



Introduction
The Spoon-billed Sandpiper Calidris pygmaea is a Critically Endangered calidrid found in Asia (BirdLife International 2017). The species breeds in north-east Siberia, migrates through the East Asian-Australasian flyway with stopover sites in the Yellow Sea, and is thought to winter principally in South China, Thailand, Myanmar, and Bangladesh (Chowdhury Reference Chowdhury2012, Lappo et al. Reference Lappo, Tomkovich and Syroechkovkiy2012, Zöckler et al. Reference Zöckler, Beresford, Bunting, Chowdhury, Clark, Fu, Htin Hla, Morozov, Syroechkovskiy, Kashiwagi, Lappo, Tong, Long, Yu, Huettmann, Akasofu, Tomida and Buchanan2016). The estimated global population of Spoon-billed Sandpiper fell from around 2,000 pairs in the 1970s to under 200 in 2014, with a rate of decline up to 26% per annum recorded during the 2000s (Flint and Kondratiev Reference Flint, Kondratiev and Voinstvenski1977, Tomkovich et al. Reference Tomkovich, Syroechkovskiy, Lappo and Zöckler2002, Zöckler et al. Reference Zöckler, Syroechkovskiy and Atkinson2010a, Clark et al. Reference Clark, Anderson, Li, Syroechkovskiy, Tomkovich, Zöckler, Lee and Green2018).
Demographic studies indicate an unusually low per capita recruitment of two-year old adults to the breeding population, while other demographic rates are similar to those of other small calidrids (Zöckler et al. Reference Zöckler, Syroechkovskiy and Atkinson2010a). This finding suggests that the major external drivers of the population decline are factors affecting the mortality rate of immature birds, such as loss of intertidal habitat at migration stopover sites and hunting on the wintering grounds (Zöckler et al. Reference Zöckler, Hla, Clark, Syroechkovskiy, Yakushev, Daengphayon and Robinson2010b, Reference Zöckler, Beresford, Bunting, Chowdhury, Clark, Fu, Htin Hla, Morozov, Syroechkovskiy, Kashiwagi, Lappo, Tong, Long, Yu, Huettmann, Akasofu, Tomida and Buchanan2016, Chowdhury Reference Chowdhury2012, Tong et al. Reference Tong, Zhang, Jing, Zöckler and Clark2012, Piersma et al. Reference Piersma, Lok, Chen, Hassell, Yang, Boyle, Slaymaker, Chan, Melville, Zhang and Ma2016, Peng et al. Reference Peng, Anderson, Chang, Choi, Chowdhury, Clark, Gan, Hearn, Li, Lappo, Liu, Ma, Melville, Phillips, Syroechkovskiy, Tong, Wang and Zöckler2017, Choi et al. Reference Choi, Jackson, Gallo-Cajiao, Murray, Clemens, Gan and Fuller2018). Spoon-billed Sandpipers are caught as bycatch during mist-netting of larger species of shorebirds at several sites in Myanmar, Bangladesh and China (Bird et al. Reference Bird, Lees, Chowdhury, Martin and Haque2010, Chowdhury Reference Chowdhury2010, Zöckler et al. Reference Zöckler, Hla, Clark, Syroechkovskiy, Yakushev, Daengphayon and Robinson2010b, Martinez and Lewthwaite Reference Martinez and Lewthwaite2013, Pyae-Phyo et al. Reference Pyae-Phyo, Moses, Clark, Anderson, Hilton, Buchanan, Zöckler and Green2018). There have already been some successful conservation interventions, discouraging hunting at known sites (Bird et al. Reference Bird, Lees, Chowdhury, Martin and Haque2010, Clark et al. Reference Clark, Pain and Green2014, Zöckler et al. Reference Zöckler, Beresford, Bunting, Chowdhury, Clark, Fu, Htin Hla, Morozov, Syroechkovskiy, Kashiwagi, Lappo, Tong, Long, Yu, Huettmann, Akasofu, Tomida and Buchanan2016).
Spoon-billed Sandpipers utilise estuarine mudflats at migration stopovers and on the wintering grounds (Tong et al. Reference Tong, Zhang, Jing, Zöckler and Clark2012). In recent decades, large expanses of these mudflats have been lost to land claim and development (Melville et al. Reference Melville, Chen and Ma2016, Peng et al. Reference Peng, Anderson, Chang, Choi, Chowdhury, Clark, Gan, Hearn, Li, Lappo, Liu, Ma, Melville, Phillips, Syroechkovskiy, Tong, Wang and Zöckler2017, Studds et al. Reference Studds, Kendall, Murray, Wilson, Rogers, Clemens, Gosbell, Hassell, Jessop, Melville, Milton, Minton, Possingham, Riegen, Straw, Woehler and Fuller2017), with loss in the East Asian-Australasian flyway estimated to occur at 1.66% per annum (Murray and Fuller Reference Murray and Fuller2015). Remaining intertidal habitat at stopover sites is further threatened by the encroachment of the invasive grass Spartina alterniflora, which traps sediment accelerating conversion of mudflats to dry land (Peng et al. Reference Peng, Anderson, Chang, Choi, Chowdhury, Clark, Gan, Hearn, Li, Lappo, Liu, Ma, Melville, Phillips, Syroechkovskiy, Tong, Wang and Zöckler2017). Areas of intertidal mudflat have an inherently patchy distribution along the coast and losses restrict the stopover sites available for migrating birds, likely increasing energetic demands, limiting food supplies and rendering the birds more vulnerable to stochastic events such as storms (Sutherland et al. Reference Sutherland, Alves, Amano, Chang, Nicholas, Finlayson, Gill, Gill, González, Gunnarsson, Kleijn, Spray and Székely2012, Murray et al. Reference Murray, Clemens, Phinn, Possingham and Fuller2014, Studds et al. Reference Studds, Kendall, Murray, Wilson, Rogers, Clemens, Gosbell, Hassell, Jessop, Melville, Milton, Minton, Possingham, Riegen, Straw, Woehler and Fuller2017, Wang et al. Reference Wang, Xiao, Zou, Chen, Ma, Dong, Doughty, Zhong, Qin, Dai, Li, Zhao and Li2020).
Surveys of individually marked birds at stopover sites indicates that up to 50% of the global population has not been located at wintering sites using traditional field survey techniques (Zöckler et al. Reference Zöckler, Beresford, Bunting, Chowdhury, Clark, Fu, Htin Hla, Morozov, Syroechkovskiy, Kashiwagi, Lappo, Tong, Long, Yu, Huettmann, Akasofu, Tomida and Buchanan2016, Clark et al. Reference Clark, Anderson, Li, Syroechkovskiy, Tomkovich, Zöckler, Lee and Green2018). Satellite tracking studies have located previously unknown migration stopover and wintering locations (Chang and Clark Reference Chang and Clark2018), but tags are expensive and only a small number of birds can be tracked, which means that not all wintering sites will be detected by this method. Hence, it is important to develop other approaches to identify additional staging and wintering locations to identify where protected areas are needed and target conservation interventions.
