



Introduction
Species introductions are a major conservation concern, presenting complex social and ecological challenges (Ehrenfeld, Reference Ehrenfeld2010; Gallardo et al., Reference Gallardo, Clavero, Sánchez and Vilà2016; Courchamp et al., Reference Courchamp, Fournier, Bellard, Bertelsmeier, Bonnaud, Jeschke and Russell2017). Concerns may be particularly pronounced when the species in question is known to have strong effects in its native range. Much literature is focused on the detrimental effects of introduced species, but there has been recent discussion of the potential role of introduced species to replace species lost through extinction (Dembitzer, Reference Dembitzer2017; Lundgren et al., Reference Lundgren, Ramp, Ripple and Wallach2018) and/or contribute to conservation through provision of ecosystem services (Schlaepfer et al., Reference Schlaepfer, Sax and Olden2011). This discussion reveals the complexity inherent in determining appropriate approaches to manage introduced species (Werner et al., Reference Werner, Cowie and Cusack2006; Kodric-Brown & Brown, Reference Kodric-Brown and Brown2007; García-Llorente et al., Reference García-Llorente, Martín-López, González, Alcorlo and Montes2008; Mascaro et al., Reference Mascaro, Hughes and Schnitzer2012; Svenning et al., Reference Svenning, Pedersen, Donlan, Ejrnæs, Faurby and Galetti2016; Svenning & Faurby, Reference Svenning and Faurby2017).
A recent introduction of the hippopotamus Hippopotamus amphibius into Colombia brings together many of the major research themes on species introductions. The hippopotamus played an important role in ancient Egyptian symbology, with females representing fertility and males representing chaos (Pinch, Reference Pinch2004). These are apt characterizations, as the species can have powerful and multifarious effects on its environment. In its native range the hippopotamus has high rates of reproduction and population growth (Martin, Reference Martin2005), is an ecosystem engineer that can have strong and complex effects on its environment, and can be dangerous and destructive (Eltringham, Reference Eltringham1999). These characteristics have been relatively well-studied in the species' native range, in which population declines as a result of poaching and habitat loss are the primary concerns (Lewison & Pluhacek, Reference Lewison and Pluhacek2017). The recent introduction of hippopotamuses into the wild in the Magdalena River basin of Colombia, the most populated basin in the country, raises questions about how the species will affect this ecosystem and its inhabitants (Dembitzer, Reference Dembitzer2017; Lundgren et al., Reference Lundgren, Ramp, Ripple and Wallach2018).
Here we synthesize the history of this introduction and interventions to date. We provide initial population estimates, predicted growth rates and a range map. We review published literature on the hippopotamus in its native range and in the Magdalena River basin, to discuss the potential ecological and socio-economic consequences of the introduction, and provide recommendations for further study.
Status of the hippopotamus in Colombia
In 1981 the drug trafficker Pablo Escobar imported four hippopotamuses (three females and one male), amongst other animals, from a zoo in the USA, for his private collection of exotic animals. The hippopotamuses lived in a pond on Escobar's ranch at Hacienda Nápoles (Plate 1). Escobar was killed in 1993, and most of the animals were distributed to Colombian zoos. The hippopotamuses, being difficult to transport, were left on the property, where they inhabited one primary pond. Hacienda Nápoles fell into disrepair, but in 2006 it was developed for tourism. By this time, there were an estimated 16 hippopotamuses living on the property. The population continued to grow, spreading into nearby aquatic environments (Fig. 1).

Plate 1 (a) A hippopotamus Hippopotamus amphibius approaching tourists for food at Hacienda Nápoles, and (b) a hippopotamus in a pond outside Hacienda Nápoles.

Fig. 1 Timeline of the introduction of the hippopotamus Hippopotamus amphibius to Colombia and dispersal into the Magdalena River basin. Distances in parentheses are straight line distances from Hacienda Nápoles, the point of introduction.
