



Documenting species occurrence allows us to improve our understanding of population movements and ecology, especially for species such as Bryde's whales, Balaenoptera edeni (Parsons et al., Reference Parsons, Baulch, Bechshoft, Bellazzi, Bouchet, Cosentino, Godard-Codding, Gulland, Hoffmann-Kuhnt, Hoyt, Livermore, MacLeod, Matrai, Munger, Ochiai, Peyman, Recalde-Salas, Regnery, Rojas-Bracho, Salgado-Kent, Slooten, Wang, Wilson, Wright, Young, Zwamborn and Sutherland2015). The Bryde's whale is among the least studied mysticete species; partly because sightings were often misattributed to sei whales (Balaenoptera borealis), resulting in an underestimation of Bryde's whale abundance (Würsig et al., Reference Würsig, Thewissen and Kovacs2015). Two subspecies of Bryde's whales are recognised: the small coastal form (B. edeni edeni); and the large oceanic form (B. edeni brydei) (Constantine et al., Reference Constantine, Iwata, Nieukirk and Penry2018; Cooke and Brownell, Reference Cooke and Brownell2018). Both subspecies of Bryde's whales inhabit warm and temperate waters and have a strong preference for warmer waters (more than 16°C) compared to other mysticete species (Biggs et al., Reference Biggs, Durkacz, Martin, Narvaez, De La Garza, Lombraña and Santos2017). In the North Pacific, three stocks or populations are recognised by the International Whaling Commission (IWC); the Eastern North Pacific (ENP), Western North Pacific (WNP), and East China Sea (ECS) (Donovan, Reference Donovan1991).
Bryde's whales of the ENP have been observed during scientific surveys off southern California (Smultea et al., Reference Smultea, Douglas, Bacon, Jefferson and Mazzuca2012), in the Gulf of California (Salvadeo et al., Reference Salvadeo, Flores-Ramírez, Gómez-Gallardo, Macleod, Lluch-Belda, Jaume-Schinkel and Urbán2011), and off southern coast of Mexico (Villegas-Zurita et al., Reference Villegas-Zurita, Castillejos-Moguel, Cabrera-Núñez and Ramírez-Barrragán2016). Bryde's whale distribution is influenced by prey availability (Kerosky et al., Reference Kerosky, Širović, Roche, Baumann-Pickering, Wiggins and Hildebrand2012), often associated with upwelling and thermal fronts characterised by high biological productivity (de Moura and Siciliano, Reference De Moura and Siciliano2012; Penry et al., Reference Penry, Cockcroft and Hammond2011; Palacios, Reference Palacios2003; Salvadeo et al., Reference Salvadeo, Flores-Ramírez, Gómez-Gallardo, Macleod, Lluch-Belda, Jaume-Schinkel and Urbán2011; Biggs et al., Reference Biggs, Durkacz, Martin, Narvaez, De La Garza, Lombraña and Santos2017; Castro et al., Reference Castro, Cid and Laborde2021). The influence of the El Niño-Southern Oscillation (ENSO) on Bryde's whale distribution has been noted, with fewer whales being observed during El Niño events in the Gulf of California (Salvadeo et al., Reference Salvadeo, Flores-Ramírez, Gómez-Gallardo, Macleod, Lluch-Belda, Jaume-Schinkel and Urbán2011) and off Peru (Cortez-Casamayor et al., Reference Cortez-Casamayor, Guidino and Pacheco2022). Central America is seasonally influenced by nutrient-rich upwelled waters from the Costa Rica Dome (CRD), triggered by the Papagayo wind jet off the southwestern coast of Nicaragua and the northwestern coast of Costa Rica (Fiedler, Reference Fiedler2002). Chlorophyll-a (Chl-a) is a proxy for primary production and gives an indication of phytoplankton productivity, however, since mysticetes are feeding on zooplankton and/or small fish, they tend to respond to 3–4-month time lags in Chl-a concentrations, a proxy for zooplankton production (Díaz López and Methion, Reference Díaz López and Methion2019; Visser et al., Reference Visser, Hartman, Pierce, Valavanis and Huisman2011).
