



Introduction
Identifying the drivers that influence the biology and ecology of scyphozoan jellyfish is key to understanding population responses to climate and environmental variability in terms of distribution, abundance and phenology (Dawson et al., Reference Dawson, Cieciel, Decker, Hays, Lucas and Pitt2015; Goldstein and Steiner, Reference Goldstein and Steiner2020; Loveridge et al., Reference Loveridge, Lucas and Pitt2021), as well as predicting responses to future large-scale climatic changes (Holst, Reference Holst2012). Scyphozoan jellyfish are endemic in almost every temperate and tropical marine ecosystem (Dawson, Reference Dawson2004; Daglio and Dawson, Reference Daglio and Dawson2017; Jarms et al., Reference Jarms, Morandini, Schmidt-Rhaesa, Giere and Straehler-Pohl2019), often displaying large inter-annual variation in population size and timing of appearance (van Walraven et al., Reference Van Walvaren, Langenberg, Dapper, Witte, Zuur and van der Veer2015; Decker et al., Reference Decker, Brodeur, Ciannelli, Britt, Bond, DiFiore and Hunt2023).
Inter-annual variability in scyphozoan populations can be attributed, in part, to their complex multi-modal life cycles (Ceh et al., Reference Ceh, Gonzalez, Pacheco and Riascos2015), typically featuring a sexually reproducing pelagic medusa and perennial asexually reproducing benthic polyp. In seeking to understand jellyfish population dynamics, the majority of work on life histories and reproduction has focused on medusae (Lucas, Reference Lucas1996; Pitt and Kingsford, Reference Pitt and Kingsford2003; Albert, Reference Albert2005), or polyps (Fuchs et al., Reference Fuchs, Wang, Graspeuntner, Li, Insua, Herbst, Dirksen, Böhm, Hemmrich, Sommer, Domazet-Lošo, Klostermeier, Anton-Erxleben, Rosenstiel, Bosch and Khalturin2014; Widmer et al., Reference Widmer, Fox and Brierley2016). From this, it appears that the polyp life stage, which displays a high degree of plasticity in its reproductive modes (D'Ambra et al., Reference D'Ambra, Merquiol, Graham and Costello2021; Wang et al., Reference Wang, Schiariti, Xu, Ma, Sun, Wang, Zhao and Dong2023), plays a critical role in maintaining jellyfish populations and is key to the formation of true blooms (Lucas et al., Reference Lucas, Graham and Widmer2012; Lucas and Dawson, Reference Lucas, Dawson, Pitt and Lucas2014; Sukhoputova and Kraus, Reference Sukhoputova and Kraus2017). In contrast to these two ‘adult’ life stages, the planula larva is relatively understudied. Yet, this short-lived juvenile life stage potentially represents a key moment in the life cycle. Successful settlement and metamorphosis of planula larvae is essential to the replenishment of benthic polyp populations, contributing directly to the success or failure of the benthic population, and indirectly to the potential development of future jellyfish blooms (Holst and Jarms, Reference Holst and Jarms2007).
Scyphozoan planulae are ciliated, non-feeding larvae that undergo significant metamorphosis following settlement (Pechenik, Reference Pechenik1999). They respond to light (Svane and Dolmer, Reference Svane and Dolmer1995), temperature (Gambill et al., Reference Gambill, McNoughton, Kreus and Peck2018) and gravity, as well as to chemical and tactile stimuli (Tomaru et al., Reference Tomaru, Sasaki, Miyahara, Okino, Ogawa and Hamasaki2014; Yoon et al., Reference Yoon, Choi, Han and Park2014) such as salinity (Conley and Uye, Reference Conley and Uye2015; Dong et al., Reference Dong, Wang, Liu and Sun2018; Takao and Uye, Reference Takao and Uye2018), pH (Dong and Sun, Reference Dong and Sun2018), oxygen (Ishii et al., Reference Ishii, Ohba and Kobayashi2008) and competitors (Young and Chia, Reference Young and Chia1981). Settlement and development has been studied in several species, such as Cyanea capillata (Brewer, Reference Brewer1976; Holst and Jarms, Reference Holst and Jarms2010), Cyanea lamarckii, Chrysaora hysoscella and Aurelia aurita (Holst and Jarms, Reference Holst and Jarms2007; Purcell et al., Reference Purcell, Hoover and Schwark2009). Aurelia aurita planula larvae have been observed to have an increased rate of metamorphosis in the presence of established conspecifics (Gröndahl, Reference Gröndahl1989); although others attribute aggregated settlement to confounding factors such as hydrodynamics (Keen, Reference Keen1987). Due to their small size and short lifespan, research on settlement success, preferences and behaviour is confined to laboratory studies examining this life stage independently from the others.