Hacienda Nápoles is located in rolling pastureland near the town of Doradal, 13.5 km west of the Magdalena River at an altitude of 240 m (Fig. 2). There are many floodplain lakes (known as ciénagas), man-made cattle ponds, and small streams throughout the landscape, which seem to be facilitating the species’ dispersal. In c. 2006 hippopotamuses were observed in ponds near Hacienda Nápoles. In 2009 three hippopotamuses were observed in a ciénaga near the town of Puerto Berrío (c. 75 km north of Hacienda Nápoles). In 2010 an individual was seen in the main channel of the Magdalena River near the town of Puerto Perales (17 km north). In 2013 three individuals were observed in the Cocorná River, a tributary of the Magdalena River, near the town of Estación Cocorná (16 km north). Hippopotamuses were also seen around that time in Ciénaga Barbacoas (105 km north). In 2014 a hippopotamus was again observed near Puerto Berrío. In 2017 two individuals were seen again near Estación Cocorná over at least 8 months. Although the majority of the hippopotamus population in Colombia is in the main pond near Hacienda Nápoles, these reports raise questions about the potential for this introduced species to spread across a wide swath of the Magdalena River basin.
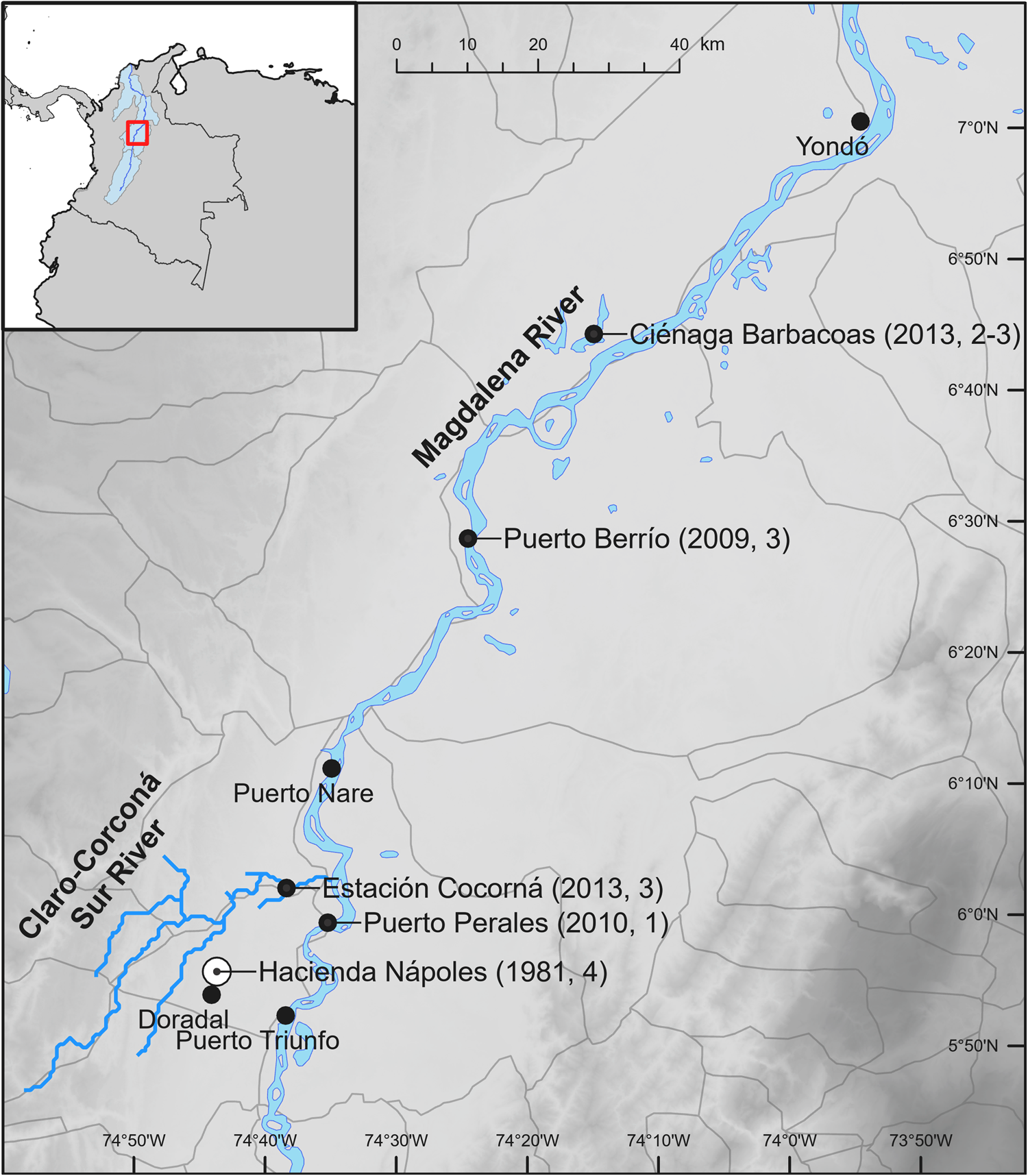
Fig. 2 The currently known range of the hippopotamus in the Magdalena River, with known localities, dates and initial numbers of hippopotamuses observed.
