



Introduction
The application of molecular analyses alongside traditional environmental techniques has huge potential for the examination of archaeological sites with either on-site or off-site proximal (e.g. under 10m) sedimentary records. Here, we present a suite of biomolecular approaches to the archaeology of prehistoric lake settlements in Scotland and Ireland to demonstrate how these techniques can be used to establish site chronologies and types of on- and off-site human activity, and to assess site erosion—all of which are of relevance to lake settlement sites found elsewhere in Europe and beyond. The definition and dating of crannogs is challenging (see below); generally, however, they are dwelling sites located on natural or artificial islands near the shores of lakes and estuaries, and they date from the Neolithic (4000–2200 BC) to the later historic period (c. 1600 AD) (Garrow & Sturt Reference Garrow and Sturt2019). Although crannogs sensu stricto are limited to Ireland, Scotland and, very occasionally, further afield, the methodology outlined here is applicable to any sites in or adjacent to waterbodies, such as lakes, estuaries and wetlands. In this article, we provide summaries of these new methods using crannog and lake settlements as case studies.
Perspectives on crannogs and lake settlements
Lake settlements such as crannogs present a number of archaeological contradictions. They are common but restricted to certain geographical areas; they are highly visible in the landscape yet surrounded by water, which restricts access and makes them difficult to excavate. They can also be legally ‘protected’ but almost un-protectable, being subject to erosion and degradation. These contradictions, along with the recognition generated by the discovery of the Alpine lake villages, fuelled antiquarian interest in the nineteenth century (e.g. Munro Reference Munro1882; Wood-Martin Reference Wood-Martin1886) and led to the first modern excavations of crannogs, such as at Loch Kinellan, Scotland, in 1914–1916 (Morrison Reference Morrison1985). Early, and many subsequent, excavations of these sites were undertaken within drained lake basins, revealing several to be lakeside settlements rather than true crannogs in the sense of an artificial island within a lake, wetland or estuary (Darvill Reference Darvill2008). This situation highlights the fact that sites such as these rarely fit into neat nomological, or indeed chronological, categories (Fredengren Reference Fredengren2002); natural islands were frequently used for crannog and wetland village construction, as were lake edges and promontories, including, for example, Glastonbury Lake Village in England, Lough Catherine in Ireland and Cults Loch in Scotland (Hencken Reference Hencken1951; Coles & Minnitt Reference Coles and Minnitt1995; Cavers & Crone Reference Cavers and Crone2017; Fonville et al. Reference Fonville, Brown, Clarke, Cavers and Crone2017). A further challenge for the interpretation of crannogs is the persistent assumption—based mostly on their relatively small size—that they were probably never unitary settlements (i.e. stand-alone settlement sites), but rather, formed part of a dispersed pattern of habitation or activities integrated with contemporaneous dryland sites. These contradictions and questions have inspired the research reported here for south-west Scotland and Northern Ireland.
Estimates of the number of crannogs in Scotland and Ireland vary for several reasons, including definitional uncertainties and the uneven distribution and coverage of crannog surveys. The latter include the Republic of Ireland, south-west and north-east Scotland and the Northern Isles (Morrison Reference Morrison1985; Farrell et al. Reference Farrell, Kelly and Gowan1989; Stratigos & Noble Reference Stratigos and Nobel2018; Cavers & Henderson Reference Cavers and Henderson2002; Henderson et al. Reference Henderson, Crone and Cavers2003, Reference Henderson, Holley and Stratigos2021; Stratigos Reference Stratigos2021). The overall distribution of crannogs is relatively even across both countries, with some probably genuine gaps, such as in south-west Ireland and south-east Scotland, and hotspots, such as in the northern midlands of Ireland (Figure 1).