As the planula larva is a non-feeding stage, adequate maternal provision is key to the success of the larva (Lucas and Lawes, Reference Lucas and Lawes1998; Wendt, Reference Wendt2000). Without sufficient provisioning, larvae are likely to die before settlement, or be forced to choose a suboptimal settlement location (Marshall and Keough, Reference Marshall and Keough2003). Additionally, release in sub-optimal conditions may also result in larval death and endanger the future of the benthic population. Once released from the medusa, planulae are estimated to have enough energy to survive in the water column for a few days to a week at 20°C (Schneider and Weisse, Reference Schneider and Weisse1985); although other laboratory experiments have demonstrated that planulae can survive for up to 3 weeks (Conley and Uye, Reference Conley and Uye2015). In C. lamarckii, increased temperatures have been linked to decreased settlement time (Gambill et al., Reference Gambill, McNoughton, Kreus and Peck2018); however, in A. aurita from southern UK, settlement success declined at temperatures of 18°C compared to 6°C (Webster and Lucas, Reference Webster and Lucas2012). This highlights how species and population-specific thermal tolerance limits influence response rates (Riascos et al., Reference Riascos, Paredes, González, Cáceres and Pacheco2013; Goldstein et al., Reference Goldstein, Augustin, Bleich and Holst2017).
Investigating the scyphozoan life cycle as a whole can illustrate how each life stage influences the next, thus affecting the overall population dynamics (Ceh et al., Reference Ceh, Gonzalez, Pacheco and Riascos2015; Goldstein and Steiner, Reference Goldstein and Steiner2020). We chose to embed our experimental procedure firmly in a realistic ecological context by sampling medusae across a year, and determining the parameters of our experiments using these data. Our unique blend of in situ and laboratory experiment allows us to take into account changing environmental conditions and replicate these in the laboratory under controlled conditions. This study focuses on examining how seasonal timing and temperature at larval release impacts on the quality of the reproductive output, including larvae survivorship and settlement success. Through this, we will obtain a better understanding of how ambient conditions translate into reproductive success or failure. We hypothesise that: (i) settlement success and development of planula larvae and polyps varies across the year; (ii) temperature plays a direct role in determining the successful settlement of larvae and development rates of polyps across the year; and (iii) maternal provisioning plays an indirect role in the successful settlement of larvae and development of polyps across the year. We also examine the case of temperature anomalies, specifically abnormally high temperatures, and the role these may play in determining the success of a population.
Methods
Medusae collection
Specimens of A. aurita were collected monthly from Horsea Lake, UK (50.833052; −1.093565) from February to December 2019. Horsea Lake is a brackish, semi-enclosed, man-made body of water connected to Portsmouth Harbour via a controlled pipe and valve, and the bottom (6 m) water temperature typically ranges from 5.5°C in February to 23.0°C in July (Lucas, Reference Lucas1996; CEFAS, 2018).
Medusae were collected on ten occasions throughout 2019 (Table 1). On each occasion, any specimens visible from the dockside were collected using a net and a bucket. Although medusae are present in the lake throughout the year, population biomass displays a degree of seasonality (Lucas and Lawes, Reference Lucas and Lawes1998), and numbers collected in each month are highly variable. Surface temperature and salinity were measured at time of collection.
Table 1. Sample dates and information on collected medusae

On each occasion, medusae were brought immediately back to the National Oceanography Centre Southampton, where sex, diameter (mm), wet weight (g) and condition (inverted bell, degraded bell edge, missing or additional gonads/oral arms) were recorded. Planula larvae from sexually mature females were collected into a single beaker by rubbing the oral arms, and set aside for the settlement experiments. Two gonads and two oral arms were removed and stored in 4% formalin for histological analysis.
Experimental setup
Prior to the start of each settlement experiment, eight six-well plates were preconditioned for at least 24 h in seawater to allow the development of a bacterial biofilm. Individual wells were filled with 10 mL seawater from Horsea Lake, collected at the same time as the medusae and filtered through a 1 μm mesh. Individual larvae were placed in each well of the six-well plates (n = 48).