Species distribution modelling is an established tool in conservation and is used to identify potentially important - but hitherto unconfirmed - areas utilised by threatened species (Franklin Reference Franklin2010). It has previously been used to model the core wintering area for Spoon-billed Sandpipers, and successfully confirmed important known wintering sites, namely the Gulf of Mottama and the Inner Gulf of Thailand (Zöckler et al. Reference Zöckler, Beresford, Bunting, Chowdhury, Clark, Fu, Htin Hla, Morozov, Syroechkovskiy, Kashiwagi, Lappo, Tong, Long, Yu, Huettmann, Akasofu, Tomida and Buchanan2016). The predicted winter distribution also highlighted a previously unknown site in the Ganges-Brahmaputra Delta (Zöckler et al. Reference Zöckler, Beresford, Bunting, Chowdhury, Clark, Fu, Htin Hla, Morozov, Syroechkovskiy, Kashiwagi, Lappo, Tong, Long, Yu, Huettmann, Akasofu, Tomida and Buchanan2016), a location since proven to support a substantial wintering population (Chowdhury et al. Reference Chowdhury, Foysal, Diyan and Ahmed2018). However, the geographic scope of Zöckler et al. (Reference Zöckler, Beresford, Bunting, Chowdhury, Clark, Fu, Htin Hla, Morozov, Syroechkovskiy, Kashiwagi, Lappo, Tong, Long, Yu, Huettmann, Akasofu, Tomida and Buchanan2016) was limited. New sites in India, Sri Lanka, and Indonesia have recently been found to support Spoon-billed Sandpipers, indicating potentially suitable sites remain to be discovered in the wintering range. Additionally, Zöckler et al. (Reference Zöckler, Beresford, Bunting, Chowdhury, Clark, Fu, Htin Hla, Morozov, Syroechkovskiy, Kashiwagi, Lappo, Tong, Long, Yu, Huettmann, Akasofu, Tomida and Buchanan2016) focused on only part of the species’ non-breeding distribution and did not examine the potential distribution of stopover sites between south-east Asia and Arctic Russia.
Since Zöckler et al.’s (Reference Zöckler, Beresford, Bunting, Chowdhury, Clark, Fu, Htin Hla, Morozov, Syroechkovskiy, Kashiwagi, Lappo, Tong, Long, Yu, Huettmann, Akasofu, Tomida and Buchanan2016) analysis, many more Spoon-billed Sandpiper records have been gathered from a much wider area. A combination of field surveys at wintering and stopover sites, and a small number of satellite-tagged birds, yielded 2,798 new observations between 2015 and 2017. Here we produce species distribution models to identify the areas that might be suitable for Spoon-billed Sandpipers along the entire migratory route and a wider potential wintering distribution area than examined by Zöckler et al. (Reference Zöckler, Beresford, Bunting, Chowdhury, Clark, Fu, Htin Hla, Morozov, Syroechkovskiy, Kashiwagi, Lappo, Tong, Long, Yu, Huettmann, Akasofu, Tomida and Buchanan2016). We then compare these predictions with protected area coverage. In 2008, 17% of sites known to be occupied by non-breeding Spoon-billed Sandpiper were covered by protected area designations (Zöckler et al. Reference Zöckler, Syroechkovskiy and Bunting2008). However, the number of designated sites in the flyway has increased substantially in recent years (UNEP-WCMC, IUCN and NGS 2018), requiring a reassessment of how much Spoon-billed Sandpiper habitat is protected. Findings from the models will both inform future survey efforts and help to target conservation interventions.
Methods
Study regions
The passage and wintering distribution of Spoon-billed Sandpiper is extensive, spanning Arctic Russia to tropical south-east Asia. Consequently, conditions change markedly across their migration and wintering areas. Thus, we divided the study area into three regions (Figure 1). The south region (between 6ºS and 30ºN latitude) is where the birds predominantly winter, between early November and late February. This was bounded between 76º and 130ºE (i.e. southern tip of India to eastern Indonesia, excluding the island of New Guinea), these longitudinal and southern limits extend beyond all historical records for the species. The central region (30º–40º N) covers the core stopover sites. These sites are used during spring (northwards) migration between early March and mid-June, and then again during the post-breeding (southwards) migration between mid-August and late October. The north region (40º–63º N) is visited by migrating birds immediately before and after the breeding season, birds are present between mid-June and mid-August. Immature birds sometimes spend their first summer on wintering grounds, before returning to the north to breed as second-year birds (Zöckler et al. Reference Zöckler, Syroechkovskiy and Atkinson2010a).
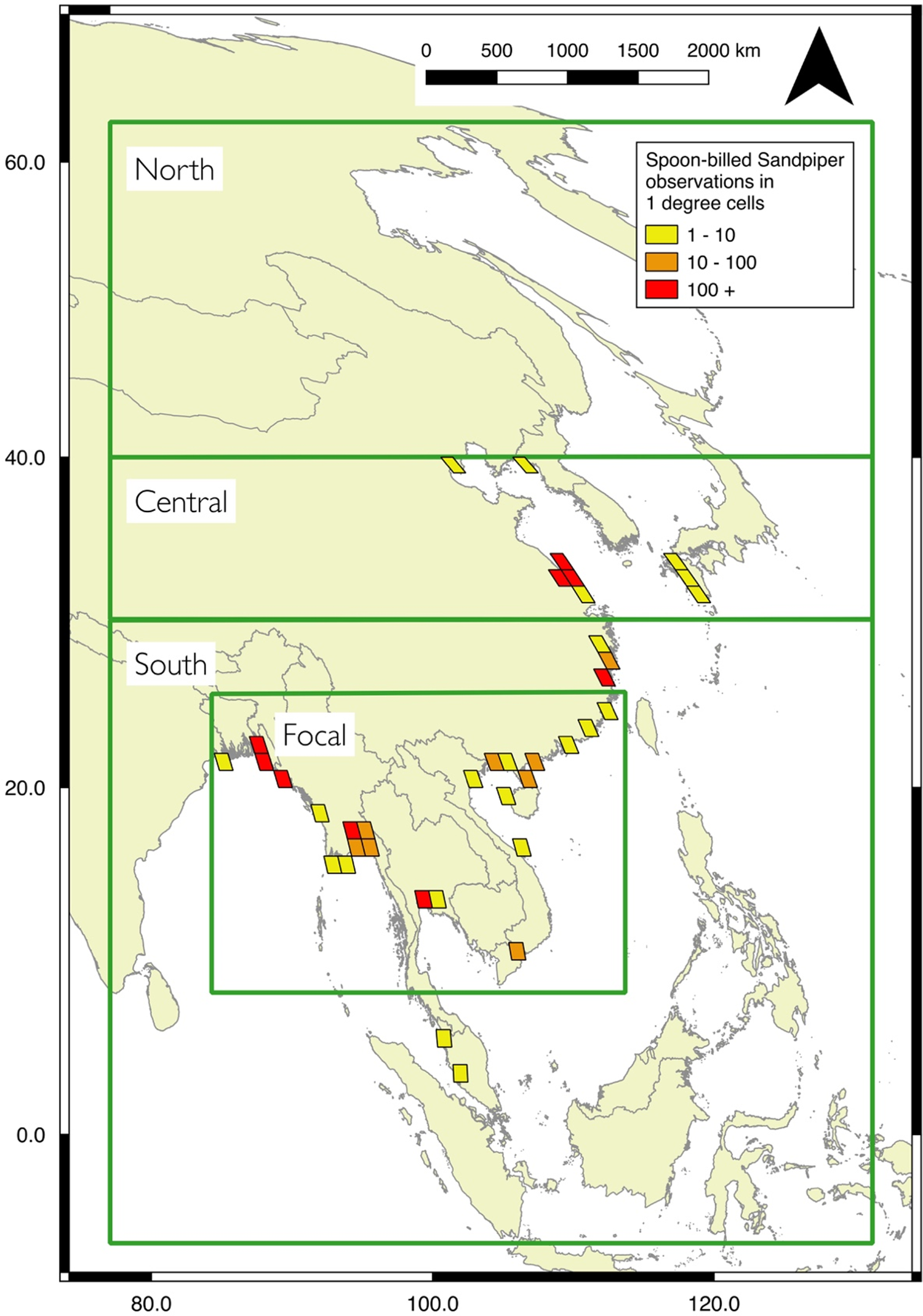
Figure 1. The potential non-breeding range of spoon-billed sandpiper, showing the three main regions used in this study: north, central and south. For the south region, the distribution model was initially built with the focal region, before extrapolation to the whole area. Number of Spoon-billed Sandpiper records from field surveys 2008–2017 in the central and south regions are shown in 1 degree squares, records from field surveys in the north region were not available.