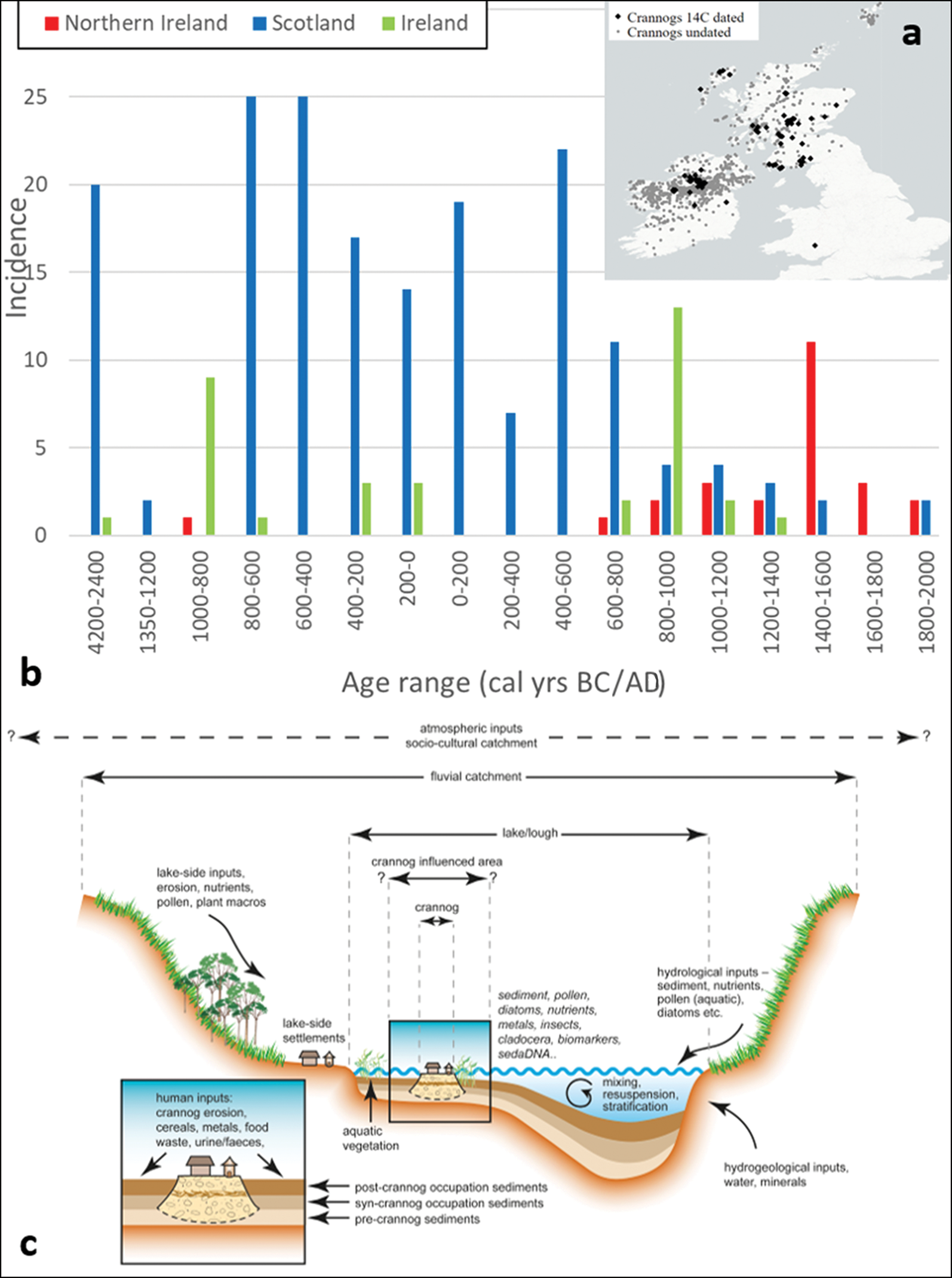
Figure 1. a) Map of crannogs recorded in the dataset; b) age distribution for Scottish and Irish crannog radiocarbon dates using median ages estimated by Calib 8.2 (Reimer et al. Reference Reimer2020); c) generalised stratigraphic model of a lake crannog and associated sediment inputs (adapted and updated from Brown et al. (Reference Brown2021)).
Beyond Scotland and Ireland, crannogs have been reported from Solvig in Denmark and in the Baltic States (Hertz Reference Hertz1974; Pranckėnaitė Reference Pranckėnaitė2014), but the most obvious outlier to their distribution is Llangorse Lake in South Wales—an early medieval royal site constructed between AD 889 and 893 (Lane & Redknapp Reference Lane and Redknapp2019). This site was probably constructed by Irish craftspeople for a king of Brycheiniog, who claimed to have been descended from a part-Irish dynasty. The historical narrative for this crannog is especially pertinent in drawing attention to the elite/royal connections of many crannogs, as is the unusually rich material culture of Llangorse that includes bone, wood, pottery, lead, iron, flint and richly embroidered textiles. Although richer than any other recently excavated crannog, this wealth of material is in line with royal crannogs, such as Lagore (Hencken Reference Hencken1950). The material culture recovered from many other crannog excavations, such as at Buiston and Cults Loch in south-west Scotland and Newtownland in Ireland, is less spectacular, but is indicative of life-ways (Crone Reference Crone2000; Cavers & Crone Reference Cavers and Crone2017; Crone et al. Reference Crone2018). Given the archaeological importance of these crannog sites, together with the difficulty of excavating them (especially those still surrounded by or under water), alternative ways of assessing and evaluating their archaeology are needed.
The site-halo effect
All archaeological sites have some effects on the surrounding areas beyond the immediate focus of settlement activity. These effects, however, are more detectable in those environments that preserve both organic and inorganic materials, such as marshes, floodplains, rivers and lakes. Over the last 20 years, palaeolimnologists have demonstrated the absence of strong water mixing and fine sediment redistribution in many small lakes, and the causes of non-uniform sediment distribution (lack of sediment focusing) caused by factors such as the spatial variation of inputs and wind effects (e.g. Morales-Marin et al. Reference Morales-Marin, French, Burningham and Battarbee2017). As a result, sedimentation and adsorption (binding of molecules to particle surfaces) can be highly localised, effectively taking the form of diffusive plumes of biological materials from point sources to create a biogeochemical halo around a site. This localisation is inconvenient for palaeolimnologists seeking general, lake-wide trends but good for archaeologists, as it means that whatever gets washed off an adjacent island, promontory or platform, or through a pile-type structure, often remains close to the site, and can be preserved within nearby sediments. An excellent example is provided by the artefacts and biological evidence found beneath pile-dwellings in Zurich (Heiss et al. Reference Heiss2017).