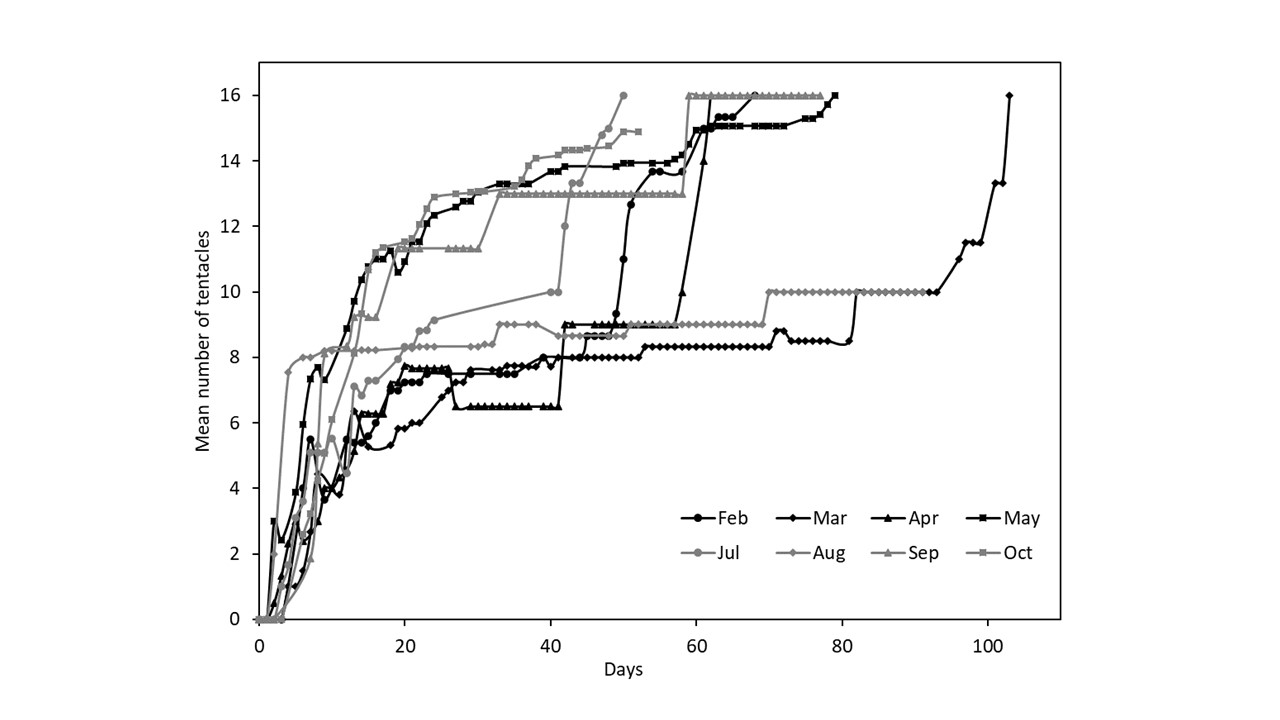
Developmental stage was recorded every 1–3 days (dead, swimming larvae or settled polyp with the number of developed tentacles) using an optic microscope. Seawater was refreshed every 5–6 days with filtered seawater of the same salinity and temperature across the experimental period. Once the first tentacles had grown, polyps were fed one 1-day-old Artemia nauplius roughly every 1–3 days (reflecting natural ingestion rate, e.g. Pengpeng et al., Reference Pengpeng, Fang, Dongjie and Song2021) by placing the nauplius on the tentacles, ensuring that the nauplii were consumed. The monthly experiments were stopped once all polyps had reached 16 tentacles or had died (31–104 days). For logistical reasons, the October experiment was stopped early on day 52 when polyps were still developing.
Proxy for maternal provisioning
Offspring size is considered a good proxy for maternal provisioning in marine organisms (see Marshall and Keough, Reference Marshall and Keough2008) with size and proximate composition often highly correlated (Berg et al., Reference Berg, Hendry, Svendsen, Bech, Arnekliev and Lohrmann2001). As variability in gamete size and organic content has been previously observed in Aurelia populations (Schneider, Reference Schneider1988), including in Horsea Lake (Lucas, Reference Lucas1996; Lucas and Lawes, Reference Lucas and Lawes1998), oocyte and planula larva sizes were used in this study as a proxy for maternal input. Dissected gonads and oral arms were dehydrated in graduated isopropanol baths and embedded in paraffin wax, before being sliced into 7 μm sections using a microtome. Sections were stained using haematoxylin and eosin (Avwioro, Reference Avwioro2011; Alturkistani et al., Reference Alturkistani, Tashkandi and Mohammedsaleh2016) and photographed using a stereomicroscope before being analysed in ImageJ. Where possible, 50 random oocytes were measured for each female medusa, with each individual oocyte measured four times to determine the average (feret) diameter.
