


Epidemiological studies have shown that diets rich in whole grain may protect against CVD, type 2 diabetes and certain types of cancer( Reference von Ruesten, Feller and Bergmann 1 , Reference Grooms, Ommerborn and Pham 2 ). Recent prospective studies also indicate a lowered total mortality and CVD mortality( Reference Wu, Flint and Qi 3 ). Despite the accumulating epidemiological evidence, the mechanisms involved in the protective effects of whole grains remain unclear, and controlled intervention studies indicate variable effects of whole-grain v. refined-grain diets( Reference Harris Jackson, West and Vanden Heuvel 4 ). Consequently, there is a need to elucidate the causal relationship between dietary fibre (DF) characteristics and health outcome. In addition to the DF per se, the phytonutrients associated with DF may also contribute to this factor. Further, the structural features and degree of processing – for example, grinding – may affect the resistant starch (RS) contents that add to the total DF complex present in whole-grain food.
Whole-grain diets have been suggested to ameliorate blood (b) glucose control( Reference Harris Jackson, West and Vanden Heuvel 4 ). Several mechanisms have been put forward – for example, acute effects, by reducing the rate of upper-gut digestion and absorption of starch, or semi-acute and longer-term effects, originating from gut fermentation of DF. In this respect, SCFA, formed during gut fermentation, have been proposed to be involved( Reference Cani, Everard and Duparc 5 , Reference Tolhurst, Heffron and Lam 6 ).
Gut-derived hormones play an important role in energy regulation and are proposed to be involved as modulators of inflammation as well. Glucagon-like peptide (GLP)-1 and peptide YY (PYY) participate in the regulation of appetite and glucose metabolism( Reference Sam, Troke and Tan 7 – Reference Persaud and Bewick 9 ). The gut hormone GLP-2 appears to be involved as a regulator of gut barrier function( Reference Cani, Possemiers and Van de Wiele 10 ), and it has been suggested that inflammatory processes linked to diminished gut barrier function with concomitant systemic influx of lipopolysaccharides (LPS) may promote development of obesity and diabetes( Reference Delzenne, Neyrinck and Cani 11 ). GLP-1, PYY and GLP-2 are secreted from L-cells present throughout the intestinal epithelium with highest density towards the distal ileum and colon and are co-released in the presence of carbohydrates( Reference Sam, Troke and Tan 7 , Reference Schwartz and Holst 12 ). Studies are at hand implicating that certain gut substrates may affect L-cells and stimulate gut hormones through a mechanism linked to gut fermentation. SCFA receptors co-localised with enteroendocrine L-cells appear to provide a link between fermentation and stimulation of gut hormones, and SCFA receptor deficiency impairs SCFA-induced gut hormone secretion both in vitro and in vivo ( Reference Psichas, Sleeth and Murphy 13 ). In rats, dietary supplementation with oligofructose increased the number of L-cells in the proximal colon, as well as the circulating GLP-1 concentration, compared with a control diet( Reference Cani, Hoste and Guiot 14 ). Interestingly, a 2-week dietary supplementation with oligofructose-enriched inulin increased p-GLP-1 and PYY concentrations in healthy adults( Reference Cani, Lecourt and Dewulf 15 ). Further, intrinsic DF present in evening meals of barley kernels or brown beans facilitated b-glucose regulation and beneficially affected gut hormones such as GLP-1 or PYY as well as subjective appetite at a subsequent breakfast meal in young healthy subjects( Reference Nilsson, Östman and Holst 16 – Reference Nilsson, Johansson and Ekstrom 18 ). The important role of gut hormones in systemic metabolism and inflammation suggests that an increased endogenous release of these hormones would be of importance with respect to prevention of cardiometabolic disorders and obesity.
The purpose of the present study was to investigate the role of indigestible carbohydrates (NSP and RS) present in barley kernel-based bread (BB) on metabolism and gut hormones in a cohort of healthy, middle-aged subjects. The intervention with BB or white wheat bread (WWB) reference was performed for 3 consecutive days before the overnight study, where a standardised breakfast was provided on day 4 and blood samples were collected in the postprandial phase (0–150 min). Blood parameters included measures of glucose metabolism (glucose and insulin) and a range of gut-derived hormones (GLP-1, GLP-2, PYY, oxyntomodulin (OXM)), ghrelin, inflammatory markers (IL-6, IL-18 and adiponectin) and NEFA. In addition, s-SCFA was determined, and subjective appetite sensations were registered. Breath H2 was measured as an indicator of gut fermentative activity.