The size of these site-haloes depends upon the quantity of biomaterial emitted, its properties and, particularly, whether it is fixed in place by sediment or marginal vegetation. Most biomolecules are relatively large, and both lipids and sedaDNA readily adheres to, and may even be absorbed by, mineral aggregates and organic compounds in lake sediments (Capo et al. Reference Capo2021). Site haloes, which can be entirely biochemical with no associated increase in sediment thickness, extend out from settlement edges to variable distances, although generally less than 50m (Figure 1c). This distance is partly dependent upon the lake bathymetry around a site location, as submerged bench-edges or cliffs would affect the movement of biomaterials (Fonville Reference Fonville2015). Site biogeochemical haloes also contain material deliberately ejected into the water, from swords to bones.
Coring vs excavation
Exploiting the potential of the site halo effect, we present data from lake sediment cores retrieved from the areas surrounding three crannogs using large diameter (100mm) gravity- or piston-type corers. Marginal coring, which is coring areas within the lake immediately surrounding the main settlement (e.g. a crannog), is performed on the outer margin to reflect the site itself, rather than a mixture of the settlement site and the hinterland. Additionally, and in order to compare the results of this off-site approach to the evidence from the sites themselves, we undertook a test excavation on one crannog and cored through two others. Coring is only possible on sites that are predominantly made of timber, other organic matter, clay and soil—as opposed to gravel or large stone blocks—and by using narrow gouges to cut through saturated timbers. In the three cases presented here, the cores penetrated the entire structural deposits, extending down into the underlying sediments. The results of the on- and off-site approaches are compared, highlighting both the strengths and weaknesses of each. Damage to the sites is minimal: using a 60mm-diameter corer, which is the largest that is practical, and on a typical crannog measuring 30m in diameter and 3m in thickness, under 0.0005 per cent of the crannog would be destroyed. This is far less than is lost from most sites in a single year as a result of natural erosion (see discussion of erosion in online supplementary material (OSM)7)—and certainly less damaging than archaeological excavation.
Refining chronologies
The gold standard for establishing the chronology of a crannog or lake settlement would be 100 per cent excavation and dendrochronological analysis of all phases. This is rarely achievable, however, and most dates from crannogs come from a few, or even a single, timbers retrieved during diving, low water conditions or lake drainage. Whilst immensely valuable—especially in the study of individual lakes (Henderson et al. Reference Henderson, Holley and Stratigos2021)— such limited samples can only ever provide a date of construction, use or reuse and cannot normally provide a terminus ante quem or terminus post quem. If timbers can be shown to come from the base or the final phase of a crannog, then the chronology is firmer, but this is rarely possible without excavation. If dendrochronology and underwater archaeology are used, however, then the question of site contemporaneity and longevity can be addressed (Crone Reference Crone, Midgley and Sanders2012).
An alternative approach is coring, either through the crannog itself or through the adjacent lake sediments. Coring around the immediate environs of sites is relatively easy to implement and allows for dating of the stratigraphic/biogeochemical signature of the site halo from the lake sediment cores some metres away from the crannog. This approach has been undertaken for 15 crannogs (Table S1). While the general method works well, there are some issues that complicate our understanding of the stratigraphic links with the settlement site. In these cases, a reduction in organic matter, as measured by loss on ignition (decreasing LOI signifying increased clastic inputs) and an increase in the mineral component, can be used to identify the crannog unit. Titanium (Ti) is typically used, as it is stable and easily measured using scanning X-ray fluorescence (XRF). AMS radiocarbon dating also requires abundant, identifiable organic material; this is quite common at crannogs, and includes wood chips from construction. The resulting age-depth models based on radiocarbon dates from lake sediment cores typically display non-linear accumulation rates in comparison to cores retrieved from the central parts of lakes and mires (Figures 1c & 2). Results from several crannogs identify a hiatus (a pause in accumulation of sediments or erosion) at the point of construction (Figure 2; Lough Yoan North). This complicates correlation of crannog construction with any potential stimuli—either societal or environmental. There is also frequently a spread in the AMS radiocarbon dates, including reversals (common in temperate lakes), which means that age-depth model development must be undertaken with care. The causes of this are shown schematically in Figure 3 and relate to the conditions of construction, reconstruction and, critically, the modes of erosion during crannog use and/or post-abandonment.

Figure 2. Modelled age-depth from Lough Yoan North and South and White Loch of Myrton. Where boundaries are present in the model, the sections between each are highlighted by different colours. Boundaries are inserted where there is a stratigraphic change, or where there is reason to believe there could be a hiatus. Defaults are selected using OxCal v4.4 (Bronk Ramsey Reference Bronk Ramsey2009). Note the date reversals in Lough Yoan South and White Loch and the high accumulation rate during crannog phases, but variation in the overall curve. Further details of the age model development may be found in OSM1 (figure by the authors).

Figure 3. Generalised model of a typical crannog, its erosion and the likely implications for marginal (off-site) age-depth curves (on right). TAQ = terminus ante quem; TPQ = terminus post quem (figure by the authors).
