



Introduction
Wild mammals are facing an uncertain future as a result of human activity, with c. 20% threatened with extinction (Hoffmann et al., Reference Hoffmann, Belant, Chanson, Cox, Lamoreux and Rodrigues2011). Tropical rainforests are species-rich, globally important ecosystems, but overexploitation and habitat loss pose major threats to their biodiversity (Butchart et al., Reference Butchart, Walpole, Collen, van Strien, Scharlemann and Almond2010; Rosa et al., Reference Rosa, Smith, Wearn, Purves and Ewers2016). With the continued rise in the human population and consequent demand for forest products, rainforest species are likely to face increasing pressure (Fa et al., Reference Fa, Currie and Meeuwig2003; United Nations, 2019). Consequently, we must ensure their adequate conservation, both within and outside protected areas.
Central Africa contains the second largest area of tropical rainforest (Potapov et al., Reference Potapov, Turubanova, Hansen, Adusei, Broich and Altstatt2012), home to a unique range of biodiversity. In rural communities, subsistence hunting of rainforest mammals for food and commercial hunting for livelihoods remain important (Nasi et al., Reference Nasi, Taber and van Vliet2011; Ziegler et al., Reference Ziegler, Fa, Wohlfart, Streit, Jacob and Wegmann2016). Additionally, some of the most threatened mammals have high economic value and are subject to hunting for illegal trade (Wasser et al., Reference Wasser, Brown, Mailand, Mondol, Clark, Laurie and Weir2015; Ingram et al., Reference Ingram, Coad, Abernethy, Maisels, Stokes and Bobo2018). The scale of commercial hunting in forests has increased for several reasons, including increased urban demand for wild meat (bushmeat), human population density and access via roads (e.g. Robinson & Bennett, Reference Robinson and Bennett2004; Ziegler et al., Reference Ziegler, Fa, Wohlfart, Streit, Jacob and Wegmann2016; Fa et al., Reference Fa, Wright, Funk, Márquez, Olivero and Farfán2019).
Overharvesting of mammals is of concern as it leads to depletion and to changes in forest structure and ecosystem functioning, such as reduced seedling establishment and tree regeneration (Wang et al., Reference Wang, Sork, Leong and Smith2007; Rosin & Poulsen, Reference Rosin and Poulsen2016). In turn, a reduction in available protein can result in human malnutrition and food insecurity (Nasi et al., Reference Nasi, Taber and van Vliet2011). Despite challenges in measuring the sustainability of offtake, simple sustainability indicators can be useful (Milner-Gulland & Rowcliffe, Reference Milner-Gulland and Rowcliffe2007). For example, hunter selectivity and intrinsic demography make larger species more vulnerable to overharvesting (Peres, Reference Peres2000; Cowlishaw et al., Reference Cowlishaw, Mendelson and Rowcliffe2005); a prey community dominated by ecologically versatile, smaller mammals typically reflects such overharvesting. This pattern of species depletion is often observed close to human settlements, reflecting spatial patterns of human population density and hunting effort (e.g. Koerner et al., Reference Koerner, Poulsen, Blanchard, Okouyi and Clark2017; Beirne et al., Reference Beirne, Meier, Mbele, Menie, Froese, Okouyi and Poulsen2019). Therefore, species composition can be a useful indicator of hunting sustainability (Milner-Gulland & Rowcliffe, Reference Milner-Gulland and Rowcliffe2007).
Distance to roads can also be used as an indicator of human activity and, in some cases, hunting pressure. In Central Africa, expansion of industrial logging has resulted in an increased number of roads to transport goods and workers (Laporte et al., Reference Laporte, Stabach, Grosch, Lin and Goetz2007). Roads allow hunters to exploit previously inaccessible forest areas and many studies have linked proximity to roads to increased hunting and negative impacts on mammals (e.g. Vanthomme et al., Reference Vanthomme, Kolowski, Korte and Alonso2013; Ziegler et al., Reference Ziegler, Fa, Wohlfart, Streit, Jacob and Wegmann2016). However, roads can also affect species behaviour and abundance, resulting in positive or negative associations that are species and location dependent and therefore hard to predict (e.g. Laurance et al., Reference Laurance, Croes, Tchignoumba, Lahm, Alonso and Lee2006; Vanthomme et al., Reference Vanthomme, Kolowski, Korte and Alonso2013).