Oral arms stored in 4% formalin were rubbed gently to release their planula larvae. For each gravid female medusa, 50 random larvae were measured using ImageJ along the longest axis according to Lucas and Lawes (Reference Lucas and Lawes1998).
Statistical analysis
All data analysis was carried out in R V4.0.3. Only individually settled replicates were analysed unless otherwise stated. All data were analysed for normality using Shapiro–Wilk tests and QQ plots, and non-parametric tests were carried out if transformation was not sufficient.
Mortality: Planula larvae mortality was analysed by comparing the percentage of deaths across the year using a logistic regression GLM to take into account the binary data (alive/dead).
Days to settlement (i.e. number of days spent in water column actively swimming): A one-way analysis of variance (ANOVA) using cube-transformed data was used to initially determine whether the number of days spent in the water column was affected by whether the planula died before it settled, or successfully settled. As this was not significant, the number of days that all planulae took to settle was compared between different months using cube-transformed data and a one-way ANOVA. Any individuals that settled and metamorphosed into polyps were included in this analysis, even if they died before reaching full maturity.
Development rate (i.e. number of tentacles produced per day): Three extreme outliers were removed from the July treatment. Individuals that did not settle or died before developing any tentacles are not included in any development rate analyses. The rate at which polyps grew their first eight tentacles is termed ‘early development rate’ and the rate at which polyps grew their final eight tentacles is termed ‘late development rate’. As neither early nor late development rate data followed normal distributions, differences in development rates between monthly experiments were analysed using a Kruskal–Wallis rank-sum test, and the difference between individual months were analysed using pairwise comparisons. In this instance, early and late development rates were analysed separately, following the same methodology. Although, A. aurita polyps have been observed to produce four primary tentacles within the first 2 days (Holst and Jarms, Reference Holst and Jarms2007), the cut-off point of eight tentacles was chosen to separate early and late development rates, due to the significant difference in development rate before and after this point (Supplementary Figure 1).
A Wilcoxon rank test determined whether there was a difference between early and late development rates. As the majority of polyps died before they reached full maturity, sample numbers varied greatly between the two treatments, so caution is advised when examining the results of this analysis. A Kruskal–Wallis rank-sum test analysed the difference in the mean maximum tentacle number grown between months. A pairwise comparison determined differences between individual treatments. The influence of temperature on development rates was examined using a Kruskal–Wallis test followed by pairwise comparisons. In this instance, early and late development rates were analysed separately, following the same methodology.
Proxy for maternal provisioning: The difference in egg Feret diameter across the year was examined using a one-way ANOVA. Planula larvae length data were log transformed to reduce positive skewness. As sample numbers differed between months, a Welch's ANOVA was used to examine differences between monthly experiments followed by a Games Howell post-hoc test to test for differences between individual treatments. A Pearson correlation determined the correlation between the average number of days to settlement and the mean planula larvae length.
Results
Mortality
On average, 8% (±9 SD) of individual planula larvae settled and survived to full polyp maturity (i.e. 16 tentacles) in any of the individual settlement experiments (Figure 1). The number of planula larvae that settled and survived to full polyp maturity was much higher in May (14 mature polyps) than in any other month (all others <5 mature polyps). Mortality in October cannot be fully determined as the experiment was not completed.

Figure 1. Percentage of individuals that survived to full maturity (i.e. 16 tentacles), or died each month as a polyp or a planula larva (excluding October).
On average, 51% (±12 SD) of planula larvae died before they settled, the fewest dying in May and July (18 larvae, 38% of total), and most in August (30 larvae, 63% of total). There was no significant difference in planula mortality across the year, nor in relation to temperature (P > 0.05).
Days to settlement
The number of days to settlement differed significantly between months (Figure 2, one-way ANOVA, F (6,329) = 14.51, P < 0.001). Planulae collected in August settled fastest, on average <5 days (P < 0.05). In contrast, planula larvae in July remained in the water column for ~12 days, significantly longer than all other months except for March.

Figure 2. Number of days to settlement of individually settled larvae across each monthly settlement experiment. Letters above data points indicate similarities (e.g. A, A), and differences (e.g. A, B) between treatments, as determined by post hoc tests.
The number of days that planulae spent in the water column did not differ between planulae that successfully settled and those that died before settlement (P < 0.05), and there is no significant correlation between days to settlement and settlement success (Pearson correlation, r = −0.02, P < 0.05).