Alongside this marginal (off-site) coring work, we undertook complementary on-site coring through settlement deposits at two crannogs, one in south-west Scotland (White Loch of Myrton) and one in Co. Fermanagh, Ireland (Lough Yoan South). This approach is rarely adopted due to both logistical and statutory reasons, and it is often impossible due to the presence of stones; it is, however, practical on sites constructed from only sediment, soils and timbers. The results are highly informative, revealing floors with faecal input and an estimate of abandonment date (Figure 4). This also allows tighter linkage of lake sediment cores with the chronology, construction and activity data, in the absence of crannog excavation.

Figure 4. Examples of crannog related stratigraphies: a) lake sediment stratigraphy from a transect of cores extending out from Island McHugh, Lough Catherine, Co. Tyrone, Northern Ireland; b) the through-crannog core at Lough Yoan South, Co. Fermanagh, Northern Ireland (dates in cal AD), with record of Trichuris eggs on the crannog floor prior to abandonment; and c) proximal crannog core from the crannog at Lough Yoan South (figure by the authors).
From on-site activities to societal interpretations
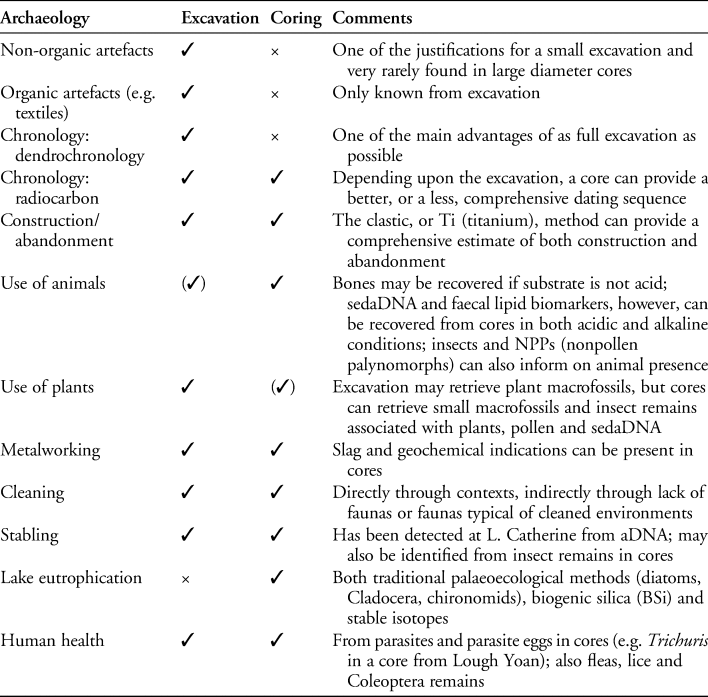
Palaeoecological work, largely in Ireland, has previously shown that crannog activities are reflected in a number of organic proxies, including charcoal, pollen and spores, midges (chironomids), beetles, water fleas (Cladocera), parasite eggs and diatoms (Brown et al. Reference Brown2005; O'Brien et al. Reference O'Brien2005; Selby et al. Reference Selby, O'Brien, Brown and Stuijts2005). The existence of a mineralogical and molecular site biogeochemical halo, as described above, however, provides the ideal conditions for the use of an orthogonal, multi-proxy design, in which the proxies are statistically independent and can therefore potentially validate an inference through falsification (Bell & Blais Reference Bell and Blais2021). The techniques used for geochemical analysis of the cores are X-ray fluorescence (XRF), X-ray diffraction (XRD), automated SEM-EDS analysis (QEMSCAN) with sedaDNA metabarcoding for plants and mammals, and lipid biomarker analyses of bile acids and faecal stanols (Table 1 and OSM1–6).
Table 1. Summary of data and proxies used in lake studies, with particular reference to the Celtic Connections and Crannogs Project.

Traditional environmental proxies all point to eutrophication of lakes around crannogs (Brown et al. Reference Brown2005; O'Brien et al. Reference O'Brien2005; Selby et al. Reference Selby, O'Brien, Brown and Stuijts2005). This is clear from diatoms (Figure 5), as well as chironomids, Cladocera and lipid biomarkers. Using a diatom transfer function (a quantitative correlation method that uses modern training data to infer past environmental conditions), we can estimate the increase in lake phosphorus (total phosphorus) around the site. The cause of this total phosphorus enrichment (and probably nitrogen enrichment) is the addition of organic matter from, for example, animals, refuse and human waste. Further inferences can be made from other traditional proxies, such as pollen. Several pollen diagrams not only show local deforestation associated with crannog construction, but also the unusual concentrations of herb types, including Plantago lanceolata (ribwort plantain) and Pteridium (bracken), which were also found in the two on-site crannog cores. The most plausible interpretation is that ribwort plantain is derived from intensive grazing of meadows near to the sites (Jones & Haggar Reference Jones and Haggar1997). The bracken, which has also been found as complete fronds within crannog sediments, was probably brought onto sites for use in animal and human bedding and/or as a roofing material, as ethnographically attested (Campbell & Pelling Reference Campbell and Pellingn.d.). At both Lough Yoan crannogs, whole bracken sporangia were present, with many at the point of spore-release, indicating that the bracken had been collected in summer (Figure 6).