In Cameroon, community forests were made possible with the adoption of the Forestry Law in 1994, with the aims of reducing poverty in forest communities and protecting the environment (Egbe, Reference Egbe2001; Oyono et al., Reference Oyono, Ribot and Larson2006). Within them, local people can manage forest resources subject to a simple management plan that is agreed between a community-level council and the government (Egbe, Reference Egbe2001; Lescuyer, Reference Lescuyer2013). Community forests occur along roads or around villages, can be up to 5,000 ha and are granted for a period of 25 years. Although most are focused on timber extraction, villagers can maintain agricultural activities and hunting of unprotected species, for subsistence only, within community forests. However, the extent to which the biodiversity conservation and socio-economic goals of community forests are met varies. Most studies have found few, if any, benefits to the community (e.g. Ezzine-de-Blas et al., Reference Ezzine-de-Blas, Pérez, Sayer, Lescuyer, Nasi and Karsenty2009; Lescuyer, Reference Lescuyer2013), with progress hindered, inter alia, by management conflicts and a lack of technical skills (e.g. Ezzine-de-Blas et al., Reference Ezzine-de-Blas, Pérez, Sayer, Lescuyer, Nasi and Karsenty2009; Alemagi, Reference Alemagi2011). Inadequate monitoring and evaluation can allow environmental degradation to continue unchecked, and little research has been conducted on animal biodiversity within community forests (Lhoest et al., Reference Lhoest, Fonteyn, Daïnou, Delbeke, Doucet and Dufrêne2020), hindering examination of the sustainability of hunting.
Our research focuses on mammals within a community forest that surrounds a village on the southern periphery of the Dja Biosphere Reserve in Cameroon. This 5,260 km2 Reserve is a UNESCO World Heritage Site that supports c. 109 mammal species, including the African forest elephant Loxodonta cyclotis, western lowland gorilla Gorilla gorilla gorilla and central African chimpanzee Pan troglodytes troglodytes (Kingdon, Reference Kingdon2015; Bruce et al., Reference Bruce, Amin, Wacher, Fankem, Ndjassi and Bata2018a). However, a recent analysis suggests that the conservation success of this Reserve is low (Gruber et al., Reference Gruber, Mbatu, Johns and Dixon2018), partly because of an increasing human population around the Reserve, where there are hundreds of small villages and a network of production forests, mining concessions and community forests. There are limited baseline data on the status of mammals in the Dja region, but recent surveys have provided information on the mammal community of the Reserve (Bruce et al., Reference Bruce, Amin, Wacher, Fankem, Ndjassi and Bata2018a,Reference Bruce, Ndjassi, Fowler, Ndimbe, Fankem and Mbodbdac) and in three neighbouring community forests to the north-east (Lhoest et al., Reference Lhoest, Fonteyn, Daïnou, Delbeke, Doucet and Dufrêne2020). We conducted a camera-trap survey within a previously unsurveyed community forest to the south-east of the Reserve to provide baseline data on mammal species richness and composition. We used occupancy analysis to estimate the proportion of the survey area used by each species and to investigate how indicators of human activity influenced the distribution of species. We then used species composition and the relationship between human activity covariates and species occupancy to look for signs of overexploitation. Our findings could form the basis of wildlife monitoring within the study area and inform conservation management of wildlife within the community forest and wider Dja region.
