). Rarefaction highlights another important contrast between the WB and Patagonian records; not only Danian (Iglesias et al. Reference Iglesias, Wilf, Johnson, Cúneo and Matheos2007) but also Maastrichtian assemblages in Patagonia are significantly more diverse than coeval macrofloras in the WB. The Lefipán floras are approximately 40% richer than the most diverse Maastrichtian Hell Creek (HCIII) NAM zone assemblage (Fig. 5). Thus, whereas previous studies of the Danian Salamanca Formation (Iglesias Reference Iglesias2007; Iglesias et al. Reference Iglesias, Wilf, Johnson, Cúneo and Matheos2007) and Eocene macrofloras of Patagonia (Wilf et al. Reference Wilf, Cúneo, Johnson, Hicks, Wing and Obradovich2003a, Reference Wilf, Johnson, Cúneo, Smith, Singer and Gandolfo2005) provided evidence for an ancient history of high South American Cenozoic floral diversity, our results (Fig. 5) extend that history into the Late Cretaceous.
The high species richness of Patagonian Paleocene and Eocene assemblages compared with NAM equivalents has been hypothesized in part to be the legacy of low K/Pg floral extinction as observed in the palynological record (Barreda et al. Reference Barreda, Cúneo, Wilf, Currano, Scasso and Brinkhuis2012). Our results suggest that the rich Cenozoic macrofloras of Patagonia also carry a legacy of rich Cretaceous floras, as well as significantly earlier recovery of Paleocene species richness resulting from the persistence of most higher taxonomic levels (Supplementary Tables 1, 2). Nonetheless, an emerging body of research shows that floral diversity in the early Paleocene of the Denver and San Juan Basins can be higher relative to the more northerly WB (Johnson and Ellis Reference Johnson and Ellis2002; Flynn and Peppe Reference Flynn and Peppe2019; Lyson et al. Reference Lyson, Miller, Bercovici, Weissenburger, Fuentes, Clyde, Hagadorn, Butrim, Johnson, Fleming, Barclay, Maccracken, Lloyd, Wilson, Krause and Chester2019). These studies show heterogeneity in NAM floral richness during the early Paleocene, a pattern warranting further intensive study.
Insect damage-type data on the same leaf collections studied here showed a similar pattern of high extinction and robust early recovery compared with NAM (Donovan et al. Reference Donovan, Iglesias, Wilf, Labandeira and Cúneo2016, Reference Donovan, Iglesias, Wilf, Labandeira and Cúneo2018). Donovan et al. (Reference Donovan, Iglesias, Wilf, Labandeira and Cúneo2016, Reference Donovan, Iglesias, Wilf, Labandeira and Cúneo2018) posed the disappearance of plant hosts across the K/Pg as a potential driving mechanism for the severe insect herbivore extinction observed in Patagonia; host-specialized insects such as leaf miners would have been severely affected by an elevated species-level floral extinction (as in NAM: Labandeira et al. Reference Labandeira, Johnson and Wilf2002; Donovan et al. Reference Donovan, Wilf, Labandeira, Johnson and Peppe2014). Our results confirm this interpretation and suggest that despite the low K/Pg turnover of major clades observed in floras of Patagonia and elsewhere (see Sauquet and Magallón Reference Sauquet and Magallón2018), high plant-species turnover resulted in devastating consequences for terrestrial faunas.
The empirical species-extinction percentage of >90% in Patagonia in all likelihood is positively biased by several factors, including coarser stratigraphic sampling than in WB macrofloras, geographic variation, and climatic effects, and that is why we present it as a maximum estimate. Regarding sampling, the uncertain temporal placement of the Lefipán Formation macrofloras within the last ca. million years before the K/Pg boundary (Barreda et al. Reference Barreda, Cúneo, Wilf, Currano, Scasso and Brinkhuis2012; Vellekoop et al. Reference Vellekoop, Holwerda, Prámparo, Willmott, Schouten, Cúneo, Scasso and Brinkhuis2017) and the lack of earliest Danian (chron 29r) sediments preserved in the Salamanca Formation probably bias against finding a higher number of survivor pairs (Fig. 1). The limited stratigraphic coverage of macrofloras in the upper Lefipán Formation is analogous to considering only a small interval of the Hell Creek Formation (Johnson et al. Reference Johnson, Nichols, Attrep and Orth1989; Johnson Reference Johnson2002; Wilf and Johnson Reference Wilf and Johnson2004), where there is a general trend of lower percentages of observed survivor dicot-leaf species in a simple experiment where the distance of sample windows below the boundary layer increases (Fig. 12). Furthermore, observed extinction may exceed 90% even for some intervals relatively close to the boundary, as can be seen in the 20–30 m and 40–50 m bins of the Hell Creek Formation (Fig. 12). Although there are macrofloral collections from lower Lefipán Formation strata (Andruchow-Colombo et al. Reference Andruchow-Colombo, Escapa, Cúneo and Gandolfo2018; Martínez et al. Reference Martínez, Gandolfo and Cúneo2018), their precise age is not known, and the preserved diversity of dicot leaves is not nearly so extensive as in the upper Lefipán studied here. Therefore, observations across a wide stratigraphic range in the Lefipán Formation are not possible at this time.