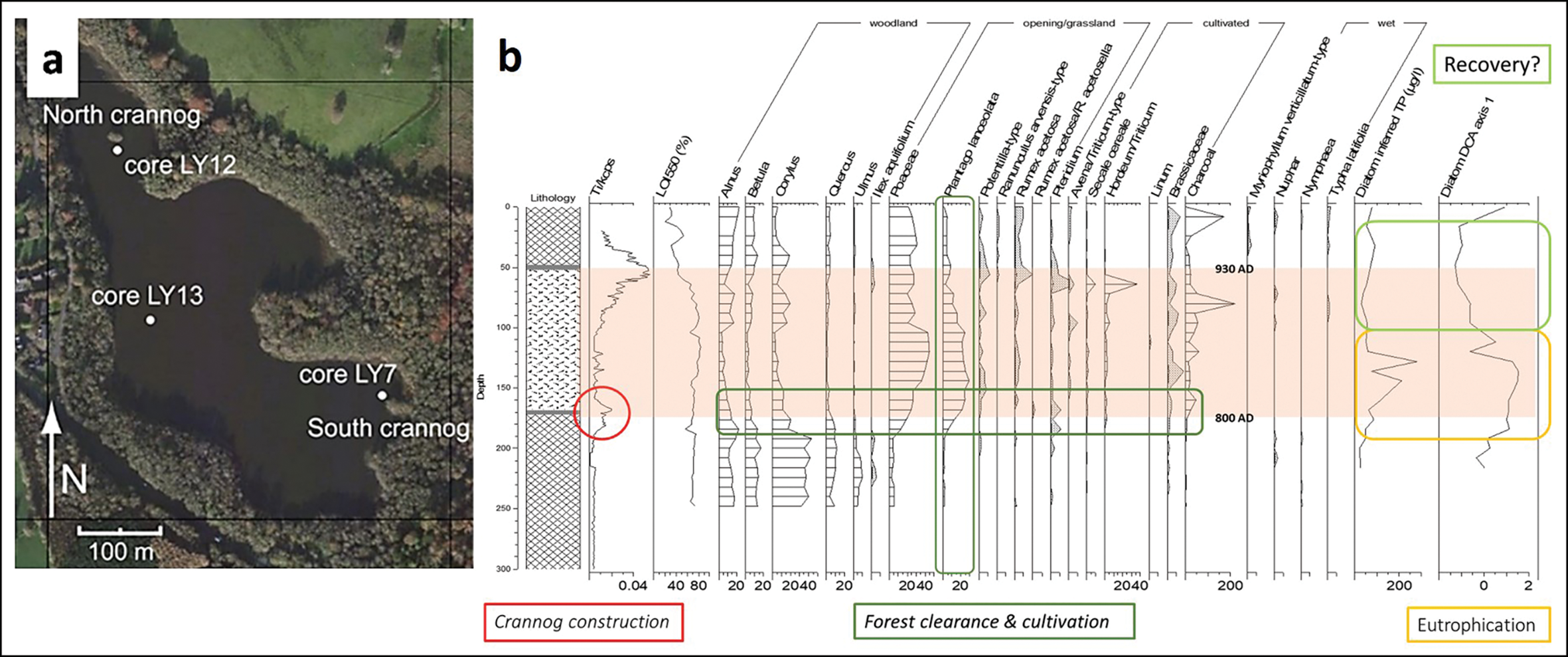
Figure 5. Lough Yoan, Co. Fermanagh crannogs: a) location of cores; b) selected traditional palaeoecological proxy data from Lough Yoan North marginal (off-site) core: titanium, selected pollen, charcoal and diatom-inferred total phosphorus, and principal components analysis results. The crannog occupation period is shaded (figure by the authors).
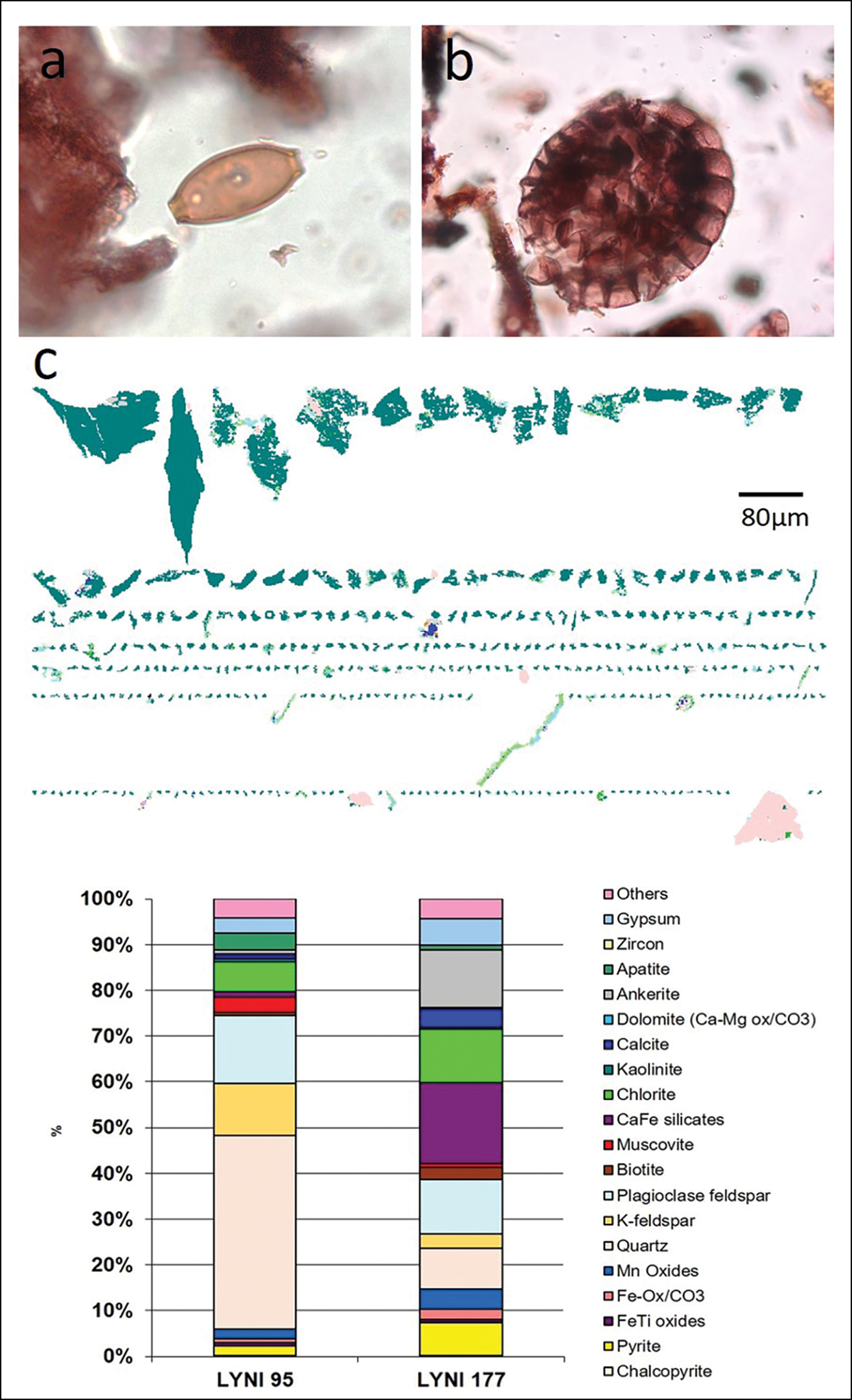
Figure 6. Microfossil and mineralogical proxies: a) Trichuris (whipworm) egg from Lough Yoan South core (850mm); b) complete sporangium of Pteridium (bracken) just about to open, from Lough Yoan South core; c) automated clast mineralogy from Lough Yoan North (950mm; crannog level) showing the micro-shards of apatite (bone) and quantitative mineralogy from the same level compared with a level prior to the construction of the crannog (figure by the authors).
Mineralogical analyses can also highlight archaeological processes. At Lough Yoan North, core automated mineralogy (QEMSAN) revealed a dominance of small, angular, porous shards of apatite at the crannog level, which is significantly different from the level below the crannog phase (Figure 6c). These are interpreted as fragments of bone from butchery waste, and support the interpretation from previous sedaDNA and bile acid analyses (Brown et al. Reference Brown2021).
Biomolecular proxies from sediments
The use of sedaDNA to reconstruct both animals and vascular plants has increased exponentially over the last few years, particularly using material extracted from lake sediments (e.g. Capo et al. Reference Capo2021; Rijal et al. Reference Rijal2021). Details of the method are