Bryde's whale occurrence and distribution patterns in many Central American coastal areas remains largely unknown, and this knowledge gap precludes the implementation of regional and international protection measures (Constantine et al., Reference Constantine, Iwata, Nieukirk and Penry2018). However, their presence has been sporadically detected in Central America through both stranding and sighting data in Guatemala (Cabrera et al., Reference Cabrera, Ortíz-Wolford, Corona Figueroa and Corona2014; Quintana-Rizzo, Reference Quintana-Rizzo2019), El Salvador (Ibarra Portillo et al., Reference Ibarra Portillo, Barraza Sandoval, Pineda, Martínez de Navas, Pacas Mejía and Molina Fuentes2021), Nicaragua (De Weerdt et al., Reference De Weerdt, Ramos, Pouplard, Kochzius and Clapham2021), Costa Rica (May-Collado et al., Reference May-Collado, Amador-Caballero, Casas, Gamboa-Poveda, Garita-Alpízar, Gerrodette, González-Barrientos, Hernández-Mora, Palacios, Palacios-Alfaro, Pérez, Rasmussen, Trejos-Lasso and Rodríguez-Fonseca2018; Rodriguez-Fonseca and Cubero-Pardo, Reference Rodriguez-Fonseca and Cubero-Pardo2001), and Panamá (Castro et al., Reference Castro, Kristen, Pacheco, Diana, Romina, Galo and Kaufman2017; May-Collado et al., Reference May-Collado, Amador-Caballero, Casas, Gamboa-Poveda, Garita-Alpízar, Gerrodette, González-Barrientos, Hernández-Mora, Palacios, Palacios-Alfaro, Pérez, Rasmussen, Trejos-Lasso and Rodríguez-Fonseca2018; Rasmussen and Palacios, Reference Rasmussen and Palacios2023) (Table 1). The presence of Bryde's whales was previously recorded on the north-western coast of Nicaragua through stranding data, which involved a juvenile that died, possibly due to a vessel collision (De Weerdt et al., Reference De Weerdt, Ramos, Pouplard, Kochzius and Clapham2021). To our knowledge, there are no sighting records of Bryde’s whales reported in Nicaragua. In this paper, we present the first sightings of Bryde's whales along the southwestern coast of Nicaragua, with a description of their behaviour.
Table 1. Literature review of Bryde's whale records based on sighting, acoustic and stranding data in countries from the Eastern Tropical Pacific (ETP)

Data were collected within reasonable Beaufort sea state (1 to 3) in San Juan del Sur, Nicaragua, on a 6 m long fiberglass boat during ad libitum surveys within the cetacean conservation project of Association ELI-S. Surveys were organized between 2015–2022 during the two humpback whale breeding seasons: January and April (dry season) and July and September (wet season) with a few surveys in November, December and June, totalling a search effort of 1,608 hours spread over 300 days. Geographic positions were recorded using a handheld GPS (Garmin etrex20). Whenever an individual or group of cetaceans was encountered, the following information was collected: time of the sighting, GPS position, general group behaviour categories (milling, travelling, feeding, suspected feeding, and resting), sea surface temperature (SST), group size, age class, group type (mother-calf pair [MC], groups of two whales [G2]) and species identification. SST measurements were made with a Hecht thermometer. To aid species and individual identification, photographs of the dorsal fin and other morphological traits were taken using a D7100 DSLR Nikon camera coupled with a 100–400 mm lens. Bryde’s whale species identification was confirmed based on the presence of three longitudinal ridges that run parallel to the rostrum and a