Spoon-billed Sandpiper records
Location records came from two data sources: field observations and satellite tracking. Field observations of Spoon-billed Sandpipers made by experienced surveyors were conducted at passage and wintering sites located in the south and central regions between 2008 and 2017 (Figure 1). Field surveys have been conducted in the north region, but records were not available for modelling. Sites were selected for surveys either because they were known to be occupied from earlier records, or because occupation was considered possible based upon the presence of extensive tidal mud flats (Zöckler et al. Reference Zöckler, Beresford, Bunting, Chowdhury, Clark, Fu, Htin Hla, Morozov, Syroechkovskiy, Kashiwagi, Lappo, Tong, Long, Yu, Huettmann, Akasofu, Tomida and Buchanan2016). Given the highly dynamic nature of mudflats and the habitat losses that have occurred in some areas, we limited data to recent records (i.e. 2008–2017) as site suitability may have changed over a longer period. We sought to produce a distribution model that would inform future monitoring and protection rather than map historic use. There were 5,148 field observations of Spoon-billed Sandpipers from 544 pixels of 500 x 500 m (0.25 km2; see below).
Nine birds were fitted with satellite tags (Microwave Telemetry Inc, Maryland, USA) at various stages in the annual cycle: three in autumn 2016, two in spring 2017, three in summer 2017 and one in autumn 2017 (Chang and Clark Reference Clark, Anderson, Li, Syroechkovskiy, Tomkovich, Zöckler, Lee and Green2018). Each bird provided data for a different part of the year, but in combination cover the entire migratory flyway. These tracking data provided additional records for known sites, and pinpointed locations used by Spoon-billed Sandpipers where field surveys were not conducted. Fix accuracy is classified at moment of capture, and for this study we only used fixes with a location error of less than 1,500 m. This provided 1,107 fixes from a total of 477 0.25 km2 pixels, so that the total dataset used for the modelling contained 1,021 occupied 0.25 km2 pixels.
Satellite imagery of study regions
We focused on coastal areas in the study regions and used country polygons from gadm.org (version 3.6) as the basis for the coastline. Given Spoon-billed Sandpiper preference for dynamic areas of coastline, we visually compared the gadm.org coastline against 2017 Sentinel 2 satellite remote sensing data with a spatial resolution of 10 m and modified the coastline where appropriate (ESA 2018). Of the 6,492 Spoon-billed Sandpiper records available from surveys and satellite tags, 85% were from the seaward side of the coastline, with the remainder inland. We therefore restricted the study focus to 5 km onshore and 30 km offshore of the coastline, this area included 96% of the total number of records (i.e. 6,255 records, comprised the 5,148 observations and 1,107 tag fixes described above). The much wider area of sea was included because Zöckler et al. (Reference Zöckler, Beresford, Bunting, Chowdhury, Clark, Fu, Htin Hla, Morozov, Syroechkovskiy, Kashiwagi, Lappo, Tong, Long, Yu, Huettmann, Akasofu, Tomida and Buchanan2016) found that offshore conditions were important predictors of Spoon-billed Sandpiper presence, particularly ocean chlorophyll, which is potentially related to inshore conditions associated with photosynthetic activity in estuaries and tidal mudflats.
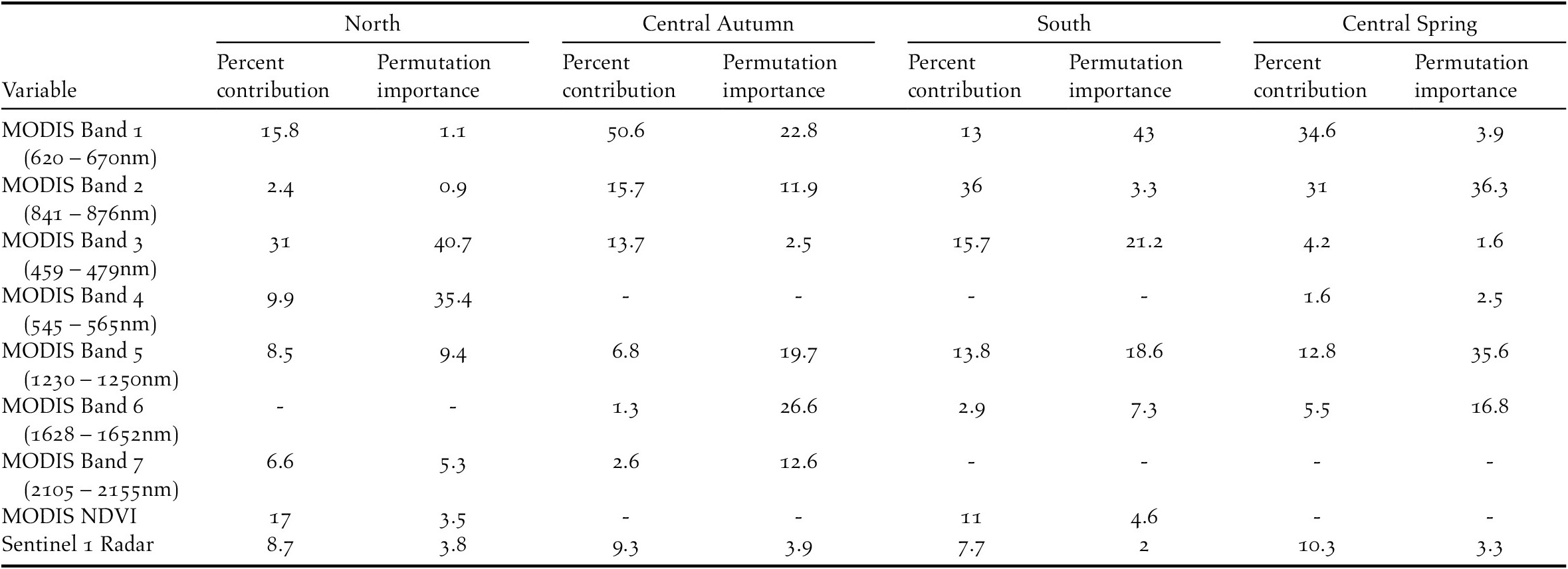
We utilised Google Earth Engine (https://earthengine.google.com) to access and download the satellite imagery for the buffered coastline in each of the three regions. For the distribution models, we selected Sentinel 1 synthetic aperture radar data (ESA 2018), and 8-day composite surface reflectance scenes from MODIS (Vermote Reference Vermote2015). Radar was included as a proxy for separating mudflat characteristics (van der Wal et al. Reference van der Wal, Herman and Wielemaker-van den Dool2005) and was resampled to 0.25 km2 to match the resolution of the MODIS imagery. The MODIS product includes seven bands that span wavelengths of 459 to 2,155nm (Table 1). We restricted imagery to the twelve months between November 2016 and October 2017. Imagery for each region was limited to the period in which Spoon-billed Sandpipers are generally present: south, 01/11/2016 – 28/02/2017, central Spring, 01/03/2017 – 14/06/2017, north, 15/06/2017 – 15/08/2017, and, central Autumn, 16/08/2017 – 31/10/2017. Some adult birds spend the entire winter in the central region, while some first-year birds remain in the central region rather than migrating north in the breeding season (Chang and Clark Reference Chang and Clark2018). Spoon-billed Sandpiper may therefore be present throughout the year in the central region, but we focused on potential distribution during the stopover periods as this is when most birds are present. We selected the mean pixel values from the time period for each region. We used Sentinel 1 radar, all seven MODIS bands, and MODIS-derived Normalised Difference Vegetation Index (NDVI) to give a total of nine variables for the modelling.