published elsewhere (Brown et al. Reference Brown2021; Capo et al. Reference Capo2021; summary in OSM3). The results from the three crannogs considered here are remarkable (Figure 7). The plant sedaDNA data support the pollen evidence in the association of the crannogs with cereal cultivation, in tandem with local deforestation (and livestock raising; McCormick et al. Reference McCormick, Kerr, McClatchie and O'Sullivan2014). Both Plantago lanceolata and Pteridium sedaDNA are also enhanced at the crannog levels. The results from the mammal primer (the selective amplification of mammal sedaDNA) are even more valuable, showing that the crannog units are associated with the appearance of, or an increase in, sedaDNA of domesticated animals—principally cattle, sheep, pigs and possibly goats—as well as wild red deer. The simplest explanation for the combined evidence of eutrophication, such as ‘ruderal’, ‘weed’ and other synanthropic species from pollen and coprophilous spores, is that these animals were being kept on the crannog, from which a combination of their dung and/or slaughter products entered the lakes. The strongest evidence comes from the Lough Yoan North crannog, where the peak of animal sedaDNA from the core has high levels of apatite micro-fragments (3.65 per cent) when compared with pre-crannog levels, which have far fewer micro-fragments (0.89 per cent) (Brown et al. Reference Brown2021). The association of crannogs with domestic animals and deer is attested through many animal bone assemblages from these sites, which have been interpreted, in some contexts, as feasting deposits (McCormick & Murray Reference McCormick, Murray and Albarella2017).