Development rate
Early (<8 tentacles) growth rate was significantly faster than the late (>8 tentacle) growth rate (2.4 and 0.5 tentacles day−1, respectively; Wilcoxon rank-sum test with continuity correction, W = 7265.5, P < 0.001, Supplementary Figure 1). There was a significant difference in early growth rates between different months (Figure 3, Kruskal–Wallis rank-sum test, chi = 33.332, df = 7, P < 0.001). Pairwise comparisons revealed that August had a significantly faster growth rate (>4 tentacles day−1) than all months except for May and September (P > 0.05). Early growth rates in May were significantly faster (~3 tentacles day−1) than in all months except for March. Late growth rates did not vary significantly between different months (<1 tentacle day−1; P > 0.05).

Figure 3. Mean daily polyp development rate for early (<8 tentacles), and late (8–16 tentacles) across each monthly settlement experiment. No polyps grew to full maturity in August, so only early development rate is displayed for this month. Letters above data points indicate similarities (e.g. A, A), and differences (e.g. A, B) between early development rates, as determined by post hoc tests.
The average total number of tentacles grown by each polyp varied significantly between months (Figure 4, Kruskal–Wallis rank-sum test, chi = 64.289, df = 7, P < 0.001). Polyps settled in May and October grew significantly more tentacles than any other month except for February, despite the October treatment remaining incomplete (i.e. there were still polyps that had <16 tentacles alive).

Figure 4. Mean (±SE) number of tentacles grown by each polyp over the course of the experiment (blue circle), the total number of planulae that settled and metamorphosed into a polyp (white bar), and the total number of polyps that survived to maturity at the end of each experiment (grey bar). Initial number of planula larvae = 48 per month. Letters above data points indicate similarities (e.g. A, A), and differences (e.g. A, B) between the mean number of tentacles, as determined by post hoc tests.
Temperature
Polyps incubated at higher temperatures produced tentacles at a faster rate than those maintained at low temperatures (Figure 5, Kruskal–Wallis rank-sum test, chi = 33.332, df = 7, P < 0.001). Later development rates over eight tentacles did not increase with rising temperatures (P > 0.05).

Figure 5. Mean daily development rates (mean [±SD] number of tentacles grown per day) of polyps incubated at different in situ temperatures (±SE). Growth rates only included for the growth of up to the first eight tentacles. Different months with similar temperatures are circled in blue (13.4°C – 26 April and 13.7°C – 22 October; 17.6°C – 29 May and 18.1°C – 25 September). Temperatures measured during the monthly collection are summarised in Table 1.
Temperatures were broadly comparable (±1°C) at four points across the experimental period: March (10.6°C) and December (9.3°C); April (13.4°C) and October (13.7°C); May (17.6°C) and September (18.1°C); and finally June (20.5°C) and August (20.8°C). Only April and October, and May and September are compared as no gravid females were collected in June and settlement experiments were not carried out in December. Note that the October treatment is incomplete so conclusions from this comparison must be considered carefully. Early polyp development rates (Figure 3) varied significantly across the sample months, indicating that temperature has a strong effect on polyp development (Figure 5). However, a greater proportion of polyps grew to maturity in May (~30%), compared with September (<5%, Figure 1). As well as this, in October each polyp grew on average over twice as many tentacles than in April, and in May each polyp grew on average 1.5 times as many tentacles than in September (Figure 4, Kruskal–Wallis rank-sum test, chi = 64.289, df = 7, P < 0.001).
Proxy for maternal provisioning
Minimum size at maturity in female medusae varied across the year, with the smallest gravid medusae in April measuring 33 mm bell diameter compared with 190 mm in December (Figure 6). Despite these differences, across all months the minimum size at maturity corresponded to the smallest female medusae collected for that month, and all but three females had either eggs present in the gonads or planula larvae in the oral arms. Reflecting the natural population abundance in the lake, only three female medusae were collected in September, two in April and one in May.

Figure 6. Female medusae bell diameter across 2019, with minimum size at maturity for each month represented by a black circle. Sample n for each month = 23 February, 18 March, 2 April, 1 May, 5 July, 3 September, 12 October, 4 December.
Egg size varied on average between 50 and 60 μm, but did not vary significantly across the sample months, nor did it vary with temperature or salinity (P < 0.05). Neither maximum nor minimum oocyte size correlates significantly with any environmental variable.