Figure 12. Simulated percentage of K/Pg-surviving Cretaceous dicot-leaf species in the Williston Basin of North Dakota by 10 m bin window below the K/Pg impact horizon (data from Wilf and Johnson [Reference Wilf and Johnson2004], including singletons; see “Discussion” for details). The Lefipán leaf floras studied here, with a 91% observed extinction, could correlate temporally to any one of these stratigraphic bins.
Addressing the geographic separation between the Maastrichtian and Danian macrofloras (Fig. 1), the cluster results showing differences between the Lefipán and Salamanca sections’ microfloral assemblages (Fig. 11) provide evidence for some geographical heterogeneity of higher-taxa associations, which would accentuate species-level contrasts observed in the respective macrofloras (Behrensmeyer et al. Reference Behrensmeyer, Kidwell and Gastaldo2000; Nichols and Johnson Reference Nichols and Johnson2008). Different underlying palynospecies associations suggest that the 400 km spatial separation of Maastrichtian and Danian assemblages plays some role in the likely overestimation of Patagonian macrofloral extinction by imparting a geographical bias (Fig. 11). On the other hand, geographic effects could not have been severe, because the number of shared palynospecies in the same strata as the leaf floras was very high (see “Results”).
With regard to climate, marine and continental proxy records show a global relatively short-lived climatic warming within the last ca. 500 kyr of the Maastrichtian, followed by cooling during the final ca. 100 kyr of the Maastrichtian persisting into the early Paleocene (Stott et al. Reference Stott, Kennett, Shackleton and Corfield1990; Huber and Watkins Reference Huber and Watkins1992; Wilf et al. Reference Wilf, Johnson and Huber2003b; Bowman et al. Reference Bowman, Francis, Askin, Riding and Swindles2014; Vellekoop et al. Reference Vellekoop, Holwerda, Prámparo, Willmott, Schouten, Cúneo, Scasso and Brinkhuis2017; Woelders et al. Reference Woelders, Vellekoop, Kroon, Smith, Casadío, Prámparo, Dinares-Turell, Peterse, Sluijs, Lenaerts and Speijer2017; Huber et al. Reference Huber, MacLeod, Watkins and Coffin2018; Hull et al. Reference Hull, Bornemann, Penman, Henehan, Norris, Wilson, Blum, Alegret, Batenburg, Bown, Bralower, Cournede, Deutsch, Donner, Friedrich, Jehle, Kim, Kroon, Lippert, Loroch, Moebius, Moriya, Peppe, Ravizza, Röhl, Schueth, Sepúlveda, Sexton, Sibert, Sliwinska, Summons, Thomas, Westerhold, Whiteside, Yamaguchi and Zachos2020). The increase in toothed and lobed leaf morphotypes across the K/Pg in Patagonia (39.7% to 60.5% and 12% to 27.9%, respectively), both of which are observed drivers of morphospace expansion in Paleogene floras, would correlate to a ca. 5°C drop in mean annual temperature, with standard caveats for paleoclimatic inference on this basis (Wolfe Reference Wolfe1971; Greenwood et al. Reference Greenwood, Wing and Christophel2004; Little et al. Reference Little, Kembel and Wilf2010). The uncertain age within the last million years of the Maastrichtian of the Lefipán Formation macrofloras studied here, however, introduces an important potential climatic bias on observed extinction that would compound the sampling bias mentioned earlier.
In the Hell Creek Formation, peak richness of floral diversity (HCIII zone flora) correlates with the highest temperature estimates of the late Maastrichtian, followed by a decline in species richness during rapid cooling in the terminal Maastrichtian (Wilf et al. Reference Wilf, Johnson and Huber2003b). If a comparison were to be made solely between a collection of Maastrichtian HCIII floras from the peak of diversity of warmth and those of the Fort Union, an overestimation of extinction percentages would result. For example, if we only compare the −15 m (HCIII) and +38 m (in C29N like the PL1 flora) horizons in North Dakota (Wilf et al. Reference Wilf, Johnson and Huber2003b; Wilf and Johnson Reference Wilf and Johnson2004), the observed dicot-leaf extinction is 99%. Although it is not possible to state the age of the Maastrichtian Lefipán Formation floras at this precision, their high species richness, dinoflagellate markers indicative of the last 0.5 Myr of the Cretaceous, and associated warm proxy temperatures (Barreda et al. Reference Barreda, Cúneo, Wilf, Currano, Scasso and Brinkhuis2012; Scasso et al. Reference Scasso, Aberhan, Ruiz, Weidemeyer, Medina and Kiessling2012; Vellekoop et al. Reference Vellekoop, Holwerda, Prámparo, Willmott, Schouten, Cúneo, Scasso and Brinkhuis2017) suggest that they could correlate to the terminal Cretaceous warming event and thus HCIII, possibly translating into an overestimation of macrofloral extinction in this study on the assumption that temperature correlates with plant diversity.