Table 1. Importance of each variable in the four regional models, “-” indicates variable dropped during model construction. “Percent contribution” shows the relative contribution of each variable when it is included in the final model, “Permutation importance” shows the percentage fall in training AUC when values for that variable are randomly permuted while other variables are left unchanged.
To determine whether sites identified by the model are recognised as being important for wildlife, we compared model outputs with coverage of Key Biodiversity Areas (KBAs) and Important Bird and Biodiversity Areas (IBAs; BirdLife International 2018). Finally, to assess the level of habitat protection currently covering sites identified by the models, we used data from the World Database on Protected Areas (UNEP-WCMC and IUCN 2018) to overlay protected area boundaries onto our predicted model outputs. We included all designations except UNESCO Man and Biosphere Reserves.
Species distribution modelling
We used Maximum Entropy to model the species’ distribution with MaxEnt (version 3.4.1, Phillips et al. Reference Phillips, Dudík and Schapire2018). Pixels with a Spoon-billed Sandpiper record were classed as presences, and 10,000 background points sampled as pseudo-absences. We constructed four models in total, relating to the north, central (Autumn), central (Spring) and south regions (Figure 1). Initial models included all nine variables and were refined by stepwise backwards elimination, following each model run, the variable with the lowest relative contribution was dropped until all variables contributed >1% to the model. Where appropriate, the regularisation multiplier was increased to avoid over-fitting the model to the training data and produce smooth response curves (Phillips et al. Reference Phillips, Anderson and Schapire2006). Once a final model for each region was constructed, 10-fold cross-validation resampling was used to assess variable importance (Elith et al. Reference Elith, Phillips, Hastie, Dudík, Chee and Yates2011). MaxEnt does not segregate data spatially (Elith et al. Reference Elith, Phillips, Hastie, Dudík, Chee and Yates2011), potentially inflating estimates of model accuracy (Bladon et al. Reference Bladon, Donald, Jones, Collar, Deng, Dadacha, Abebe and Green2018). We attempted to minimise inflating model accuracy assessments by using only one record for each occupied 0.25 km2 cell in the models. Model fit was assessed using the AUC (Area Under the receiver-operator Curve) statistic, where a value of 0.5 implies the model is no better than random, while that of 0.9 and above indicates a good fit (Swets Reference Swets1988).
For the north and central regions, models were constructed for the entire area. For the south region, the model was initially built using a focal area encompassing the principal known wintering sites, corresponding broadly to the area used in Zöckler et al. (Reference Zöckler, Beresford, Bunting, Chowdhury, Clark, Fu, Htin Hla, Morozov, Syroechkovskiy, Kashiwagi, Lappo, Tong, Long, Yu, Huettmann, Akasofu, Tomida and Buchanan2016). This focal model was produced as described above and extrapolated to cover the full south region (Figure 1). Following Zöckler et al. (Reference Zöckler, Beresford, Bunting, Chowdhury, Clark, Fu, Htin Hla, Morozov, Syroechkovskiy, Kashiwagi, Lappo, Tong, Long, Yu, Huettmann, Akasofu, Tomida and Buchanan2016), we defined key potential sites as the 5% of pixels with the highest modelled probability of occupancy.
Results
Field observations of Spoon-billed Sandpiper came from coastal sites across the central and south regions (Figure 1). Distribution was patchy, with the majority of records from just a few sites in the wintering areas: the Meghna estuary and Cox’s Bazar in Bangladesh, Nan Thar island and the Gulf of Mottama in Myanmar, the Inner Gulf of Thailand, and Hainan and the Leizhou Peninsula in Guangdong, China. The most important stopover site was the southern Jiangsu coast in China. To an extent this skewed distribution reflects concentrated survey effort at known sites, however the species’ association with extensive tidal mudflats means that larger congregations are most probable in these locations.
The high AUC values signified that all four models were adequate descriptions of Spoon-billed Sandpiper distributions (Table 2). This indicates that the predicted outputs could be accurate representations of potential Spoon-billed Sandpiper distributions. Considering individual variables, MODIS bands 1, 2 and 3 (corresponding to visible red, infrared, and blue wavelengths respectively; Table 1) were generally the most important in all models, although the importance of each changed among regions (Table 1). Band 1 was the most important for central region models, particularly for the Autumn model. Bands 2 and 5 were notably important for the south and central Spring models, whereas band 3 was the most important for the north model. Radar was retained in all models, contributing between 7.7 and 10.3%. NDVI was included in models for north and south regions, but dropped from the central region models. MODIS bands 4, 6 and 7 were of low importance across all regions. Response curves and standard deviations of the variables for each model are given in Figure S1 in the online supplementary material.
Table 2. Summary of the AUC values from the 10-fold cross-validation analyses for each region model.