Across its native range, which extends throughout sub-Saharan Africa, the hippopotamus is categorized as Vulnerable on the IUCN Red List (Lewison & Pluhacek, Reference Lewison and Pluhacek2017). Populations declined by 7–20% from the mid 1990s to the early 2000s, and disappeared from some regions, although they may now have stabilized (Lewison & Pluhacek, Reference Lewison and Pluhacek2017). In regions where hippopotamuses have been protected, their populations have rebounded, in large part a result of life history characteristics that promote high population growth rates, particularly at low densities (Martin, Reference Martin2005; Kanga et al., Reference Kanga, Ogutu, Olff and Santema2011). Females can become reproductive at 3 years of age and can give birth to a calf every 2–3 years (Laws & Clough, Reference Laws, Clough and Rowlands1966; Pienaar et al., Reference Pienaar, Van Wyk and Fairall1966; Smuts & Whyte, Reference Smuts and Whyte1981). The species is long-lived (up to 40–50 years of age), reproductive for decades, and has low mortality, as adults have few predators other than people. Life history tables developed to model hippopotamus population growth upon reintroduction into Namibia showed that populations can grow at an annual rate of 5–11% (Martin, Reference Martin2005). In Namibia the population grew at 8%, similar to the rate observed in the Mara River region of Kenya following increased protection of the population since the 1950s (Kanga et al., Reference Kanga, Ogutu, Olff and Santema2011). Population growth rates can be particularly high at low densities, as recruitment tends to decline because of competition as populations approach carrying capacity (Chomba, Reference Chomba2013).
There are currently no census data available for the Colombian population, but there are observations from people working at Hacienda Nápoles regarding both life history characteristics and population estimates over time. Female hippopotamuses at Hacienda Nápoles become reproductive at 5–6 years of age and can have a calf every 18 months, and there has been little to no mortality among juveniles or adults. Using these traits, the life history models developed for the hippopotamus in Africa suggest the population could be growing at an annual rate of 7–8%, in line with population growth rates observed in other low-density hippopotamus populations. Comparing the predictions from this estimated growth rate to population estimates for Colombia at various times over the last 10 years, the data align well, suggesting these are reasonable initial estimates of population growth for this population (Fig. 3). Using the most recent population estimate (40–60 individuals; DEL, pers. obs.) and a constant rate of growth at 7–8% per year, we estimate there could be 400–800 hippopotamuses in the Magdalena River Basin by 2050. However, a growth rate of nearly 11% per year is not unrealistic, in which case there could be > 5,000 hippopotamuses by 2050.
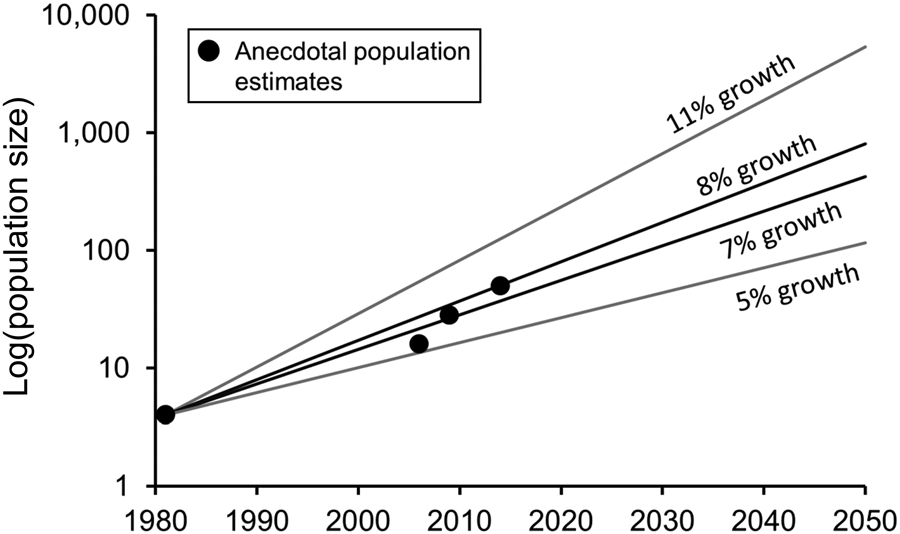
Fig. 3 Range of estimated population growth rates for the hippopotamus population in Colombia, based on an initial population of four individuals and population growth rates documented in low-density hippopotamus populations in Africa, and anecdotal population estimates (DEL, pers. obs.).