falcate dorsal fin (Jefferson et al., Reference Jefferson, Webber and Pitman2015). Individual identification was based on dorsal fin shape and nicks, which has been applied successfully on different Bryde's whale populations (Tershy, Reference Tershy1992; Steiner et al., Reference Steiner, Silva, Zereba and Leal2008; Athayde et al., Reference Athayde, Cardoso, Francisco and Siciliano2020). In mysticete species, a calf is identified as one-third to half of the body length of the accompanying adult, presumed to be the mother (Taber and Thomas, Reference Taber and Thomas1982). Sighting locations were mapped with QGIS v3.22.5 software. Bottom depth data were extracted from raster files obtained from GEBCO files (the General Bathymetrical Chart of the Oceans; www.gebco.com) with the point sampling tool, and the shortest distance to the coast was measured with the v.distance tool in QGIS. Speed was assessed based on the mean speed (km/h) of the boat tracks according to the start and the end of each sighting. Data for Chl-a were extracted from MODIS/Aqua Level-3 global monthly products (36 images in total, i.e., all months during which cetacean sightings were made) at 0.1° × 0.1° spatial resolution (Imagery produced by the NASA Earth Observations (NEO) in coordination with Gene Feldman and Norman Kuring, NASA Goddard Ocean Color Group). Level 3 products consist of monthly averaged cloud-free ocean bins of Level 2 products. Geographic coordinates of the track data were used to extract Chl-a data from the MODIS/Aqua fields using the QGIS software. Lagged Chl-a was defined as a 3-month temporal delay in Chl-a and as a proxy for zooplankton production (Visser et al., Reference Visser, Hartman, Pierce, Valavanis and Huisman2011). Lagged Chl-a was extracted from the Chl-a MODIS/Aqua fields of three months earlier.
During the focal following, a minimum distance of 50 m was maintained, and observations of behaviours were continuously recorded for each individual in the group (Altmann, Reference Altmann1974). The focal data were verbally described by a designated observer and manually transcribed by another person on the boat. The aim was to obtain a sample of at least 60-min of uninterrupted behaviours for each individual in the group. Behaviour data were processed using the Behavioural Observation Research Interactive Software (BORIS) to generate plot-coded events and time budgets (Friard and Gamba, Reference Friard and Gamba2016). Breath intervals were categorised according to blowing interval lengths, with blowing intervals shorter than 60 s defined as short breath intervals (SBI) and blowing intervals longer than 60 s as long breath intervals (LBI) (Dunlop et al., Reference Dunlop, Noad, Cato, Kniest, Miller, Smith and Stokes2013).
Four Bryde’s whale sightings were made between 10 March and 4 April 2022 (Figure 1), of which three sightings were mother and calf pairs. No sightings of Bryde’s whales were made between 2015 and 2021. The presence of three longitudinal ridges on the top of the rostrum and the absence of a white/bright colour on the lower right mandible (to exclude fin whales [Balaenoptera physalus]) confirmed that the sightings were Bryde's whales (Figure 2). Photo analysis resulted in the identification of five different individuals. The mean SST of the sightings was 25.5°C (SD = 0.9°C), the mean Chl-a was 1.3 mg m−3 (SD = 0.7 mg m−3), the mean lagged Chl-a was 19.0 mg m−3 (SD = 8.7 mg m−3) and the mean bottom depth was 68.8 m (SD = 9.1 m) (Table 2).