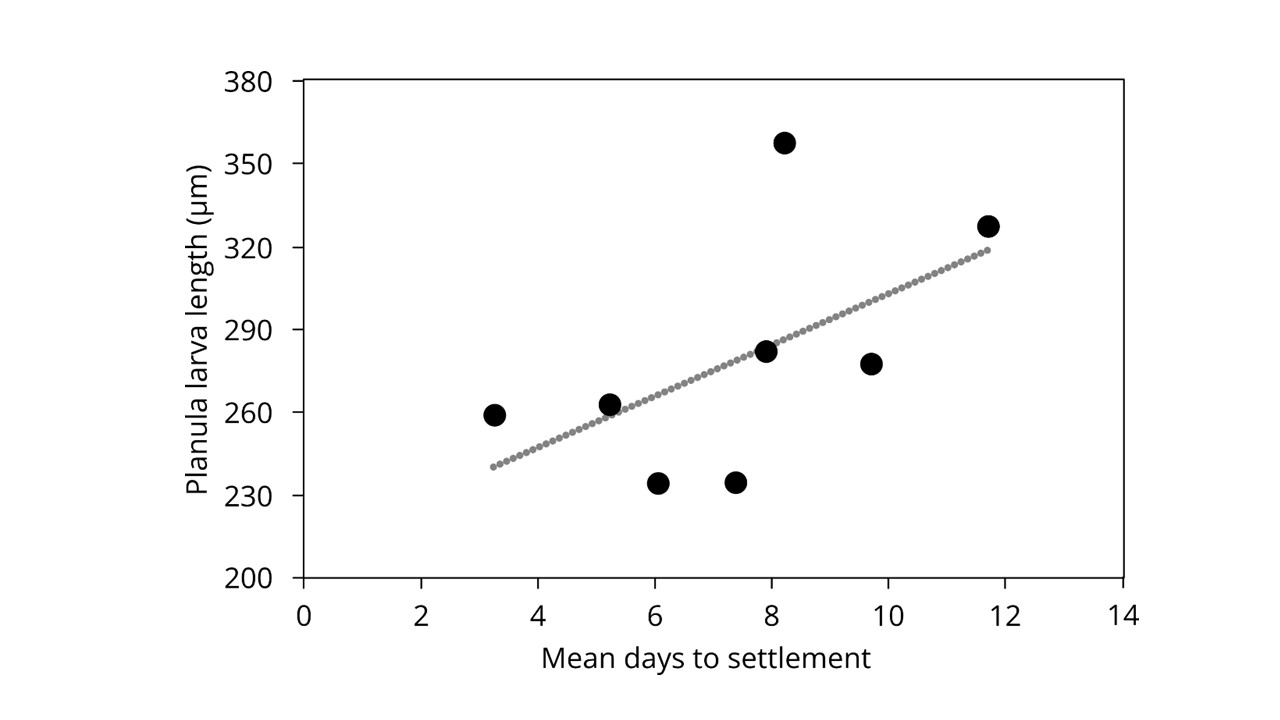
Planula larva length was significantly different across the year (Figure 7; Welch's ANOVA, F (7,262) = 26.3, P < 0.001). Planula were significantly longer in May (358 μm ± 106 SD) and July (366 μm ± 119 SD) compared to the rest of the year (Games Howell post-hoc test, P < 0.05), and were significantly shorter in April (235 μm ± 34 SD) and September (235 μm ± 46 SD) compared to the rest of the year (Games Howell post-hoc test, P < 0.05). There was a moderate positive correlation between the average number of days to settlement and the mean planula larva length (Figure S2; Pearson correlation, r = 0.56, P < 0.05).

Figure 7. Mean planula larvae length (μm) across 2019. Box and whisker plot. Letters above data points indicate similarities (e.g. A, A), and differences (e.g. A, B) between treatments, as determined by post hoc tests.
Discussion
Our results suggest that temperature drives early development rates of newly settled A. aurita polyps. Early development rates of polyps (i.e. up to 8 tentacles) were positively correlated with temperature, indicating that once planulae settled, newly formed polyps grow their first eight tentacles more rapidly at warmer temperatures. This may enable these immature polyps to start feeding earlier, thus growing to a reproductive age/size more quickly and therefore replenish the benthic population more rapidly than larvae that settle and metamorphose in the cooler months of the year. Slower-growing polyps are likely to be vulnerable to overgrowth and predation (Watanabe and Ishii, Reference Watanabe and Ishii2001; Colin and Kremer, Reference Colin and Kremer2002), as well as to running out of internal food stocks before maturing sufficiently. In contrast, polyp development rates beyond eight tentacles were not driven by temperature, and we hypothesise that this change may represent the transition point between using internal food stocks provided by the parent, and the polyp being able to catch enough of its own food.