Study area
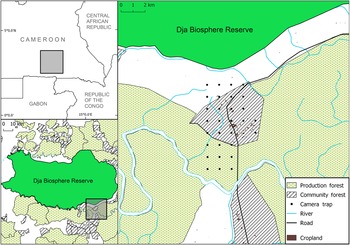
The Dja region has a humid, equatorial climate and is characterized by semi-deciduous, lowland rainforest with a mean annual temperature of 23 °C and annual rainfall of 1,600 mm (Lagarde, Reference Lagarde2004). Our study area is a village within a community forest in dense rainforest between the Dja Biosphere Reserve and the Dja River, c. 4 km south-east of the Reserve (Fig. 1). It comprises 90 households that rely heavily on forest resources for subsistence. The mean human population density is 5 people/1 km2 (Center for International Earth Science Information Network, 2018) and villagers have mixed livelihoods, primarily agricultural, but many also fish, hunt and collect non-timber forest products. The village became a community forest in 2010 with the objectives of helping the community generate income and sustainably improve their living conditions. No previous research has examined hunting within the community forest, but during a scoping trip, villagers reported to us that it was relatively easy to find food by hunting and gathering. A permanent hunting camp was encountered during the survey, which is an area where residents of the village stay for a few nights when hunting or gathering further afield. Some small-scale logging took place within the community forest in 2011–2013 but there was no logging at the time of our survey. The village is relatively isolated and far from the nearest main road, although a logging road was built through the village in 2015, which during our survey carried c. 15–20 trucks per day to nearby logging concessions. The community forest has been bordered by a logging concession to the south since 2013 and there are numerous others in the vicinity.

Fig. 1 Location of the study area and camera-trap grid, in place during August–November 2017, across the community forest and adjacent production forest in the buffer zone of the Dja Biosphere Reserve, Cameroon.
Methods
Survey design
We placed a systematic grid of 30 camera traps (Bushnell, Overland Park, USA) set 1 km apart across the village, surrounding community forest and adjacent production forest. There was no logging within the surveyed area of production forest and, as it provided a continuation of the habitat found within the community forest, we did not consider it as a separate habitat in this study. Cameras were unbaited and operational continuously during August–November 2017. They took three photographs each time they were triggered, using low glow infrared flash set to medium, with a trigger speed of 0.14 s and a 1-s delay between triggers. Suitable places to position the cameras were chosen within 100 m of each grid point, close to frequently used animal trails (Amin et al., Reference Amin, Andanje, Ogwonka, Ali, Bowkett, Omar and Wacher2015) or possible feeding spots. Cameras were attached to trees c. 4 m from the trails. The cameras had high sensitivity and were placed 30–45 cm off the ground, angled horizontally. To avoid sunlight interfering with the cameras, they were set facing north or south (Bruce et al., Reference Bruce, Amin, Wacher, Fankem, Ndjassi and Bata2018a). Tall grass and foliage that could have obstructed the field of view were cleared. The placement of two cameras close to the road had to be adjusted as they would have been within a hunting camp, but the 1 km spacing was not substantially compromised. The total survey area was c. 26 km2.
Species richness and composition
Species identification was aided by Kingdon (Reference Kingdon2015). For Herpestidae (mongoose) species, data were pooled because of the low numbers of records of individual species and difficulties of identification. We excluded images of small rodents that could not be identified to species level as their small body size limited detectability, resulting in sampling error (Tobler et al., Reference Tobler, Carrillo-Percastegui, Leite Pitman, Mares and Powell2008; Bruce et al., Reference Bruce, Amin, Wacher, Fankem, Ndjassi and Bata2018a). We generated a species accumulation curve using the package vegan in R 3.6.2 (Oksanen et al., Reference Oksanen, Blanchet, Friendly, Kindt, Legendre and McGlinn2019; R Core team, 2019) to examine whether our sampling effort was sufficient to capture the majority of mammal species and to estimate mammal species richness. To infer whether hunting was sustainable or not, species composition was compared with the terrestrial mammal community that would be expected within our study area with minimal human disturbance. The expected community was based on species whose distributions coincided with our study area according to Kingdon (Reference Kingdon2015) and the IUCN Red List (IUCN, Reference IUCN2021), were likely to be detected by ground-level camera traps within continuous forest in this area based on species ecology and habitat preferences, and were detected by camera-trap surveys in similar habitat within the Reserve (Bruce et al., Reference Bruce, Amin, Wacher, Fankem, Ndjassi and Bata2018a,Reference Bruce, Ndjassi, Fowler, Ndimbe, Fankem and Mbodbdac) or in community forests to the north-east (Lhoest et al., Reference Lhoest, Fonteyn, Daïnou, Delbeke, Doucet and Dufrêne2020).