There are ongoing discussions about who is responsible for the management of the Colombian hippopotamus population. After Escobar's death, the Colombian Dirección Nacional de Estupefacientes (the national drug enforcement agency) took responsibility for his property, but because of a lack of technical expertise with large animals, responsibility for the hippopotamuses was transferred to the environmental authority CORNARE (Corporación Autónoma Regional de las Cuencas de los Ríos Negro y Nare), the provincial authority for management of environment and wildlife in the region encompassing Hacienda Nápoles. The management goal is to halt the growth of the hippopotamus population and relocate them from the wild. Regions in which hippopotamuses have been observed (including Puerto Berrío) also include areas under the jurisdiction of CORANTIOQUIA (Corporación Autónoma Regional del Centro de Antioquia), another regional environmental authority in the Magdalena River Basin.
There have been discussions about which strategies should be used to remove the hippopotamuses from the wild, but management interventions have not been straightforward. In 2009 three hippopotamuses living near Puerto Berrío were reportedly damaging crops and scaring fishers and cattle owners. The government brought in specialists to hunt and kill the hippopotamuses, helped by the Colombian military. A large male hippopotamus, known locally as Pepe, was killed, and a photograph of the team posing with the dead hippopotamus was published, prompting an outcry within Colombia. Animal rights activists took the government to court, and the subsequent ruling banned the killing of hippopotamuses to manage their population. In 2010 a group of 10 Colombian researchers wrote to the government to publicly denounce the ruling, arguing that the hippopotamuses were a large and potentially dangerous non-native species and should be killed before they could spread more widely. Nevertheless, the ruling remained, and surgical sterilization was adopted as the official management strategy. Since 2011 four hippopotamuses have been sterilized (Betancur et al., Reference Betancur, Giraldo and Suarez2016), but surgical sterilization is expensive and logistically challenging. Hippopotamuses are particularly difficult to tranquilize for capture because they have thick skin, are frequently found in or near water (in which a tranquilized individual could drown), and can be dangerous to people in close proximity (Dembitzer, Reference Dembitzer2017). The ability of dominant males to mate with multiple females also suggests sterilization will not be effective unless a high proportion of males are castrated. Several individuals have been translocated to zoos but there are a limited number of facilities with sufficient habitat and resources to support them.
Potential ecological effects on the Magdalena River Basin
The Magdalena River is the largest river system in Colombia, flowing 1,528 km and draining 262,075 km2 (24% of Colombia's area). The river begins at 3,685 m in the Andes Mountains and flows north-west through an extensive valley between the eastern and central Cordilleras (Restrepo et al., Reference Restrepo, Kjerfve, Hermelin and Restrepo2006). Downstream of the convergence of the Magdalena and Cauca Rivers, the main channel flows through an extensive floodplain (22,000 km2), the Momposina depression, and into the Caribbean Sea at Barranquilla. The Magdalena–Cauca has one of the highest sediment yields of the large South American rivers, transporting 184 Mt of sediment to the ocean annually (Restrepo & Escobar, Reference Restrepo and Escobar2018). The Magdalena–Cauca River basin contains 80% of Colombia's human population, and there are a number of anthropogenic activities influencing its conservation, including hydropower development, deforestation, urbanization, mining, and livestock and agricultural development (Galvis & Mojica, Reference Galvis and Mojica2007; Jiménez-Segura et al., Reference Jiménez-Segura, Galvis-Vergara, Cala-Cala, García-Alzate, López-Casas and Ríos-Pulgarín2016). There are currently 35 dams in the Magdalena–Cauca River basin, and plans for additional dams, although further hydropower development is likely to have substantial environmental costs (Jimenez-Segura et al., Reference Jimenez-Segura, Santamaria, López-Casas, Delgado, Valderrama and Gil2014; Angarita et al., Reference Angarita, Wickel, Sieber, Chavarro, Maldonado Ocampo and Herrera-R.2018).