Model outputs for each region are shown in Figures 2 to 4, with the key sites labelled. For display purposes the 0.25 km2 pixels have been resampled to 10 km2 (full resolution versions are available in Figures S2 to S5). Potential sites with a high likelihood of occupancy for Spoon-billed Sandpipers are spread across the north region: near Shelikhova Bay (A), Karaginskiy and Oliutorskiy Bays (B), in Kamchatka, the western coast (C) and the mouth of the Kamchatka river (D), Turgurski and Academy Bays (E), at the mouth of the Amur liman and around northern Sakhalin (F), In the central region, there is a larger area predicted as suitable for Spoon-billed Sandpipers during the Autumn (southward) migration than Spring (northward) migration. However, both autumn and spring models identify three main areas in the Yellow Sea as particularly valuable habitat: Bohai and Laizhou Bays (G), the Jiangsu coast (H), and the Yangtze Delta and Hangzhou Bay (J), either side of Shanghai. The focal area of the south region identifies the key overwintering sites known to support Spoon-billed Sandpiper, namely the Ganges-Brahmaputra Delta (M), the Rakhine coast and Nan Thar island (N), Hainan and the western Guangdong coast (P), the Gulf of Mottama and Ayeyarwady Delta (Q), the Inner Gulf of Thailand (R), and the Mekong Delta (S). The model for the wider south region also highlights sites where Spoon-billed Sandpipers were seen in 2018: Mannar in Sri Lanka (K), Fraserganj in India (L), and Aceh in Indonesia (T).
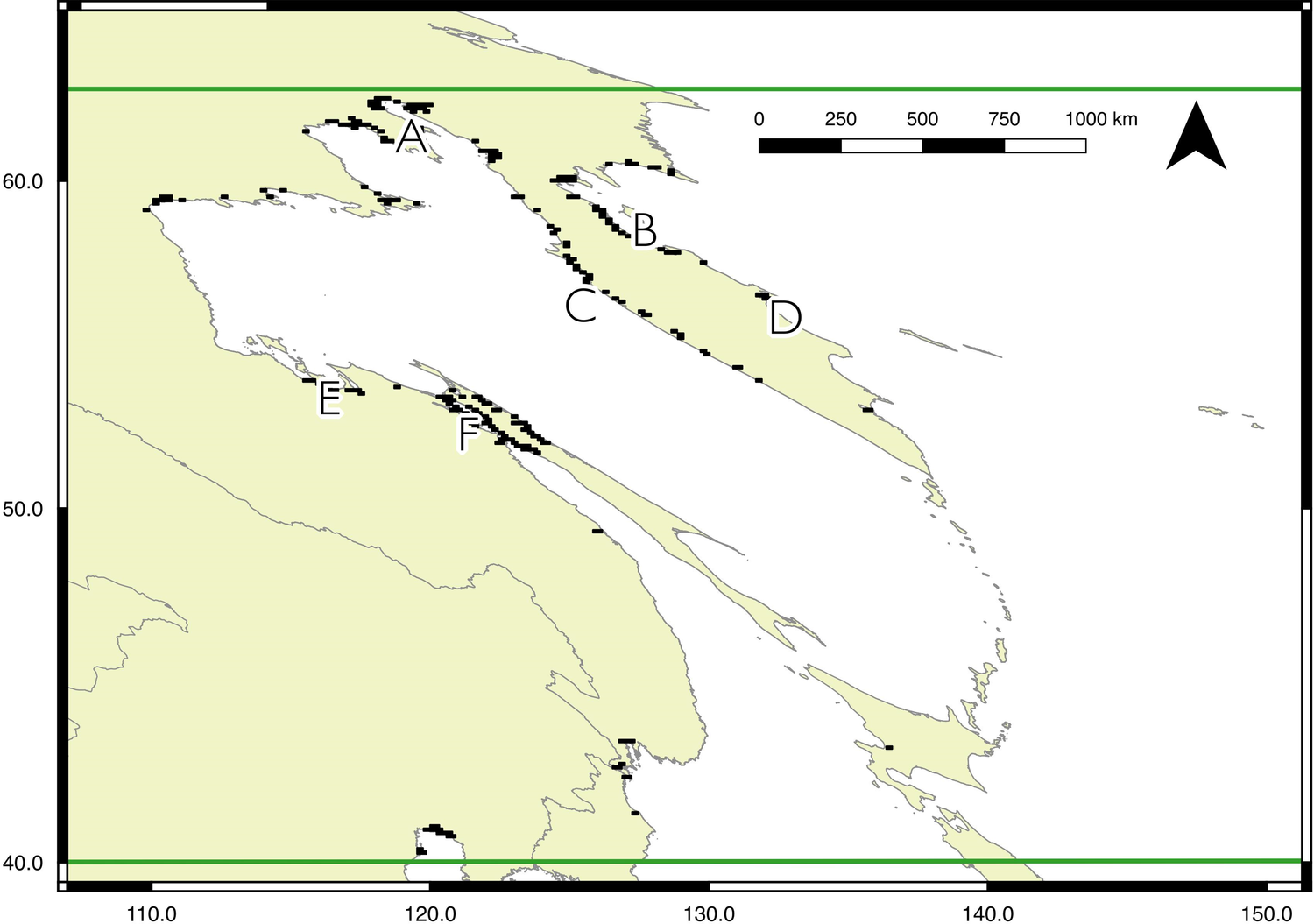
Figure 2. Predicted Spoon-billed Sandpiper distribution for the north region, bounded by horizontal lines shown at 40º and 63ºN. For display purposes 500-m pixels were resampled to 10-km squares, and the 5% of squares with the highest likelihood of occupancy are shown. A full resolution version of the map is available in the online materials (Figure S2). Sites predicted to be suitable for the species are labelled thus: A - Shelikhova Bay, B - Karaginskiy and Oliutorskiy Bays, C - Western Kamchatka coast, D - Kamchatka river mouth, E - Tugurski and Academy Bays, F - Amur liman and Northern Sakhalin. Details of these sites, with coordinates and protected area status are available in Table S1.