Occupancy analysis
We used occupancy analysis as it can be used to model species distributions while accounting for imperfect detectability (MacKenzie et al., Reference MacKenzie, Nichols, Lachman, Droege, Royle and Langtimm2002, Reference MacKenzie, Nichols, Royle, Pollock, Bailey and Hines2006), and is being increasingly used as a monitoring tool (e.g. Ahumada et al., Reference Ahumada, Silva, Gajapersad, Hallam, Hurtado and Martin2011; Rich et al., Reference Rich, Davis, Farris, Miller, Tucker and Hamel2017). Species detection histories were created by recording the presence (1) or absence (0) of a species within 5-day windows at each camera trap, a compromise between having an adequate number of repeat visits and model stability. Single-species, single-season occupancy models were run using the package unmarked in R (MacKenzie et al., Reference MacKenzie, Nichols, Lachman, Droege, Royle and Langtimm2002; Fiske & Chandler, Reference Fiske and Chandler2011). As the home ranges of the larger species in our study area are typically > 1 km2, we interpreted occupancy as the proportion of sites used by a species (MacKenzie et al., Reference MacKenzie, Nichols, Royle, Pollock, Bailey and Hines2006; Da Silva et al., Reference Da Silva, Paviolo, Tambosi and Pardini2018).
We used the geoprocessing tools in QGIS 3.0.2 (QGIS Development Team, 2018) to derive indicators of human activity and habitat quality hypothesized to influence occupancy (ψ) and/or detection probability (p, the probability that a species is detected given that it occupies a site). For each camera location, we calculated proximity to the Dja Biosphere Reserve, proximity to the road, village, river and nearest production forest, altitude, human population density (Center for International Earth Science Information Network, 2018), slope and per cent of tree cover within a 200 m buffer (where land that was not forest was cropland) based on a land-cover map of Africa (ESA CCI Land Cover Team, 2017). Pearson correlation coefficients (r) were used to test for correlation between pairs of covariates; if r was > 0.7 we excluded one of the pair and kept the covariate that most closely aligned with the aims of our study and our knowledge of the study area (Murphy et al., Reference Murphy, Goodman, Farris, Karpanty, Andrianjakarivelo and Kelly2017). The final covariates included in our occupancy models were proximity to the road, Reserve and river, tree cover and slope (Supplementary Table 1). Slope was only used to model detection probability and the river to model occupancy, and the other covariates were used for both. Human population density and distance to the Reserve were highly correlated (r = 0.9). Covariates were standardized before modelling (Reilly et al., Reference Reilly, Tobler, Sonderegger and Beier2017).
We followed a systematic model selection approach, in which all combinations of covariates were modelled, following Murphy et al. (Reference Murphy, Goodman, Farris, Karpanty, Andrianjakarivelo and Kelly2017). Model selection was based on the second-order Akaike information criterion (AICc), including adjustment for overdispersion if the dispersion parameter ĉ was > 1 (QAICc; Burnham & Anderson, Reference Burnham and Anderson2002; MacKenzie & Bailey, Reference MacKenzie and Bailey2004). We excluded models that did not converge or produced estimates of p < 0.15 and ψ = 1 (MacKenzie et al., Reference MacKenzie, Nichols, Lachman, Droege, Royle and Langtimm2002). The top-ranked models were those with Δ(Q)AICc < 2 (MacKenzie et al., Reference MacKenzie, Nichols, Royle, Pollock, Bailey and Hines2006). Where there was more than one top-ranked model, we conducted model-averaging with the AICcmodavg package in R (Burnham & Anderson, Reference Burnham and Anderson2002; Mazerolle, Reference Mazerolle2017). The weight of evidence for each covariate was calculated by summing the AIC weights across candidate models containing that covariate (Burnham & Anderson, Reference Burnham and Anderson2002; Santos et al., Reference Santos, Chiarello, Ribeiro, Ribeiro and Paglia2016).