Conclusions
We report the first K/Pg macrofloral investigation in the Southern Hemisphere that is based on large, stratigraphically constrained collections. Our 92% observed macrofloral extinction is best viewed as a maximum because of likely geographic, stratigraphic, and paleoclimatic biases. Nonetheless, our estimate clearly reflects underlying high species-level turnover in Patagonia. High macrofloral turnover and associated insect herbivore extinction, compositionally homogeneous Danian floras, and a sharp drop in rarefied floral species richness in Patagonia reveal extinction and recovery patterns broadly resembling those observed in NAM and support a geographically widespread extinction event. However, Patagonian and North American turnover show contrasts.
Compared with NAM floras of the WB in North Dakota, Patagonian floras are much more diverse in both Maastrichtian (Fig. 5) and Danian (Iglesias et al. Reference Iglesias, Wilf, Johnson, Cúneo and Matheos2007) assemblages. Richer Maastrichtian floras in Patagonia may have contributed to the less severe drop in overall species richness and significantly faster Danian recovery than observed in the WB or other northern Great Plains floras. Insect herbivory observations on the same collections (Donovan et al. Reference Donovan, Iglesias, Wilf, Labandeira and Cúneo2016, Reference Donovan, Iglesias, Wilf, Labandeira and Cúneo2018) follow patterns similar to North American records, indicating that the macrofloral species extinction in Patagonia had catastrophic effects on herbivorous insect communities. However, like their host macrofloras, specialized insect damage types in Patagonia recover much faster than those in NAM, despite high turnover in both regions.
This study reports speciose Maastrichtian macrofloras in Patagonia for the first time, extending the history of elevated floral diversity in South America into the terminal Cretaceous. The limited palynological extinction and continuity of most higher taxa (families and genera) across time and space indicate persistence of major plant clades even as significant extinction took place at the species level. Leaves with lauraceous affinity are characteristically dominant in the Patagonian macrofloral assemblages, as in the NAM Ratón Basin, drawing an interesting parallel between the two regions. In addition to Lauraceae, we provide macrofloral evidence to support the survival of the families Cunoniaceae, Malvaceae, and Rosaceae in Patagonia.
Morphospace analysis of Maastrichtian–Danian Patagonian macrofloras shows minimal character loss from the K/Pg extinction and that Danian macrofloras are morphologically more diverse than Maastrichtian assemblages. Danian floras rich in lobed and toothed species appear to be a notable Patagonian characteristic that contrasts with North American macrofloras of the WB, where lobed forms largely disappear across the K/Pg. Increased diversity of toothed species in Danian Patagonian assemblages is probably related to a general climatic cooling, potentially aligning with global temperature records.
Extinction and recovery in Patagonian macrofloras parallel North American WB assemblages in the K/Pg drop of rarefied species richness, low-diversity homogeneous Paleocene floras, and dramatic loss of specialized insect herbivores, indicating that terrestrial ecosystems in both areas were severely disrupted. Contrasts between Patagonian and North American macrofloras emerge in a faster Patagonian recovery, comparably much higher floral diversity throughout the Cretaceous–Paleogene time interval, and differences in morphological turnover.
Acknowledgments
We thank the technical staff of the Museo Paleontológico Egidio Feruglio (MEF), including P. Puerta, M. Caffa, E. Ruigomez, L. Reiner, and L. Canessa, as well as K. Johnson, R. Scasso, M. Donovan, and many others for their field and laboratory assistance over several expeditions and collections visits. We thank V. Barreda for her assistance reviewing palynological records and M. Patzkowsky and T. Bralower for valuable advice during the development of this project. We thank the editors, K. Boyce and C. Looy, and V. Vajda and three anonymous reviewers for providing constructive feedback that helped us improve this article. Funding was provided to E.S. by Geological Society of America Student Research grant no.12008-18, a Mid-American Paleontological Society (MAPS) Outstanding Research Award, the Penn State Geosciences Charles E. Knopf, Sr., Memorial Scholarship, and two Penn State Geosciences Paul D. Krynine Scholarships, as well as contributions from National Science Foundation grants DEB-15556666/1556136, EAR-1925755/1925552, DEB-0919071/0918932, and DEB-0345750 that benefited all authors. This work partially fulfilled the M.Sc. in Geosciences degree requirements of E.S. at Penn State University.













 Open access
Open access