Polyp development rates at the highest temperature (24.1°C) were slower than those at ~20°C. In our dataset, July 2019 was a thermal anomaly for this area, with temperatures much warmer than normally experienced on the south coast of the UK. Between 1984 and 2012 the mean temperature at Fawley power station (50°50′N, 1°20′W) in July was 18.8°C (±1.38), with a maximum recorded temperature of 21.6°C (CEFAS, 2018). Horsea Lake is shallow and likely to experience greater temperature extremes than coastal waters, and although previously published data from Horsea Lake note a high of 23°C (Lucas, Reference Lucas1996), the temperature recorded at Horsea Lake in July 2019 (24.1°C) was anomalously high, likely as a result of a short-term heatwave in the local area. Thermal windows constrain scyphozoan populations, limiting their geographical range (Höhn et al., Reference Höhn, Lucas and Thatje2017). Each population's thermal window differs according to the environmental conditions they experience, and this may contribute to population-specific phenology (Lucas, Reference Lucas1996; Dawson and Jacobs, Reference Dawson and Jacobs2001; Dawson et al., Reference Dawson, Cieciel, Decker, Hays, Lucas and Pitt2015). Once polyps exceed the upper limits of their thermal tolerance, typically temperatures warmer than the monthly mean for the area, a decline in aerobic metabolism is observed (Gambill and Peck, Reference Gambill and Peck2014; Höhn et al., Reference Höhn, Lucas and Thatje2017).
Our data indicate that the Horsea Lake Aurelia population may have reached its thermal limit in July 2019. This is particularly apparent when examining larval settlement. Settlement rates were on the longer side of past estimates (Schneider and Weisse, Reference Schneider and Weisse1985), with most larvae remaining in the water column for on average a few days to just over a week. Larvae released in July, coincident with the anomalously high temperature, spent more time in the water column than the other months, with the high temperatures seemingly affecting their ability to settle efficiently. The July temperature data may have been coincident with a short-term marine heatwave, although this cannot be confirmed without temperature records across a period of five consecutive days or more (Hobday et al., Reference Hobday, Alexander, Perkins, Smale, Straub, Oliver, Benthuysen, Burrows, Donat, Feng, Holbrook, Moore, Scannell, Gupta and Wernberf2016). Nevertheless, heatwaves have adverse effects on marine invertebrates, including increasing the frequency of failed reproduction and affecting recruitment and population maintenance in numerous marine taxa (Smale et al., Reference Smale, Wernberg, Oliver, Thomsen, Harvey, Straub, Burrows, Alexander, Benthuysen, Donat and Feng2019; Shanks et al., Reference Shanks, Rasmussen, Valley, Jarvis, Salant, Sutherland, Lamont, Hainey and Emlet2020; Smith et al., Reference Smith, Burrows, Hobday, King, Moore, Sen Gupta, Thomsen, Wernberg and Smale2023). Related jellyfish species such as Aurelia coerulea report negative effects of high temperatures on settlement, including smaller planula size and reduced survival rate (Dong et al., Reference Dong, Wang, Liu and Sun2018). In our experiment, there were no significant differences in settlement success over the different months. Planula larvae in July were significantly longer than other months which may have influenced their longevity in the water column, but not necessarily their ability to settle successfully. In the future, if temperatures continue to rise as predicted (Belkin, Reference Belkin2009), benthic populations may suffer in the summer months as a result of delayed settlement which increases vulnerability to the environmental and to predators. Alternatively, if temperatures rise in the winter months then larvae may encounter more favourable conditions for settlement and development, but may suffer from reduced strobilation in the spring (Loveridge et al., Reference Loveridge, Lucas and Pitt2021).
R strategists, exemplified by coastal scyphomedusae typically inhabit unstable, unpredictable environments. These organisms are characterised by high fecundity and minimal investment in the larvae, leading to offspring that settle in diverse environments, often without optimal growing conditions (Pechenik, Reference Pechenik1999). Across any of the individual settlement experiments fewer than 10% of planula survived from release to polyp maturity. Increased time spent in the water column poses significant risks to planula larvae (see review of Lucas et al., Reference Lucas, Graham and Widmer2012). They are vulnerable to predation, environmental conditions such as extreme temperatures (Gambill et al., Reference Gambill, McNoughton, Kreus and Peck2018), reduced salinity (Dong and Sun, Reference Dong and Sun2018) or being carried away from suitable settlement sites and other conspecifics which help to protect against predators and overgrowth (Marshall and Keough, Reference Marshall and Keough2003). Our results support previously published experiments that temperature is a strong driving factor behind planula larva settlement and successful development to maturity in polyps (Webster and Lucas, Reference Webster and Lucas2012). However, it does not explain all the variability we encountered in our dataset and other factors, such as maternal provisioning, should be considered.