Results
Species richness and composition
Three memory cards malfunctioned in the field and did not provide any data, and another camera only worked for 1 day and therefore was excluded. The remaining 26 cameras took 16,050 photographs over 1,730 trap-days. The mammal species accumulation curve nearly reached a plateau, indicating that we detected most of the mammals within our survey area, except perhaps the most elusive or rare species, such as the leopard Panthera pardus (Bruce et al., Reference Bruce, Ndjassi, Fowler, Ndimbe, Fankem and Mbodbda2018c; Fig. 2). Thirty-one species were detected: six birds, one monitor lizard and 24 mammals, including six mammals of conservation concern (Near Threatened bay duiker Cephalophus dorsalis, yellow-backed duiker Cephalophus silvicultor and putty-nosed monkey Cercopithecus nictitans, Endangered tree pangolin Phataginus tricuspis and chimpanzee, and Critically Endangered western lowland gorilla) and one galago that could not be identified to species. We identified four large mammals (> 25 kg), 16 medium-sized mammals (1–25 kg) and three small mammals (< 1 kg) to species. We did not include birds or reptiles in our occupancy analyses as they were not the targets of our survey.

Fig. 2 Species accumulation curve for mammals identified by camera trapping during August–November 2017 in the community forest and surrounding habitat close to the Dja Biosphere Reserve, Cameroon (Fig. 1). The shaded ribbon indicates the 95% confidence interval.
Gorillas were recorded rarely (eight photographs), resulting in insufficient data for occupancy analysis. Putty-nosed monkey and moustached guenon Cercopithecus cephus were also recorded rarely, but as these primates are arboreal, occupancy analysis based on detections of these species from ground-level camera traps would not have been appropriate. The species recorded the most were blue duiker Philantomba monticola, Peter's duiker Cephalophus callipygus, agile mangabey Cercocebus agilis and brush-tailed porcupine Atherurus africanus (Table 1). We did not detect Critically Endangered forest elephants, large felids, Endangered giant ground pangolin Smutsia gigantea or the largest ungulates predicted to occur in the area (Supplementary Table 2). Thirty per cent of the medium-sized and large species we detected were listed in the IUCN Red List as Near Threatened or in one of the threatened categories, as were 50% (eight species) of those that we did not detect.
Table 1 Mammal species identified by camera trapping in the community forest and adjacent habitat surrounding the study village in the buffer zone of the Dja Biosphere Reserve in Cameroon (Fig. 1), with their Red List status (IUCN, Reference IUCN2021), trophic guild, body size, and naïve occupancy (the latter calculated as the number of cameras where the species was found divided by the total number of cameras).

1 LC, Least Concern; NT, Near Threatened; EN, Endangered; CR, Critically Endangered.
2 Small, < 1 kg; medium 1–25 kg; large > 25 kg.
Occupancy analysis
Occupancy models could be fitted for 18 mammals. For these species, estimated occupancy ranged from 0.92 ± SE 0.07 for blue duiker, to 0.35 ± SE 0.10 for Lady Burton's rope squirrel Funisciurus Isabella (Table 2). Detection probability ranged from 0.46 ± SE 0.05 for Lady Burton's rope squirrel, to 0.02 ± SE 0.01 for yellow-backed duiker. The occupancy of all species of conservation concern for which occupancy analysis was possible was > 0.5, but their detection probabilities were low (≤ 0.15).
Table 2 Estimates of occupancy (ψ) and detection probability (p) for mammals identified by camera trapping in the community forestand adjacent habitat surrounding the study village, from the top-ranked model or the average of the top ranked models. The effect of each covariate on ψ and p is shown by symbols if it occurs in the top ranked model or models: + is a positive effect and – is a negative effect. Parentheses indicate significant effects (P < 0.05).

Bates' pygmy antelope Neotragus batesi and agile mangabey were the only species for which the null model (no covariates included) was the top-ranked model. Proximity to the river was the most common predictor of occupancy. Tree cover was negatively correlated with occupancy for chimpanzees, implying that chimpanzee occupancy increased in cropland. Proximity to the road did not have a negative influence on occupancy for any species, and had a positive influence for Peter's duiker. Proximity to the Reserve had a positive effect on the occupancy of three rodents (brush-tailed porcupine, giant squirrel Protoxerus stangeri and giant pouched rat Cricetomys emini) and a negative effect on the occupancy of bay duiker and fire-footed rope squirrel Funisciurus pyrropus. For model selection details, model outputs and (Q)AICcWt information, see Supplementary Tables 3–5.