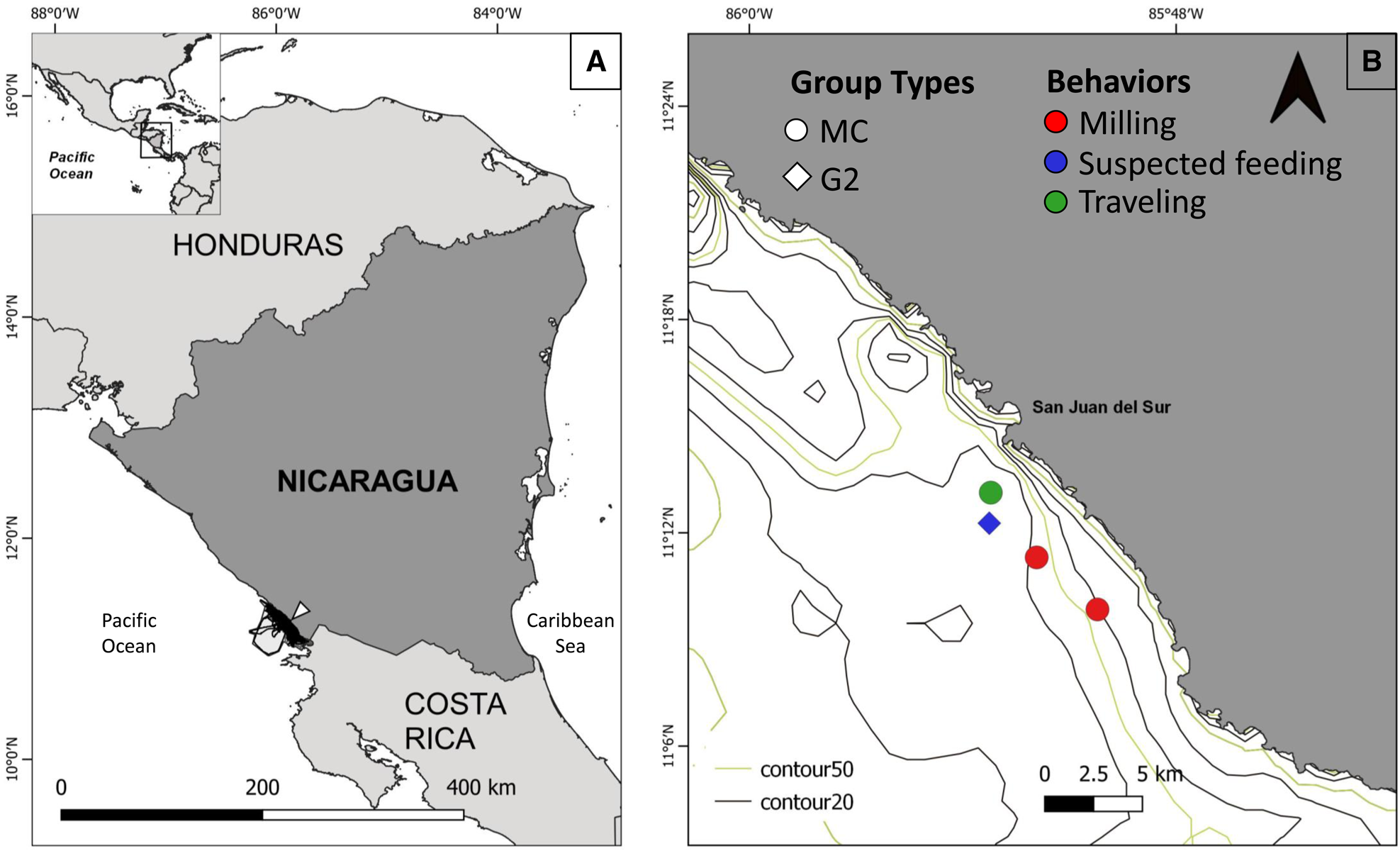
Figure 1. Location of the study site, survey tracks (black lines), and San Juan del Sur (denoted by triangle) in Nicaragua (a) and Bryde's whale sightings in San Juan del Sur (b) according to group type (MC, mother-calf; G2, two individuals) and behaviours including milling, travelling, and suspected feeding.

Figure 2. Bryde's whale species identification with the presence of (a) falcate dorsal fin (black arrow) and (b) three longitudinal lateral ridges (white arrow).
Table 2. Summary table of Bryde's whale sighting off the southwestern Pacific coast of San Juan del Sur, Nicaragua

GB, general behaviour; SF, suspected feeding; T, travelling; M, milling; G2, two whales; MC, mother-calf pair; SST, sea surface temperature; Chl-a, chlorophyll-a.
At 1508 on 10 March 2022, two adult whales were observed within a 100 m, 4195 m from the coast (Table 2). The SST recorded at the sighting location was 24.5°C, and the bottom depth was 66 m (Table 2). One individual (individual Be001) was milling in an area where schools of fish (likely anchovy species; Order Clupeiformes, Class Actinopterygii) were observed through the water surface. The general behaviour of suspected feeding was therefore assigned to this group. Humpback whales (Megaptera novaeangliae) were seen milling in the same area (approx. 500 m distance) at the same moment, moving south-eastwards, while the Bryde’s whales swam north-westward. No interactions between humpback and Bryde’s whales were inferred by the research team during the sighting. A minimum sighting distance of 100 m was maintained during the sighting. No focal following was conducted.