The middle Magdalena River has a highly variable flow regime, with peak flows in April–June and October–November corresponding to the rainy seasons, and low flows in January–February and July–August corresponding to the dry seasons (Restrepo et al., Reference Restrepo, Kjerfve, Hermelin and Restrepo2006). The hydrological dynamism of this river helps maintain a complex riparian/floodplain habitat of inter-connected marshes, swamps and channels that connect to the main river channel and exert a regulatory effect on river discharge (Cormagdalena, 2007; Guhl, Reference Guhl2016). Seasonal changes in the river level result in dramatic changes in the connectivity of aquatic habitats and migration of freshwater fishes. Seasonally inundated floodplain lakes, known as ciénagas, provide nursery grounds for 63 native fish species, many of which are threatened (Jiménez-Segura et al., Reference Jiménez-Segura, Palacio and Leite2010, Reference Jiménez-Segura, Galvis-Vergara, Cala-Cala, García-Alzate, López-Casas and Ríos-Pulgarín2016). As river levels and connectivity decline, fish migrate into the main river channel in search of spawning grounds. This regular migration sustains the most important fishery in the country (Jiménez-Segura et al., Reference Jiménez-Segura, Galvis-Vergara, Cala-Cala, García-Alzate, López-Casas and Ríos-Pulgarín2016).
The middle Magdalena River basin, where the hippopotamuses have been observed, has a warm and humid climate, with a mean annual temperature of 28 °C and a mean annual rainfall of 2,050 mm. The natural landscape was historically tropical moist forest, c. 75% of which has been replaced by anthropogenic land use, including human settlements and agro-ecosystems such as palm plantations and livestock pasture (Cormagdalena, 2007, 2013). Nevertheless, the region remains important for floristic (Balcázar-Vargas et al., Reference Balcázar-Vargas, Rangel-Ch. and Linares-C.2000) and avian biodiversity (Hurd & Bohórquez, Reference Hurd and Bohórquez2000; Laverde-R. et al., Reference Laverde-R., Stiles and Múnera-R.2005), and is home to the Critically Endangered brown spider monkey Ateles hybridus, one of the most threatened Neotropical primates (Aldana et al., Reference Aldana, Beltrán, Torres-Neira and Stevenson2008; Link et al., Reference Link, Stevenson, de Luna, Schwitzer, Mittermeier, Rylands, Chiozza, Williamson, Wallis and Cotton2015a), and the Vulnerable Antillean manatee Trichechus manatus (Deutsch et al., Reference Deutsch, Self-Sullivan and Mignucci-Giannoni2008).
Hippopotamuses could have potentially significant ecological effects on the ecosystems of the middle Magdalena River, as they are capable of modifying their physical environment in a way that alters habitat and resource availability for a range of other species (Eltringham, Reference Eltringham1999; Moore, Reference Moore2006; Bakker et al., Reference Bakker, Pagès, Arthur and Alcoverro2016). Hippopotamuses wallow in aquatic environments, which can include rivers, lakes and wetlands, during the day and at night graze on the adjacent land. An individual can transport 750 kg dry mass per year of carbon and nutrients from terrestrial to aquatic ecosystems via defecation and excretion (Subalusky et al., Reference Subalusky, Dutton, Rosi-Marshall and Post2015). These resource subsidies can influence aquatic productivity in complex ways (Subalusky et al., Reference Subalusky, Dutton, Njoroge, Rosi and Post2018). Low to moderate levels of resource input may stimulate productivity and provide food resources for aquatic insects and fish (Masese et al., Reference Masese, Abrantes, Gettel, Bouillon, Irvine and McClain2015). High levels of input can decrease productivity in benthic environments (Dawson et al., Reference Dawson, Pillay, Roberts and Perissinotto2016; Stears et al., Reference Stears, McCauley, Finlay, Mpemba, Warrington and Mutayoba2018). These inputs may not be a significant factor at the whole-river scale in a river the size of the Magdalena, but the effects of such resource loading on aquatic ecosystems can be highly localized because of the tendency of hippopotamuses to aggregate in pools, leading to zones of anoxia and downstream effects (Wolanski & Gereta, Reference Wolanski and Gereta1999; Dutton et al., Reference Dutton, Subalusky, Hamilton, Rosi and Post2018). For example, aggregations in ciénagas along the Magdalena River could lead to eutrophic conditions and ultimately anoxic pools, which could be harmful to aquatic life, particularly during low water flow. As ciénagas are nursery grounds for a wide range of aquatic species, even localized zones of poor water quality could have negative effects on aquatic biodiversity and fisheries, particularly when high flows reconnect these anoxic pools to other components of the river system (Dutton et al., Reference Dutton, Subalusky, Hamilton, Rosi and Post2018).