Discussion
The Dja region in Cameroon is important for mammals (Bruce et al., Reference Bruce, Amin, Wacher, Fankem, Ndjassi and Bata2018a), but there is increasing human activity and pressure on wildlife, and more surveys are needed to evaluate the status of mammal populations. We conducted a camera-trap survey in a previously unsurveyed community forest and adjacent habitat surrounding a village to the south-east of the Dja Biosphere Reserve and identified 24 mammal species, including six with an IUCN Red List status of Near Threatened or threatened: two medium-sized and large duikers (bay and yellow-backed), putty-nosed monkey, tree pangolin, chimpanzee and western lowland gorilla. Four of these species of conservation concern used > 50% of the study area, albeit with low detection probability (≤ 0.15), suggesting they are relatively widespread, despite the presence of a human settlement, road and persistent hunting. Our results highlight the conservation value of the community forest and support previous findings that the Dja Biosphere Reserve landscape, including the core and buffer zone, remains important for Central African mammals (Bruce et al., Reference Bruce, Ndjassi, Fowler, Ndimbe, Fankem and Mbodbda2018c; Lhoest et al., Reference Lhoest, Fonteyn, Daïnou, Delbeke, Doucet and Dufrêne2020).
The species richness in our study area is less than estimates from within the Dja Biosphere Reserve of 26 (Northern Sector) and 31 (Southern Sector) medium-sized and large species (Bruce et al., Reference Bruce, Amin, Wacher, Fankem, Ndjassi and Bata2018a), consistent with the assumption that human activity within and around the village and community forest is greater than inside the Reserve. In cases of severe overexploitation, with hunters optimizing their catch value (Alvard, Reference Alvard1998), only highly productive, generalist species remain, such as giant pouched rats and blue duikers (Abernethy et al., Reference Abernethy, Coad, Taylor, Lee and Maisels2013; Rosin & Poulsen, Reference Rosin and Poulsen2016). However, some medium-sized and large species were detected in the community forest, suggesting it is not a completely empty forest sensu Redford (Reference Redford1992). However, we did not detect most of the large species, including forest elephant, bongo Tragelaphus eurycerus, large felids and the giant ground pangolin, and the community was dominated by smaller species. Although we do not have conclusive evidence that the larger species are absent, our findings are similar to other studies that have documented a reduced mammal species richness and dominance of smaller species around villages and community forests in Central Africa, characteristic of local wildlife depletion (Beirne et al., Reference Beirne, Meier, Mbele, Menie, Froese, Okouyi and Poulsen2019; Lhoest et al., Reference Lhoest, Fonteyn, Daïnou, Delbeke, Doucet and Dufrêne2020).
The numbers of great apes usually decline rapidly under intensive hunting (Walsh et al., Reference Walsh, Abernethy, Bermejo, Beyers, De Wachter and Akou2003); the presence of both the gorilla and chimpanzee is therefore a positive sign for conservation within the community forest. Populations of both have declined in the Dja Biosphere Reserve (Bruce et al., Reference Bruce, Ndjassi, Fowler, Ndimbe, Fankem and Mbodbda2018c; UNESCO, 2019), and neither were identified in the community forests surveyed by Lhoest et al. (Reference Lhoest, Fonteyn, Daïnou, Delbeke, Doucet and Dufrêne2020). However, the low number of records of gorillas suggests they may be rare or present at low density. Chimpanzees appear to be well distributed throughout most of the study area, although the occupancy estimate is imprecise, and we found no relationship between occupancy and proximity to the logging road, suggesting some tolerance of disturbance (Vanthomme et al., Reference Vanthomme, Kolowski, Korte and Alonso2013). We also found a slightly positive correlation between chimpanzee occupancy and cropland, possibly reflecting an attraction to crops in the study area (Arlet & Molleman, Reference Arlet and Molleman2010).
Medium-sized duikers responded positively to human activity, with greater occupancy near the road (Peter's duiker), closer to the village and in areas with greater human population density (bay duiker). This could result from an increase in favourable resources. Clark et al. (Reference Clark, Poulsen, Malonga, Elkan and r2009) and Vanthomme et al. (Reference Vanthomme, Kolowski, Korte and Alonso2013) similarly found a positive correlation between these duikers and disturbance from roads, logging and settlements. Peter's duikers may be able to withstand hunting as a result of a higher maximum sustainable yield (van Vliet & Nasi, Reference van Vliet and Nasi2008), but bay duikers are less resilient (van Vliet et al., Reference van Vliet, Nasi, Emmons, Feer, Mbazza and Bourgarel2007) and could be more susceptible to unsustainable hunting in the community forest.