Methods
Test subjects
A total of twenty healthy volunteers, three men and seventeen women, aged 64·1 (sd 5·9) years, with normal BMI (mean 23·6 (sd 2·3 kg/m2)) participated in this study. Recruitments of test subjects were conducted through advertisements in local newspapers during September 2010 and January 2011. The experimental study was conducted at Lund University, and was completed in March 2011. The inclusion criteria were as follows: men and women between 50 and 70 years of age, normal to slightly overweight (BMI 19–28 kg/m2), fasting plasma glucose ≤6·1 mmol/l, non-smoker, overall healthy and with no known metabolic disorders or food allergies. Subjects who took anti-hypertensive medications (one subject) and prescription-free painkillers without anti-inflammatory action were also included. All the enrolled test subjects completed the study. This study was conducted according to the guidelines laid down in the Declaration of Helsinki, and all the procedures involving human subjects were approved by the Regional Ethical Review Board in Lund, Sweden (Reference 2010/457). Written informed consent was obtained from all the subjects. The study was registered at ClinicalTrials.gov (NCT01718431).
Experimental procedure
The test subjects participated in two 3-d dietary interventions, with either test products or reference products taken in a random order separated by at least 2 weeks (between 2·0 and 4·5 weeks, mean 2·7 (sem 0·8) weeks). A test product consisting of BB and a reference product consisting of WWB were included.
Intervention protocol
The quantity of the test products and reference products to be ingested daily during the 3-d study period was standardised to provide 100 g of potentially available starch per day, calculated as the difference between total starch and RS. This corresponded to 9·1 g total DF/d (72 % NSP and 28 % RS) in the case of the reference WWB intervention and 37·6 g total DF/d (55·8 % NSP and 45·2 % RS) in the case of the BB test intervention period. The daily intake of test bread or reference bread was distributed into three equal portions to be consumed at approximately 08.00, 14.00 and 21.00 hours for the first 2 d. On day 3, half of the daily intake (50 g available starch) was distributed equally between the 08.00 and 14.00 hours meals, and the other half (50 g available starch) was consumed at 21.00 hours in the evening.
The subjects were encouraged to standardise their meal pattern and to avoid alcohol, excessive physical exercise or foods rich in DF during the 3-d intervention periods. Furthermore, they should not have taken antibiotics or probiotics during the previous 2 weeks and also throughout the study period. After the evening meal (consisting of the last portion of the test product or reference product, respectively), on day 3, the subjects were fasting until the standardised breakfast was served at the experimental department. The subjects arrived at the experimental department at 07.30 hours. An intravenous cannula (BD Venflon; Becton Dickinson) was inserted into an antecubital vein to be used for blood sampling. Fasting blood samples were collected and subjective appetite and breath H2 were registered before providing the standardised breakfast. The standardised breakfast (see below) was provided at approximately 08.00 hours and was ingested within 13 min. During the 2·5 h of repeated sampling, the subjects were told to maintain low physical activity.
Standardised breakfast on day 4
The standardised breakfast consisted of 122·9 g WWB corresponding to 50 g of available carbohydrates, analysed according to Holm et al.( Reference Holm, Björck and Drews 19 ), and 2·5 dl tap water.
Recipes and preparation of reference and test products as well as the standardised breakfast white wheat bread (white wheat bread: reference product and standardised breakfast)
The WWB was baked according to a standardised procedure in a home baking machine (Tefal Home Bread Model no. 573102; menu choice, programme 2 (white bread, 1000 g, quick (time 2·32 h))). The bread was made from 540 g of white wheat flour (Kungsörnen AB), 360 g water, 4·8 g dry yeast and 4·8 g NaCl (without iodine). After cooling, the bread was sliced and wrapped in aluminium foil in portion sizes, placed in plastic bags and stored in a freezer (–20°C). The crust was removed from the WWB intended for the standardised breakfast.
Barley kernel-based bread (test product)
A total of 595 g barley kernels (Finax) were boiled in 520 g water for 12 min and then cooled for 30 min at ambient room temperature. Water was completely absorbed into the kernels when cooked. Added to the kernels were 105 g wheat flour, 6 g dry yeast, 5 g salt and 300 g water. The dough was kneaded for 4 min (Electrolux AKM 3000, N23 N25) and proofed for 30 min in a bowl, followed by another proofing (35 min) in a baking tin. The baking tin was covered with aluminium foil and baked in a household oven at 225°C until the inner temperature of the bread reached 96°C with a pan of water present to maximise steam. After baking, the bread was cooled at ambient room temperature, removed from the tin and wrapped in wet towels. After cooling, the towels were removed and the bread samples were placed in plastic bags at room temperature overnight. The day after, the bread samples were sliced and wrapped in aluminum foil in portion sizes, placed in plastic bags and stored in a freezer (–20°C). On days before consumption, the test persons were instructed to take a daily portion of bread from the freezer and thaw it at ambient temperature, with the bread still wrapped in aluminum foil and maintained in the plastic bag.
Chemical analysis of the test and reference products
The test and reference products were analysed with respect to total starch( Reference Björck and Siljeström 20 ), RS( Reference Åkerberg, Liljeberg and Granfeldt 21 ) and insoluble and soluble NSP( Reference Asp, Johansson and Hallmer 22 ). Before analysis of total starch and NSP, the bread samples were air-dried and milled. RS was analysed on products as eaten. Available starch was calculated by subtracting RS from total starch. The composition of the test and reference products is presented in Table 1.