Hippopotamus movements could have significant effects on the geomorphology and hydrology of the middle Magdalena River by wallowing, scouring and compressing the bottom of pools, increasing the duration for which they hold water (Naiman & Rogers, Reference Naiman and Rogers1997). When hippopotamuses move through densely vegetated aquatic habitats they create paths that may form or connect ponds, or that may fill with sediment in subsequent floods, altering hydrological connectivity and geomorphological heterogeneity (McCarthy et al., Reference McCarthy, Ellery and Bloem1998). These behaviours create habitats that have been shown to benefit fish in African ecosystems, such as the Okavango Delta, by creating deep pools that hold water for long time periods and by increasing hydrological connectivity into backswamp habitats (Mosepele et al., Reference Mosepele, Moyle, Merron, Purkey and Mosepele2009). Such behaviours could influence ciénagas along the middle Magdalena River in a similar way, increasing the duration of inundation and hydrological connectivity with the Magdalena River, which may help maintain open channels and pools that would otherwise be filled with sediment from land-use change. However, the life histories of aquatic plants and animals in this ecosystem are tightly linked to the ecosystem's hydroperiods, and any changes are likely to alter the composition of aquatic plant and animal communities in unpredictable ways. For example, the ecological importance of ciénagas and their macrophyte communities as nursery habitat for larval and juvenile fish partly depends upon their separation from the mainstem channel at low flows and associated protection from other, larger predators; this could be compromised by increased hydrological connectivity with the main river. These habitats also provide important feeding, resting and breeding sites for Antillean manatees (Mojica-Figueroa et al., Reference Mojica-Figueroa, Arévalo-González, González and Murillo2014), raising the possibility of intraspecific interactions between these two aquatic species. Hippopotamuses coexist with the African manatee Trichechus senegalensis in West Africa, but we are not aware of any research that has examined interactions between them.
The movement of hippopotamuses between aquatic and terrestrial habitats creates paths on riverbanks and increases connectivity between riparian and aquatic ecosystems (Naiman & Rogers, Reference Naiman and Rogers1997), which may become a conduit for terrestrial nutrients and sediment into the aquatic ecosystem from overland flows (Jacobs et al., Reference Jacobs, Bechtold, Biggs, Grimm, Lorentz and McClain2007), and may also allow expansion of wetlands into the surrounding terrestrial ecosystem (McCarthy et al., Reference McCarthy, Ellery and Bloem1998). Such movements could expand ciénagas in the middle Magdalena into surrounding habitats utilized by people and livestock, and could influence riparian vegetation by destabilizing riverbanks and altering the hydrological regime.
In terrestrial systems, hippopotamuses alter biodiversity through grazing patterns. Their mouth morphology enables them to graze close to the ground, cropping grass and forbs at their base (Eltringham, Reference Eltringham1999). This creates grazing lawns, which influence the species composition of the vegetation and that of ungulates that graze there (Lock, Reference Lock1972; Kanga et al., Reference Kanga, Ogutu, Piepho and Olff2013). There has been a large degree of conversion from native landcover to agroecosystems in the Magdalena River basin, so it is unlikely hippopotamuses will have a significant effect on the native terrestrial vegetation. However, their focal zones of activity along riparian corridors are important for faunal diversity (Link et al., Reference Link, Valencia, Céspedes, Duque, Cadena and Di Fiore2015b; Sandoval-H et al., Reference Sandoval-H, Gómez and Cadena2016), which could lead to effects in areas that have escaped conversion. Hippopotamuses also supplement their grazing with aquatic macrophytes when available (Grey & Harper, Reference Grey and Harper2002), which could negatively impact macrophyte communities in the ciénagas that provide fish nursery habitat.