At 0830 on 12 March 2022, a mother-calf pair was observed at a distance of 2653 m from the coast (Table 2). The SST recorded at the sighting location was 25°C, and the bottom depth was 55 m (Table 2). Individual Be002 (mother) and Be003 (calf) were swimming south following a straight trajectory, and the general behaviour of travelling was therefore assigned to this group. A focal follow of 01 h 28 min 47 s was made on this group starting at 08 h 41 min 45s. Mean SBI was respectively 28 s (SD = 12 s; n = 25) for the mother Be002; and 28 s (SD = 16 s; n = 25) for the calf Be003. Mean LBI for the mother Be002 and the calf Be003 were 183 s (SD = 113 s; n = 25) and 180 s (SD = 89 s; n = 25), respectively (Table 3). The group travelled about 11 km from the start to the end of the sighting. This group travelled toward a location where humpback whales were seen lunge-feeding on the same day approximately 16 km further south. A minimum sighting distance of 100 m was maintained during the sighting.
Table 3. Summary table of individual Bryde's whale blow intervals, short breath interval (SBI), and long breath interval (LBI) expressed in seconds (s)

SD, standard deviation.
At 0959 on 31 March 2022, a mother-calf group was surfacing under a group of feeding frigate birds (Fregata magnificens) at approximately 3151 m from the coast. The SST recorded at the sighting location was 27°C, and the bottom depth was 77 m (Table 2). The MC pair often changed direction in the same area and were assigned the general behaviour of suspected feeding (SF). A focal follow of a total duration of 01 h 10 min 50s was made on this group starting at 12 h 45 min 32 s. Mean SBI was 36 s (SD = 12 s; n = 11) for the mother Be004; and 29 s (SD = 12 s; n = 20) for the calf Be005. Mean LBI for the mother Be004 and the calf Be005 were 211 s (SD = 166 s; n = 16) and 245 s (SD = 265 s; n = 15) respectively (Table 3).
At 1620 on 4 April 2022, a mother-calf group was observed approximately at 2747 m from the coast (Table 2). The SST recorded at the sighting location was 25.5°C, and the bottom depth was 77 m (Table 2). Individuals Be004 (mother) and Be005 (calf) were identified in this group. The individuals remained in the same area and regularly changed direction; therefore, the general behaviour of milling was assigned to this group. A focal follow of a total duration of 01 h 28 min and 47 s starting at 16 h 34 min 59 s. Mean SBI was 33 s (SD = 12; n = 31) for the mother Be004; and 37 s (SD = 14 s; n = 11) for the calf Be005. Mean LBI for the mother Be004 and the calf Be005 were 166 s (SD = 139 s; n = 26) and 182 s (SD = 142 s; n = 27) respectively (Table 3). A minimum sighting distance of 50 m was maintained during the sighting.