The most commonly hunted bushmeat species (blue duiker, brush-tailed porcupine and giant pouched rat; Fa et al., Reference Fa, Seymour, Dupain, Amin, Albrechtsen and Macdonald2006) had high estimates of occupancy and detection probability, indicating generally high abundance because, all else being equal, detection probability increases with abundance (MacKenzie et al., Reference MacKenzie, Nichols, Royle, Pollock, Bailey and Hines2006). Their high detection probability could also be linked to our survey protocol, as these species tend to use the trails along which cameras were placed, and similarly some species that were not detected may be less likely to use these trails or may require more specific camera placements (e.g. Bruce et al., Reference Bruce, Kamta, Mbobda, Kanto, Djibrilla and Moses2018b). Nevertheless, occupancy increased towards the Reserve for brush-tailed porcupines and giant pouched rats, suggesting that hunting of these species could be depleting populations close to the village. Blue duikers appear to be unaffected by this, as their distribution does not show the same pattern, potentially reflecting their ability to cope with hunting (van Vliet & Nasi, Reference van Vliet and Nasi2008; Ehlers Smith et al., Reference Ehlers Smith, Ehlers Smith, Ramesh and Downs2018).
Although we cannot tell from one survey what role the community forest has played in shaping the mammal community, its presence may have increased awareness about the need for hunting sustainably (Ngang, Reference Ngang2015). However, several other factors could also explain why our study area remains important for mammals. The location of the village close to the protected area allows movement of individuals from the Reserve to the community forest (Dupain et al., Reference Dupain, Guislain, Nguenang, De Vleeschouwer and Van Elsacker2004). Additionally, the village is remote and, before the logging road was built, it was very isolated, limiting access to bushmeat markets and perhaps reducing the incentive to hunt for income, until recently. The improved market access provided by the road may have led to increased hunting offtake (Ziegler et al., Reference Ziegler, Fa, Wohlfart, Streit, Jacob and Wegmann2016) and increased threat to the species we detected. However, as there are no previous survey data, it is vital that further surveys are conducted.
Many of the mammal species found in the Dja region are threatened or have declining populations (IUCN, Reference IUCN2021) and would benefit from systematic monitoring. Half of the medium-sized and large species we did not detect have an IUCN Red List status of Near Threatened or threatened, and although we detected Critically Endangered gorillas, we did not obtain a sufficient number of photographs to model their occupancy. For the threatened species for which we could model occupancy, our occupancy estimates had large standard errors as a result of low detection probabilities. Our findings highlight the difficulty of monitoring threatened mammals, especially in forest habitats (Wilcox et al., Reference Wilcox, Nash, Trageser, Kim, Hywood and Connelly2019). However, only with continued monitoring efforts over time and measurements of offtake will it be possible to identify species population trends and estimate hunting sustainability (Ntube et al., Reference Ntube, Ondoa, Mba, Fawoh, Mojoko, Tekwe and Acworth2002; Milner-Gulland & Rowcliffe, Reference Milner-Gulland and Rowcliffe2007), which are essential for species conservation and sustainable forest management within the community forest.
Acknowledgements
We thank Thomas Bruce for his help with species identifications, The European Space Agency Climate Change Initiative Land Cover project for data, and the anonymous reviewers for their feedback and suggestions. This research was carried out as part of the corresponding author's master's degree at Imperial College London and the open access article processing charge was covered by the Imperial Open Access Fund. This research received no other specific grant from any funding agency, or commercial or not-for-profit sectors.
Author contributions
Survey design: SB; fieldwork: SB, FK, CTKT; data analysis, writing: SJT; guidance, editing: SB, JMR.
Conflicts of interest
None.
Ethical standards
This research abided by the Oryx guidelines on ethical standards. Free informed consent was obtained from the village chief and inhabitants of the study village to conduct this research, and they were aware they could withdraw their consent at any time. Any images of people obtained by the camera traps were deleted. The village name is not presented to ensure anonymity. This research was approved by Oxford University's Central University Research Ethics Committee (R45771/RE001).





 Open access
Open access