Potential socio-economic effects in the Magdalena River basin
The Magdalena River is vital for transport and commerce for a large swath of Colombia. It produces 95% of the total thermal energy and 70% of the hydroelectric power generated in the country (Cormagdalena, 2007; Jimenez-Segura et al., Reference Jimenez-Segura, Santamaria, López-Casas, Delgado, Valderrama and Gil2014). The middle Magdalena was historically important for commercial navigation, but sediment accumulation has reduced the main channel's depth and prevented the navigation of large ships (Cormagdalena, 2013). The majority of artisanal fishing in Colombia is located in the Magdalena basin, with 8,000 artisanal fishers in the middle Magdalena (Cormagdalena, 2007), but the river's fishery has declined since 1970 because of unsustainable fishing practices, construction of reservoirs, and changes in land use resulting in anthropogenic contamination (Cormagdalena, 2007; Galvis & Mojica, Reference Galvis and Mojica2007; Jiménez-Segura et al., Reference Jiménez-Segura, Galvis-Vergara, Cala-Cala, García-Alzate, López-Casas and Ríos-Pulgarín2016). Increasing extraction of sand and rocks from the river bed for infrastructure construction is a new challenge for river conservation.
A large proportion of socio-economic development and human livelihoods in the middle Magdalena River basin depends directly upon the river and its waterways (Galvis & Mojica, Reference Galvis and Mojica2007), and the further expansion of the hippopotamus into this ecosystem is likely to affect these livelihoods. Hippopotamuses are territorial and aggressive, and could pose threats to fishermen and cascajeros (people who extract sand and rocks from the riverbed), who frequently use the side channels and shallow waterways likely to be favoured by hippopotamuses. Some local people have already changed their behaviour to avoid encounters with hippopotamuses (LJS, pers. obs.).
The species’ nocturnal terrestrial foraging, which is typically over 1–2 km but can be up to 5–10 km inland (Eltringham, Reference Eltringham1999), could also have socio-economic effects. In Africa conflict with the species arises from their destruction of agricultural crops, particularly during droughts (Post, Reference Post2000; Kanga et al., Reference Kanga, Ogutu, Piepho and Olff2012). Crops cultivated within 5–10 km of the middle Magdalena River and its ciénagas could be at risk, and this would be challenging for small-scale farmers. Hippopotamuses also pose risks to human safety, particularly when they are on land. Across sub-Saharan Africa numerous deaths are attributed annually to the species (Kanga et al., Reference Kanga, Ogutu, Piepho and Olff2012). Any communities living within 10 km of the current range of the species in Colombia (Fig. 2) need to be educated about the risks and how to avoid them.
At its current density, the hippopotamus may be perceived positively by some people living in the region. Hippopotamuses around Hacienda Nápoles have become a tourist attraction, advertized by the water and theme park and celebrated with statues and signs by local businesses in Doradal, seemingly to appeal to tourists. Residents have become accustomed to seeing hippopotamuses walking within the city limits in the evenings. Further research is needed to adequately characterize people's perceptions of the species in Colombia and their assessment of the costs and benefits associated with the introduction.
Discussion
The middle Magdalena River basin is in the nascent stages of the introduction of a non-native megaherbivore, but little is as yet known beyond anecdotal information about the current status of the population and its potential ecological and social effects (Dembitzer, Reference Dembitzer2017). Research is required to document this novel occurrence, to contextualize it within the current scientific understanding of introduced species and human–wildlife interactions, and to inform resource managers and local stakeholders confronting the realities of this introduction. We offer three recommendations for further study.
Firstly, we need a better understanding of the current and predicted future population size. The estimates we have provided, which span an order of magnitude, are based upon anecdotal accounts of the current population size. A comprehensive survey is required, to refine this estimate. We also need more information on life history characteristics of this population (age at maturity, fecundity and mortality), to improve population growth estimates, and on the potential fitness costs of inbreeding depression, given the small founding population of four individuals, with only one male (Huisman et al., Reference Huisman, Kruuk, Ellis, Clutton-Brock and Pemberton2016). These data would facilitate predictions of population growth under different management scenarios and help resource managers determine the best response to this introduction.
Secondly, coincident with the population survey, the current range of the species needs to be confirmed. The characteristics of potential habitat for hippopotamuses could be used to develop a habitat suitability map for prediction of the geographical spread of the species. Together, these tools could allow researchers and environmental authorities to determine the likelihood of the potential expansion of the hippopotamus into other regions and countries.