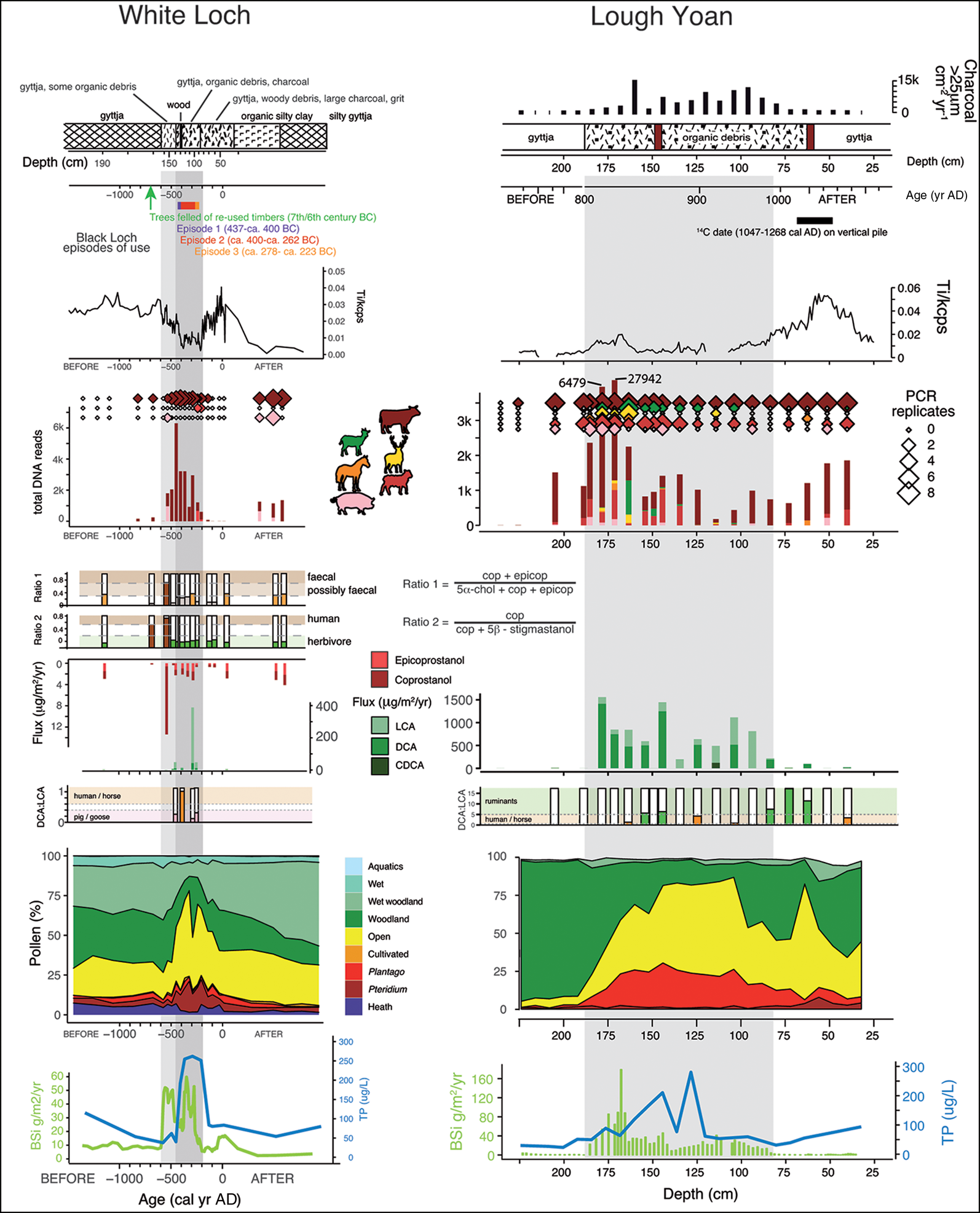
Figure 7. Selected sedaDNA and lipid biomarkers results from White Loch of Myrton and Lough Yoan North, showing from top to bottom: stratigraphy, titanium (Ti), mammal sedaDNA (bars are repeats, which are replicate samples, while diamonds are reads, which are the actual count of replicated DNA fragments), faecal stanols and bile acids, lithocholic acid (LCA), deoxycholic acid (DCA) and chenodeoxycholic acid (CDCA), pollen and total phosphorus (blue), with BSi (green) (figure by the authors).
The advantage of plant sedaDNA over pollen and spores is twofold: first, higher taxonomic resolution resulting from the ability of sedaDNA to identify to species or genus level; and second, the certainty that the species identified were growing locally. Pollen and spores retain the advantages of being able to characterise the extra-local and regional vegetation and potential quantification of land cover (through modelling), which, at present, is impossible using sedaDNA. The sedaDNA animal data—mammal primers often catch many other organisms—are highly compatible both with the coprophilous spore data and the faecal lipid biomarkers (stanols and bile acids), which also exhibit high values during the periods of crannog use (Mackay et al. Reference Mackay2020).
Support for the presence of both mammals and humans comes from the faecal stanols and bile acids, both of which increase during crannog use and have an overall relationship to the sedaDNA (Figure 7). These molecular indicators also align well with the changes in pollen and the total phosphorus and biogenic silica (BSi) record, highlighting variation in lake nutrients and productivity resulting from activities on the crannog.
The archaeological exploitation of the site biogeochemical halo through the application of increasingly sophisticated methods can reveal on-site activities, but how these analyses relate to the archaeological interpretation of crannogs depends on societal and regional contexts. Evidence for metalworking on a crannog, for example, would be interpreted in a different way if associated with similar evidence from a nearby, contemporaneous, dryland site. Ideally, therefore, crannogs should be investigated in combination with the nearby dry-land sites with which they were linked. Crannogs are also connected to their environs by their management of resources: the woodlands required for building materials and fuel; horticulture and animal husbandry; and rock and mineral exploitation for construction and processing activities, from grinding cereals to metalworking. While not able to reveal as much cultural data as excavation, coring and the techniques described here can provide remarkably detailed site narratives (Table 2).