As reported for previous years (Lucas and Lawes, Reference Lucas and Lawes1998), the proxy of maternal provisioning did not vary across the year in terms of egg size, although planula larvae were significantly different lengths across the year, indicating that there was some variation in terms of maternal influence (Lucas and Lawes, Reference Lucas and Lawes1998; Wendt, Reference Wendt2000). Planula larvae have a limited amount of time to find a suitable settlement surface (Marshall and Keough, Reference Marshall and Keough2003). As larvae age, finding a place to settle becomes more urgent (see Lucas et al., Reference Lucas, Graham and Widmer2012). Research suggests that the maximum time spent in the water column depends on not only the environment into which the larvae is released (Conley and Uye, Reference Conley and Uye2015), but also on the energetic reserves provided by the parent (Schneider and Weisse, Reference Schneider and Weisse1985; Wendt, Reference Wendt2000). In our study, the longest planula larvae were found in May and July, which may have been due to more food available for the medusa in the spring, coupled with warmer conditions enabling faster larva development and development. However, it has previously been suggested that in Horsea Lake medusae may direct food resources into somatic development when abundant, and reproductive effort when food is scarce (Lucas, Reference Lucas1996). There was no variation in substrate type, and in the absence of settlement cues, larger larvae may take longer to settle (Marshall and Keough, Reference Marshall and Keough2003).
Adaptation to local thermal conditions is a possibility in Horsea Lake. The A. aurita population there does display unusual characteristics not seen in many other populations such as the production of eggs and larvae across the year, as well as medusae being present year round (Lucas et al., Reference Lucas, Hirst and Williams1997). These factors enable us to examine planula larvae settlement and development to maturity in a range of realistic environmental conditions. Our results have provided us with informative responses to realistic temperatures; however, this approach is not without limitations. Highly variable medusa sample numbers between different months, as well as low sample number in some of the settled polyp groups means some of our conclusions could benefit from being verified by further experiments. Nevertheless, this study has further illuminated the complexity of the scyphozoan life cycle and its many driving factors. Reproductive strategy, maternal provisioning and the environmental conditions all feed into a complex model that determines planula larvae settlement success and rate of polyp development to maturity.
Conclusion
This study focuses on clarifying how timing and temperature at larval release impacts on the quality of the reproductive output, including settlement rate, success, and survivorship. Our results indicate that if studied in isolation from other life stages, increasing temperatures could appear beneficial to scyphozoan populations, by increasing early polyp development rates. However, when put in context with other parts of the life cycle, increasing temperatures past thermal limits even in the short term through the increasing occurrence of temperature anomalies is likely to be detrimental to larval settlement and indirectly to the replenishment of temperate polyp populations. Whilst temperature remains a driving force for settlement and development rates, it does not explain all of the variation that we observed in our dataset. Revisiting our hypotheses, settlement success and development of planula larvae and polyps did vary across the year. Environmental factors such as temperature were found to drive early polyp development rates; however, other factors such as maternal ‘provisioning’ also appeared to influence the successful settlement of larvae and development of polyps across the year. Finally, temperature anomalies may exceed the population's thermal limits and future increases due to climate change may lead to a decline in some temperate jellyfish populations.
Supplementary material
The supplementary material for this article can be found at https://doi.org/10.1017/S0025315424000468.
Data
The datasets generated during and/or analysed during the current study are available from the corresponding author on reasonable request.
Acknowledgements
This research was supported by the University of Southampton, and was carried out through the NERC-funded SPITFIRE Doctoral Training Partnership. We want to thank Robbie Robinson and George Clarke for technical assistance throughout the study.
Author contributions
A. L. and C. H. L. came up with the overall concept and designed the experiments. A. L. carried out the experiments. A. L. and D. F. carried out the data analysis. C. H. L., A. L. and D. F. prepared the manuscript.
Financial support
This work was supported by the Natural Environmental Research Council [grant number NE/L002531/1].
Competing interests
The authors have declared that no competing interests exist.
Ethical standards
The moon jellyfish Aurelia aurita is not a protected species in the area of study. Permission was obtained from the Royal Navy to access Horsea Lake, UK (50°49′58.8; −1°05′36.9) to collect A. aurita specimens. All international, national and/or institutional guidelines for the care and use of animals were followed where applicable.










 Open access
Open access