Thirdly, a study is needed of the ecological and socioeconomic effects that the species is already having and may have in the future. This introduction provides a unique opportunity to examine how hippopotamuses may influence ecological functions through provision of resource subsidies to aquatic ecosystems, alteration of terrestrial and aquatic vegetation communities through grazing and disturbance, and influence on riparian and hydrological connectivity. We also need to identify any native flora and fauna that could be sensitive to the effects of the species' presence, to understand potential effects on humans and their livelihoods, and to predict and mitigate negative interactions between humans and hippopotamuses. The latter will require an understanding of people's perceptions of risk and engagement in participatory planning for any interventions (Treves et al., Reference Treves, Wallace, Naughton-Treves and Morales2006).
The introduction of novel species can have major ramifications for native ecosystems and the human populations that depend upon these ecosystems. There are already a number of non-native mammal species in Colombia, several of which are considered invasive (Ramírez-Chaves et al., Reference Ramírez-Chaves, Ortega-Rincón, Pérez and Marín2011). These species can impact native biodiversity directly and managing them draws limited funding away from conservation of native species. In South America the introduced beaver Castor canadensis, for example, has altered large areas of native forest and led to changes in species composition and ecosystem function (Anderson et al., Reference Anderson, Griffith, Rosemond, Rozzi and Dollenz2006; Anderson & Rosemond, Reference Anderson and Rosemond2007); the introduction of the hippopotamus could have similar effects on native habitats in Colombia.
Alternatively, in a time of global defaunation, the establishment of a new megaherbivore population could have benefits (Dirzo et al., Reference Dirzo, Young, Galetti, Ceballos, Isaac and Collen2014; Lundgren et al., Reference Lundgren, Ramp, Ripple and Wallach2018). The introduction of the hippopotamus in Colombia may be a form of accidental rewilding, functionally replacing historically native megafaunal species that went extinct > 10,000 years ago at the end of the Pleistocene (Dembitzer, Reference Dembitzer2017; Lundgren et al., Reference Lundgren, Ramp, Ripple and Wallach2018). It has been suggested there is potential for rewilding in South America because of the absence of megafauna, and megaherbivores in particular, compared to both prehistorical and historical baselines (Svenning & Faurby, Reference Svenning and Faurby2017), suggesting the hippopotamus could provide ecological functions that were lost with the extinction of taxonomically different but functionally similar species. In addition, the establishment of a robust hippopotamus population in South America could have conservation benefits, as hippopotamus populations in their native range of sub-Saharan Africa have, until recently, been declining (Lewison & Oliver, Reference Lewison and Oliver2008; Lewison & Pluhacek, Reference Lewison and Pluhacek2017). The potential costs and benefits of this introduction must be carefully weighed to determine the most appropriate response.
The eradication of an introduced species after it has become established can be difficult and expensive, and it is imperative to decide early in the introduction how the species will be handled, and then to properly fund the desired approach (Puth & Post, Reference Puth and Post2005). The current policy is to halt the growth of the population, and ultimately relocate them from the wild, but not by culling. It is important to predict the future distribution of hippopotamuses in Colombia in response to current management approaches and the likely social and ecological effects across this range to inform the decision-making process sufficiently (Ricciardi et al., Reference Ricciardi, Hoopes, Marchetti and Lockwood2013; Jeschke et al., Reference Jeschke, Bacher, Blackburn, Dick, Essl and Evans2014). Consideration also must be taken of local perceptions of hippopotamuses and proposed methods of controlling their spread, to avoid conflict over proposed management actions (García-Llorente et al., Reference García-Llorente, Martín-López, González, Alcorlo and Montes2008; Crowley et al., Reference Crowley, Hinchliffe and McDonald2017). A complex confluence of history, culture and environment in Colombia have set the stage for this novel, and unintended, ecological introduction of hippopotamuses into the Magdalena River basin. Research and information sharing will be critical for management authorities as they navigate an array of potential social, ecological and financial concerns in their response to the introduction.
Acknowledgements
Funding was provided by a grant from the National Geographic Society to EPA (W447-16) and by grants from the US National Science Foundation to DMP (DEB 1354053, 1753727). This is contribution no. 929 from the Southeast Environmental Research Center in the Institute of Water & Environment at Florida International University. We thank colleagues in Colombia for sharing valuable information about introduced hippopotamuses, especially historical accounts, and A. Farah and G. Herrera for creating the map.
Author contributions
Compilation of historical information and literature review: all authors; development of population estimate: AS, DP; writing: all authors.
Conflicts of interest
None.
Ethical standards
This research abided by the Oryx guidelines on ethical standards.





 Open access
Open access