This is the first sighting of live (non-stranded) Bryde’s whales in Nicaragua. Bryde’s whales are known to move in search of prey (Biggs et al., Reference Biggs, Durkacz, Martin, Narvaez, De La Garza, Lombraña and Santos2017). Bryde’s whale’s prey preferences include species of mackerel, anchovy (Lodi et al., Reference Lodi, Tardin, Hetzel, Maciel, Figueiredo and Simão2015; Penry et al., Reference Penry, Cockcroft and Hammond2011) and euphausiids (Watanabe et al., Reference Watanabe, Okazaki, Tamura, Konishi, Inagake, Bando, Kiwada and Miyashita2012) on which humpback whales are also known to feed on in the area (De Weerdt and Ramos, Reference De Weerdt and Ramos2020). Indirect evidence of increased local productivity was provided by: 1) the presence of humpback whales that were lunge feeding on euphausiids a few kilometres from the sighting location on 12th March 2022; 2) the presence of flocks of feeding frigate birds flying over surfacing Bryde’s whales on 31st March 2022; and 3) observed behaviours in the field (e.g. milling). The lagged Chl-a levels for all sightings (except 31 March 2022) were relatively high (mean = 24.1 mg m−3; SD = 5.2 mg m−3), which supports our field observations on general behaviours related to suspected feeding. Feeding in Bryde’s whales is usually visually confirmed by active lunge feeding at the surface (Croll et al., Reference Croll, Acevedo-Gutiérrez, Tershy and Urbán-Ramírez2001; Goldbogen et al., Reference Goldbogen, Cade, Calambokidis, Friedlaender, Potvin, Segre and Werth2017; Iwata et al., Reference Iwata, Akamatsu, Thongsukdee, Cherdsukjai, Adulyanukosol and Sato2017; Siciliano et al., Reference Siciliano, De Oliveira Santos, Vicente, Alvarenga, Zampirolli, Brito, Azevedo and Pizzorno2004) and/or by the close proximity of other feeding bird and cetacean species (Penry, Reference Penry2010; Tershy, Reference Tershy1992). Although direct feeding behaviours were not observed during our sightings, the occurrence of subsurface horizontal lunge feeding cannot be excluded.
It is uncertain whether our observations relate to oceanographic events such as ENSO, to a temporal increase of local productivity (upwelling, CRD), and/or to increased research effort in the area in the last years. Search effort has remained similar between years since 2016 and therefore unlikely to explain these rare observations. Increased prey availability in Nicaragua can likely be due to a temporary increase in local productivity caused by ENSO climatic events. Bryde’s whales were more often recorded during ENSO – La Niña (colder and highly productive waters) conditions in the Gulf of California, (Salvadeo et al., Reference Salvadeo, Flores-Ramírez, Gómez-Gallardo, Macleod, Lluch-Belda, Jaume-Schinkel and Urbán2011), however such correlations were not found in Southern California (Smultea et al., Reference Smultea, Douglas, Bacon, Jefferson and Mazzuca2012). Warming of the ocean temperatures due to oceanographic events such as ENSO and climate change could explain distribution shifts in Bryde’s whales (Smultea et al., Reference Smultea, Douglas, Bacon, Jefferson and Mazzuca2012). It was not possible to determine whether the Bryde’s whale sightings reported in Nicaragua represent a potential range extension of the ENP population and/or the Peruvian Stock to Central America due to the low sample size and the lack of sightings in the previous years (2016–2021).
Even if direct feeding evidence was not observed in Nicaragua, the presence of prey that Bryde's whales potentially feed on, the presence of humpback whales and frigate bird feeding, and the observation of short residency time might indicate that this species occasionally visit Nicaraguan coastal areas in search of feeding opportunities. It is unclear whether oceanographic or climatic events (e.g. the CRD and/or ENSO) are responsible for the observed behaviour and/or distributional shifts of Bryde's whales toward Nicaragua. However, this deserves attention in future research. Further investigation in other Central American countries would help to understand whether Central America represents a critical habitat for the breeding and feeding the ENP Bryde's whale population.
Acknowledgements
Special thanks to our captain Noel Lugo for his work during the fieldwork. Thanks to Tom Van der Stocken for his input for the use of NASA imagery. This research was done under research permit DGPNB-IC-011-2021. Thanks to ASP and PC, and two anonymous reviewers, for their revision on this manuscript.
Author Contributions
JDW: conceptualization; data curation; formal analysis; funding acquisition; investigation; methodology; writing – original draft; writing – review and editing. CP: writing – original draft; writing – review and editing. NP: formal analysis, writing – review and editing. LB: data investigation; formal analysis; methodology; resources; supervision; visualization; writing – original draft; writing – review and editing
Financial Support
This research received no specific grant from any funding agency, commercial or not-for- profit sectors.
Competing interest
The author declares none.
Ethical Standards
Not applicable.
Data Availability
All the data are available upon request.






 Open access
Open access