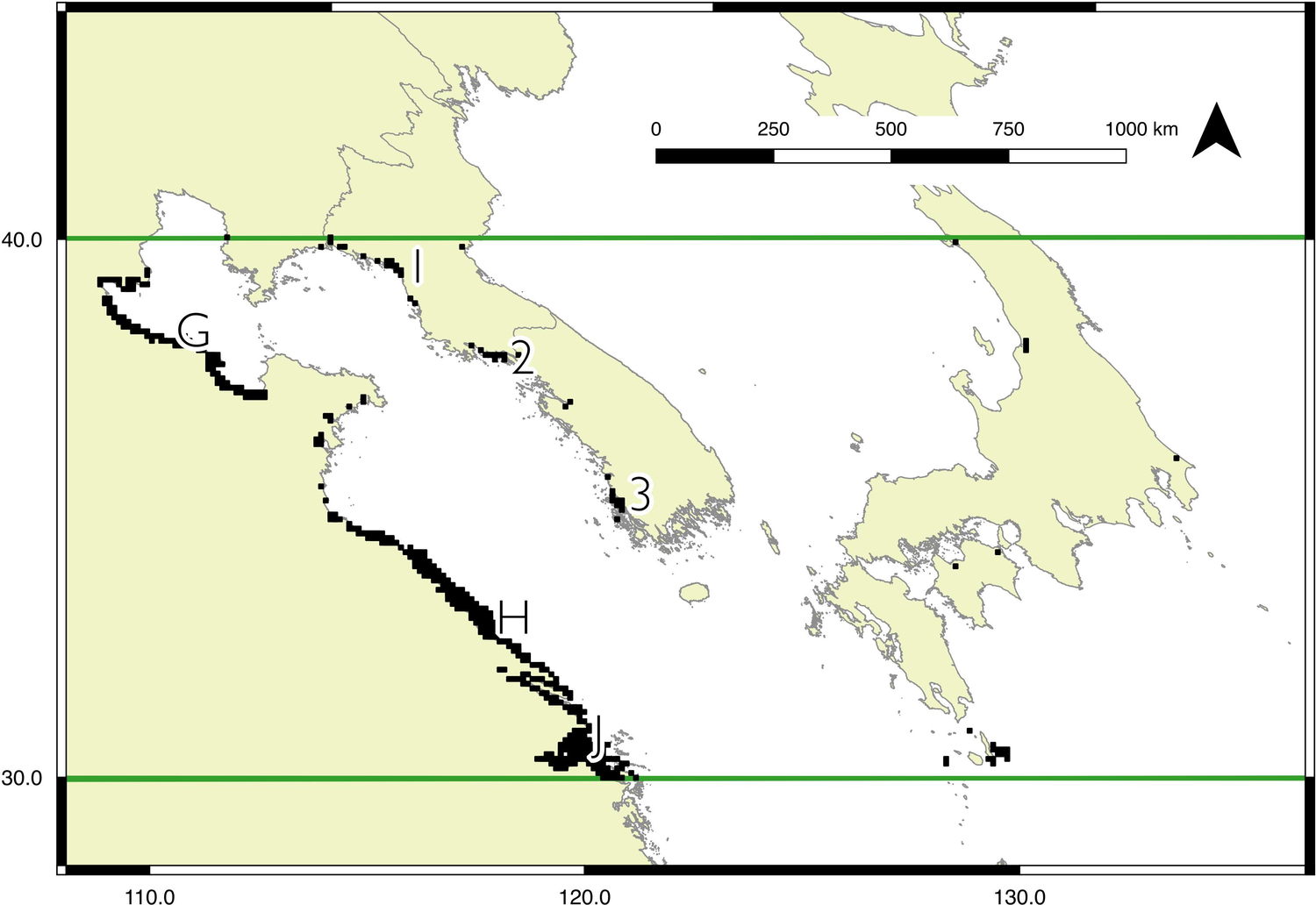
Figure 3. Predicted Spoon-billed Sandpiper distribution for the central region during migration, bounded by horizontal lines shown at 30º and 40ºN. For display purposes 500-m pixels were resampled to 10-km squares, and the 5% of squares with the highest likelihood of occupancy are shown. At this scale there is no change between the Autumn and Spring migration in the areas most likely occupied. However, a full resolution version of these maps are available in the online materials that show some subtle differences between the two time periods (Figures S3 and S4). Key sites known to be occupied are labelled thus: G - Bohai and Laizhou Bays, H - Jiangsu coast, J - Yangtze Delta and Hangzhou Bay near Shanghai. Sites predicted to be suitable by the model that have not been formally surveyed: 1 - coast at Pyongyang, 2 - Yonan coast, 3 - Jeollanam-do coast. Details of labelled sites with coordinates and protected area status are available in Table S1.
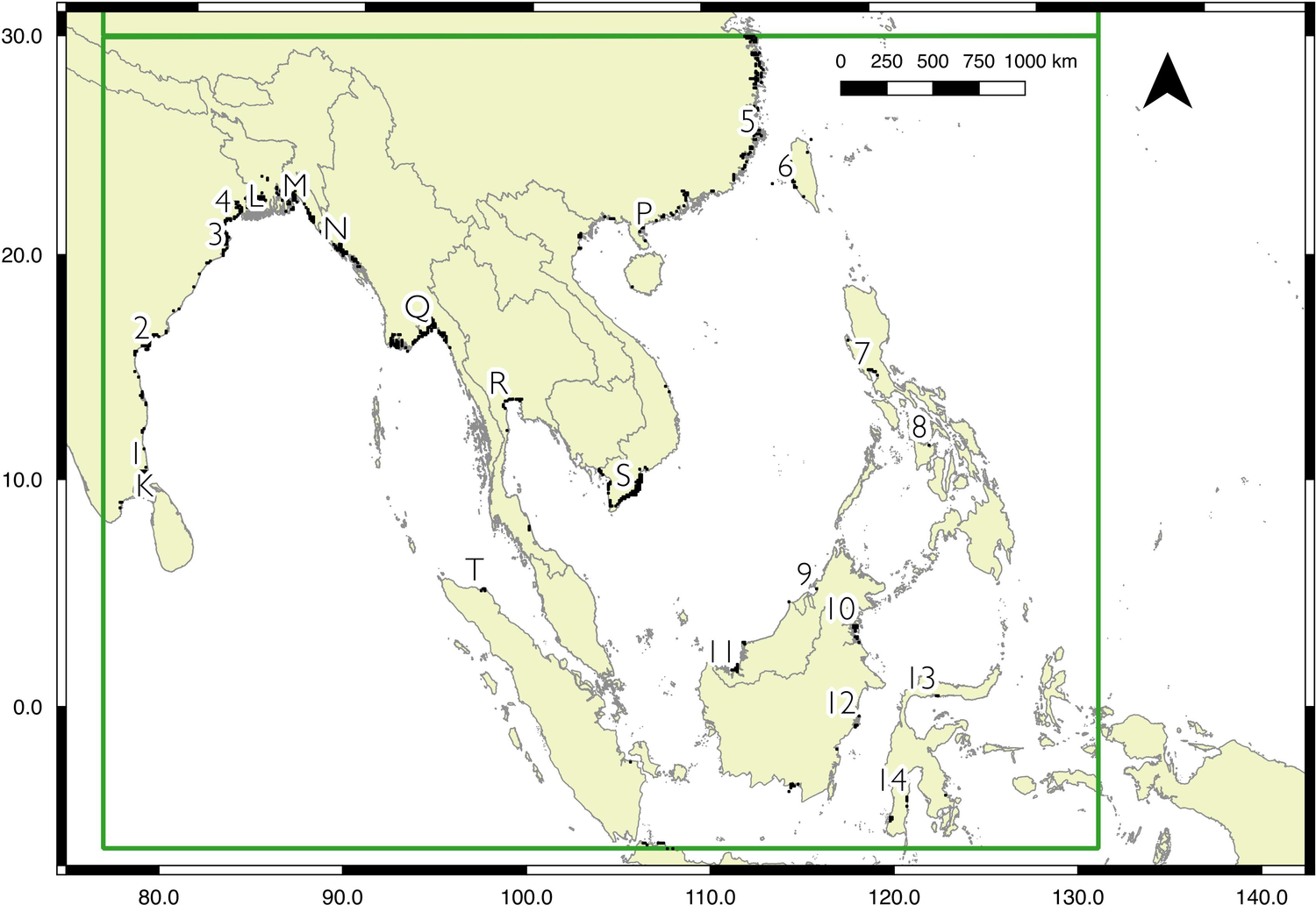
Figure 4. Predicted Spoon-billed Sandpiper distribution for the south region, bounded by the box between 6ºS and 30ºN, and between 76º and 130ºE. For display purposes 500-m pixels were resampled to 10-km squares, and the 5% of squares with the highest likelihood of occupancy are shown. A full resolution version of the map is available in the online materials (Figure S5). Sites known to be occupied are labelled thus: K - Mannar in Sri Lanka, L - Fraserganj in India, M - Ganges-Brahmaputra Delta, N – Rakhine coast and Nan Thar island, P – Hainan and the Leizhou Peninsula, Guangdong, Q - Gulf of Mottama and the Ayeyarwaday Delta, R - Inner gulf of Thailand, S - Mekong Delta, and T - Aceh in Indonesia. Sites predicted by the model to be occupied that have either not been formally surveyed between 2008 and 2017, or have never been assessed: 1 - Point Calimere, 2 - Khrishna river, 3 - Lake Chilika and Mahanadi river, 4 - Hooghly river, 5 - Fujian coast, 6 - South-west coast of Taiwan, 7 - Manilla Bay, 8 - Mindoro and Panay, 9 - Brunei Bay, 10 - Kayan river, 11 - Maludam National Park, 12 - Mahakam river, 13 - Gorontalo coast, 14 - South Sulawesi. Details of labelled sites with coordinates and protected area status are available in Table S1.