Table 2. Summary table of advantages and disadvantages of marginal cores compared with excavation.
Crannogs: monuments at risk?
The research presented here also, inadvertently, highlights the condition of crannogs. As pointed out by Barber and Crone (Reference Barber and Crone1993), crannogs are vulnerable due to five processes:
1) subsidence due to compression of the sediment below them (unless founded on bedrock);
2) compaction of the sediment pile;
3) decay of the timber components;
4) erosion due to wave action; and
5) siltation and associated root damage.
Although restricted in number, the 15 sites studied here all exhibit signs of erosion; seven are moderately to highly eroded and two have been eroded to below normal lake levels (see OSM7). Indeed, it is precisely the threats posed by this erosion and disturbance that warrants the application of the methods of both marginal (off-site) and through-crannog coring, coupled with biomolecular analyses.
Context and discussion
The scientific techniques outlined here can provide sophisticated insights into the lifeways of societies ranging from the Neolithic to the post-medieval periods (AD 1603–present), even if via the cultural filter of wetland site activities. A key task for future work is to integrate these environmental methods with the more representative evidence generated by excavation of crannog settlements and associated dryland sites, and their wider cultural contexts. Focusing more specifically on crannogs, the frequent identification of ‘high-status’ activities and goods on some of these sites not only supports their role as places for the protective custody of valuable resources, but also suggests a degree of social exclusion combined with the display of power and wealth (O'Sullivan Reference O'Sullivan2004). Although seemingly low-status crannogs have been recorded, such as Sroove in Ireland (Boyle Reference Boyle2004), this impression may be the result of later reuse and disturbance, as the scale of resources required for crannog construction implies considerable investment. Indeed, biomolecular (and geochemical) methods may also be able to identify earlier activities that have left no trace in the on-site material culture, particularly as this could include infrequent events, such as feasting or ceremonial activities.
Shelley (Reference Shelley2009) has shown that for the medieval period in Scotland, crannogs can be regarded as watery lodges or palaces, which later went out of fashion as display became increasingly mediated through estates, mansions and gardens. The royal association of many crannogs suggests that this status display may also have been a factor during earlier periods in both Scotland and Ireland. Similarly, there was ecclesiastical use of some crannogs—especially in the early medieval period (AD 410–1066), following the model of ascetic monasticism whereby a central monastery was surrounded by satellite hermitages in remote locations and, particularly, on islands—although this was far more variable in Ireland (Bitel Reference Bitel, Alison and Bitel2020). An additional element here is the early Christian tradition of islands as places of holiness, retreat and redemption—a practise which can also have practical advantages through a degree of protection and self-sufficiency. Religious associations are visible through artefacts, such as crosses, and the later documented use of crannogs by the Church (Shelley Reference Shelley2009; Stratigos & Noble Reference Stratigos and Nobel2015). All of these considerations imply that the use and importance of crannogs were highly variable in different periods and places. One implication is that there is a fundamental limit to what we can discover about crannogs by studying them in isolation. A key insight comes from their changing use over time, but this relies entirely on increasing the number of dated crannogs, dating their construction and reuse, and comparing these dates to nearby dryland sites (e.g. settlements, raths and ringforts).
Conclusions
Crannogs and other wetland sites, although often problematic to excavate and frequently under threat, retain unrivalled potential to reveal past lifeways when compared with most terrestrial sites. Here, we describe a coring approach that exploits the biogeochemical halo of sedaDNA and lipids around crannogs in Scotland and Ireland. Using a suite of techniques, we have been able to correlate the data from lakebed cores taken from around a series of crannogs with the results of on-site coring of crannog settlement deposits. Our results indicate that this complementary approach may be more practical than previously thought. The biomolecular techniques used here have the potential to identify (and date) a range of activities that could normally only be recognised with expensive excavation, along with substantial post-excavation environmental analyses. The identification of activities such as animal keeping and slaughter, food storage, and crafting, provides the basis for interpretations of site use, including feasting and ceremonial activities—all framed within secure chronologies for construction, use and abandonment. Although we have focused here on a relatively restricted type of site—the crannog—the approach could be applied to other categories of sites located on islands, by lakeshores, or surrounded by wetlands, all of which are common in other regions of the world, dating from prehistory to the post-medieval period.
Acknowledgements
Coring through the Loch Yoan South crannog was carried out under Northern Environment and Heritage Service licence no. AE/16/18. We thank Geoff Hill and Iona Neilson for field assistance, Ludo Gielly for laboratory work in Grenoble, and the landowners who supported this work.
Funding statement
The research presented here was funded by the UK Arts and Humanities Research Council (AH/M005259/1).
Data availability statement
The sedaDNA file for L Yoan is available online from EBI-ENA (European Nucleotide Archive) under project no. PRJEB45292 (ERP129375). Data for the other proxies and sites are available by request, pending its deposition in EBI-ENA and Neotoma.
Supplementary material
To view supplementary material for this article, please visit https://doi.org/10.15184/aqy.2022.70












 Open access
Open access