Protected areas listed in the 2018 version of the WDPA cover only 8% of the most likely occupied locations in the non-breeding range of Spoon-billed Sandpiper. Of the top 5% of areas most likely to be occupied, 15% of the north region, 10% of the central region and 5% of the south region are covered by protected areas. KBA and IBA coverage is slightly greater, covering 13% of the most likely occupied locations. In the north region 26% of sites are covered, 15% in the central region and 10% in the south.
Discussion
Spoon-billed Sandpiper distribution
This study is the first to combine field observations and satellite tracking data of the Critically Endangered Spoon-billed Sandpiper to identify areas with a high likelihood of occupancy across the entire potential non-breeding range. The models identified many potential sites that have not been formally surveyed for Spoon-billed Sandpiper and highlighted the paucity of conservation designations covering key locations throughout the flyway.
All models had high AUC values, suggesting they were appropriate for the prediction of Spoon-billed Sandpiper distribution. The main areas known to be occupied by birds in the north and central regions were successfully identified by the models. The south model highlighted all the known sites, and there is strong agreement with Zöckler et al. (Reference Zöckler, Beresford, Bunting, Chowdhury, Clark, Fu, Htin Hla, Morozov, Syroechkovskiy, Kashiwagi, Lappo, Tong, Long, Yu, Huettmann, Akasofu, Tomida and Buchanan2016) in identifying the most important sites in the core wintering range. Moreover, the model performed well in mapping the potential distribution of the species in new areas, successfully identifying locations only recently found to be occupied such as Mannar in Sri Lanka and Fraserganj in India (Chakraborty et al. Reference Chakraborty, Tripathi and Bhattacharya2018, Darshana Reference Darshana2018), although the latter might only serve as a stopover site as extensive winter surveys in the past did not record Spoon-billed Sandpiper (Zöckler et al. Reference Zöckler, Balachandran, Bunting, Fanck, Kashiwagi, Lappo, Maheswaran, Sharma, Syroechkovski and Webb2005).
Identification of new populations or new areas of suitable habitat is frequently cited as one of the purposes of species distribution models. Encouragingly, earlier attempts to model Spoon-billed Sandpiper distribution resulted in discovery of previously unknown occupied sites (Zöckler et al. Reference Zöckler, Beresford, Bunting, Chowdhury, Clark, Fu, Htin Hla, Morozov, Syroechkovskiy, Kashiwagi, Lappo, Tong, Long, Yu, Huettmann, Akasofu, Tomida and Buchanan2016, Chowdhury et al. Reference Chowdhury, Foysal, Diyan and Ahmed2018). The models presented in this study highlighted numerous potential passage sites, and the findings can be used for planning future formal survey efforts. Details of occupied and potential sites, with coordinates and protected area status are available in Table S1. The majority of potential locations in the north region are in the vicinity of areas previously identified as stopover sites for Spoon-billed Sandpiper and a range of other shorebird species (Antonov and Huettmann Reference Antonov and Huettmann2004, Gerasimov Reference Gerasimov, Boere, Galbraith and Stroud2006, Tomkovich et al. Reference Tomkovich, Porter, Loktionov and Niles2013, Aharon-Rotman et al. Reference Aharon-Rotman, Bauer and Klaassen2016). However, there are several sites around the Shelikhova Gulf that merit further investigation, including Gizhiga, Mametchinskiy and Rikiniki Bays. In the central region the most likely candidates for previously unrecognised sites are on the west coast of the Korean peninsula. In the Democratic People’s Republic of Korea there is a small area close to Pyongyang, and a large stretch of coastline in Yonan county. Satellite tagging data from two birds has since shown Yonan to be an important moulting site for Spoon-billed Sandpiper (Green et al. Reference Green, Clark, Anderson, Weston and Hughes2018). In the Republic of Korea there are extensive areas of tidal flats on the Jeollanam-do coast and the west coast of the Republic of Korea is already known to support internationally important concentrations of wading birds (Moores Reference Moores1999, Reference Moores2006, RSIS 2018), although Spoon-billed Sandpipers have not yet been recorded there.
In the south region there are numerous potential areas that merit formal surveying. Stretches of the east coast of India were identified with a high potential for occupation: between the Hooghly and Mahanadi rivers, north of the Khrishna river, and at Point Calimere in the far south. There are historic records of Spoon-billed Sandpiper from both Point Calimere and the Lake Chilika-Mahanadi Delta area, all sites that host large numbers of other migratory waterbirds (Balachandran Reference Balachandran, Boere, Galbraith and Stroud2006, Ghosh et al. Reference Ghosh, Pattnaik and Ballatore2006, RSIS 2018). The Fujian coastline in China, and Changhua and Tainan counties on the west coast of Taiwan all have a high likelihood of occupancy. There are informal, historic records of Spoon-billed Sandpiper from these areas (Bunting and Zöckler Reference Bunting, Zöckler, Boere, Galbraith and Stroud2006, eBird 2018), and the species was successfully recorded during surveys in Fujian in 2019. Wenzhou Bay in Zhejiang also appears suitable and has recently been found to be an important stopover site for Great Knot Calidris tenuirostris (Chan et al. Reference Chan, Tibbets, Lok, Hassell, Peng, Ma and Piersma2019). In the Philippines, Manila Bay, the south end of Mindoro, and parts of Panay all appear highly suitable, although there are no records of Spoon-billed Sandpiper from these areas. There are several prospective areas in Borneo, including Brunei Bay IBA, the Sadong-Saribas coast IBA in Sarawak, and the Kayan and Mahakam river estuaries on the East coast of Kalimantan. In Sulawesi there are small areas in the Tanjung Panjang KBA on the south coast of Gorontalo province, and on both east and west coasts of South Sulawesi. The sites in Borneo and Sulawesi are more speculative as there are no records for the species on these islands, although many of the sites are recognised as important for other shorebird species (BirdLife International 2018).
Conservation implications
Intertidal habitats on the East Asian-Australasian flyway are imperilled by a range of threats including pollution, invasive species, sea level rise, habitat loss and hunting (Studds et al. Reference Studds, Kendall, Murray, Wilson, Rogers, Clemens, Gosbell, Hassell, Jessop, Melville, Milton, Minton, Possingham, Riegen, Straw, Woehler and Fuller2017, Sutherland et al. Reference Sutherland, Alves, Amano, Chang, Nicholas, Finlayson, Gill, Gill, González, Gunnarsson, Kleijn, Spray and Székely2012). In the case of habitat loss for example, over 40,000 ha of intertidal flats were destroyed at the Saemangeum estuary in the Republic of Korea following construction of a 33 km long sea wall (Rogers et al. Reference Rogers, Moores and Battley2006). This had dramatic consequences for wading bird populations in the area, up to 200 Spoon-billed Sandpipers were recorded during the 1990s, but once the estuary was enclosed in 2006, this dropped to only three individuals (Barter Reference Barter2002, Moores et al. Reference Moores, Rogers, Koh, Ju, Kim and Park2008, Reference Moores, Rogers, Rogers and Hansbro2016). Austral migrant wading bird species reliant on stopover sites in the Yellow Sea have undergone severe declines in recent decades, arguably as a result of habitat loss and disturbance (Studds et al. Reference Studds, Kendall, Murray, Wilson, Rogers, Clemens, Gosbell, Hassell, Jessop, Melville, Milton, Minton, Possingham, Riegen, Straw, Woehler and Fuller2017). Despite documented declines in site quality in the Yellow Sea, many species have not shifted from traditional areas, implying a lack of alternatives (Zhang et al. Reference Zhang, Ma, Choi, Peng, Bai, Liu, Tan, Melville, He, Chan, Van Gils and Piersma2018). Given commonalities in ecology, Spoon-billed Sandpiper are likely impacted in a similar way, threatened by declining habitat quality but unable to shift to alternative sites.
Such strong site limitation during migration emphasises the precarious situation of Spoon-billed Sandpiper and the species’ sensitivity to further habitat disturbance or destruction. In consequence, recognised staging sites such as the Jiangsu coast are critically important, as the entire population may stop over at these sites during spring and autumn migration. These stopover sites are vital links in the species’ movements along the East Asian-Australasian flyway, and loss of such sites might consign Spoon-billed Sandpipers to extinction (Tong et al. Reference Tong, Zhang, Jing, Zöckler and Clark2012). Furthermore, sites predicted to have a high likelihood of occupancy by Spoon-billed Sandpiper are also likely to be used by other wading bird species of conservation concern such as the Endangered Nordmann’s Greenshank Tringa guttifer and Great Knot, increasing sites’ importance for biodiversity conservation generally (Zöckler et al. Reference Zöckler, Li, Chowdhury, Iqbal and Chenxing2018).
Preventing declines and extinctions of wading bird species in the East Asian-Australasian Flyway would be assisted by a cohesive network of protected areas. Throughout the East Asian-Australasian flyway, there is growing governmental recognition of the need to protect threatened coastal areas. China recently initiated new environmental protection legislation, declaring that strict controls would be placed on land claim projects in the Yellow Sea area (State Council 2018) and nominating priority Yellow Sea coastal wetlands as Natural World Heritage Sites (UNESCO World Heritage Centre 2017). New Ramsar sites have also been designated in Myanmar and the Republic of Korea amongst others (RSIS 2018). However, only a small fraction of the locations potentially suitable for Spoon-billed Sandpiper are currently covered by nature conservation designations. While it is unfeasible to protect or manage every potentially suitable location, many of the areas deemed most likely occupied but as yet without Spoon-billed Sandpiper records are IBAs, and therefore recognised for their importance for wider biodiversity. Unfortunately, IBAs often have little or no formal protection (BirdLife International 2018).
Given the level of threats faced by Spoon-billed Sandpipers and other wading birds in the flyway, existing protected areas must be respected. Unprotected, but potentially highly suitable sites should be surveyed for Spoon-billed Sandpiper as soon as possible, in order to guide site-based conservation management, including expansion of the protected area network (Zhang et al. Reference Zhang, Wang, Zhang, Ouyang, Chan, Crosby, Watkins, Martinez, Su, Yu, Szabo, Cao and Fox2017). Further conservation interventions should be encouraged, education and advocacy work has proven effective at reducing hunting pressures in known areas (Bird et al. Reference Bird, Lees, Chowdhury, Martin and Haque2010, Htin Hla and Eberhardt Reference Htin Hla and Eberhardt2011, Chowdhury Reference Chowdhury2012, Clark et al. Reference Clark, Pain and Green2014). However, for these to be successful in the longer term, outside engagement must be maintained, and supported by funding for alternative livelihoods (Chowdhury Reference Chowdhury2010, Pyae Phyo et al. Reference Pyae-Phyo, Moses, Clark, Anderson, Hilton, Buchanan, Zöckler and Green2018). The distribution models presented here identify priority areas for future surveying, and conservation intervention and protection.
Supplementary Materials
To view supplementary material for this article, please visit http://dx.doi.org/10.1017/S0959270920000398.
Acknowledgements
Our thanks to the many surveyors who conducted the field surveys throughout the non-breeding range. The Satellite tagging team is a collaboration between Birds Russia, Nanjing Normal University, RSPB, WWT, BTO, SBS TF and many individuals. Tracking data were curated by WWT. We are grateful to two anonymous referees for their helpful comments.
Field surveys in Russian non-breeding grounds were supported by RSPB, MHS and NABU. Field surveys in Gulf of Mottama were partly supported by BBC Wildlife Fund. Satellite tagging data collection was partly supported by The Biodiversity Investigation, Observation and Assessment Program (2019–2023) of the Ministry of Ecology and Environment of China, RSPB and a private donor. Bangladesh Spoon-billed Sandpiper Conservation Project’s fieldwork in Meghna Estuary (2015–2016) was supported by RSPB. Data collection by EL was partly supported by Basic research program (budgetary funds), projects number АААА-А19-119022190168-8 and АААА-А19-119021990093-8). PT was supported by Moscow State University Grant for Leading Scientific Schools "Depository of the Living Systems" in the MSU Development Program framework. TBL was funded by the Natural Environment Research Council UK and the IAPETUS Doctoral Training Partnership.









 Open access
Open access