Table 1 Portion size and carbohydrate composition of the reference and test products, respectivelyFootnote *

RS, resistant starch; DF, dietary fibre; WWB, white wheat bread; BB, barley kernel-based bread.
* Available starch is calculated as the difference between total starch( Reference Björck and Siljeström 20 ) and RS( Reference Åkerberg, Liljeberg and Granfeldt 21 ). Values of total starch are based on means of two replicates, RS means of six replicates and NSP are based on means of three replicates. Included in total DF are RS and insoluble and soluble NSP, determined gravimetrically according to Asp et al.( Reference Asp, Johansson and Hallmer 22 ).
Sampling and analysis of physiological variables
Venous blood samples were taken to determine serum (s-) insulin, s-NEFA, s-SCFA, s-IL-6, s-IL-18 and s-adiponectin and for the determination of plasma (p-) GLP-1, p-GLP-2, p-PYY, p-OXM and p-ghrelin. Finger-prick capillary blood samples were taken for the determination of whole b-glucose concentrations (HemoCue®B-glucose; HemoCue AB). Breath H2 was measured as an indicator of colonic fermentation using a Gastro+ (Bedfont EC60 Gastrolyzer; Bedfont). Measures of subjective appetite sensations (satiety, hunger and desire to eat) were obtained using a 100-mm visual analogue scale. Glucose, H2 and appetite sensations were determined at fasting and at 15, 30, 45, 60, 90, 120 and 150 min after commencing the standardised breakfast. Insulin level was determined at similar time points, with the exception of 15 min. GLP-1 was analysed at fasting and at 30, 45, 60, 90 and 150 min, and GLP-2 and ghrelin were analysed at fasting and at 30, 60, 90 and 150 min. IL-6, IL-18, OXM and PYY were determined at fasting and at 60 and 120 min after start of the breakfast. Adiponectin level was analysed at fasting and at 120 min, and NEFA was determined at fasting and at 150 min. SCFA was determined at fasting.
Serum and plasma were separated by centrifugation and immediately stored in a freezer (–40ºC) until analysis. Blood-collecting tubes intended for the analysis of p-ghrelin, p-GLP-1, p-GLP-2, p-PYY and p-OXM were prepared using an inhibition cocktail consisting of a dipeptidyl peptidase-4 (DPPIV) inhibitor (10 µl/ml blood) (Millipore) and Trasylol® 10 000 KIE/ml aprotinin (50 µl/ml blood) (Bayer HealthCare AG) before blood sampling. Tubes containing the inhibition cocktail were kept on ice until use for a maximum of 6 d. Plasma for the analysis of ghrelin was additionally treated with 1 m-HCl before freezing (10:1).
Serum insulin levels were measured using an enzyme immunoassay kit procedure (Mercodia) and performed on an integrated immunoassay analyser (CODA Open Microplate System; Bio-Rad Laboratories). Commercial kits based on ELISA were used for the determination of s-adiponectin (Mercodia Adiponectin ELISA; Mercodia), p-OXM (Cusabio Biotech), p-PYY (3-36 and 1-36), p-GLP-2 (Yanaihara Institute Inc.), p-GLP-1 (active 7-36) (Alpco Diagnostics), p-ghrelin (BioVendor GmbH), s-IL-6 (R&D Systems) and s-IL-18 (MBL). Serum NEFA levels were measured using an enzymatic colorimetric method (NEFA C, ACS-ACOD method; Wako Chemicals GmbH), and s-SCFA (acetate, propionate and butyrate) were determined using a GC method( Reference Brighenti 23 ).
Calculations and statistical methods
Statistical evaluations of glucose and insulin areas and peaks were based on incremental changes from fasting concentrations. Calculations regarding all other test variables were performed using actual values. The incremental AUC (iAUC) and AUC were calculated for each subject and the respective breakfast meal, using the trapezoid model. GraphPad Prism (version 5; GraphPad Software) was used for graph plotting and calculation of areas. Incremental peak (iPeak) concentrations were determined for glucose and insulin as individual maximum postprandial increase from baseline. Individual mean values during the experimental day were calculated for physiological test variables when a postprandial response after breakfast was scarcely seen. Total SCFA were calculated as the sum of the concentrations of acetate, propionate and butyrate. As a measure of insulin resistance, a homoeostatic model assessment (HOMA-IR) was used (fasting glucose (mmol/l)×fasting insulin (mU/l)/22·5)(
Reference Matthews, Hosker and Rudenski
24
). For the assessment of insulin sensitivity, a modified composite insulin sensitivity index (ISIcomposite), also called the Matsuda index, was used, including measures of b-glucose and s-insulin both at fasting and in the postprandial phase after the standardised breakfast
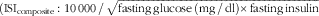 $$({\rm ISI}_{{{\rm composite}}} \,\colon\,{\rm 1}0\,000\,/\,\sqrt {{\rm fasting}\,{\rm glucose}\,\left( {{\rm mg}\,/\,{\rm dl}} \right){\times}{\rm fasting}\,{\rm insulin}$$
$$({\rm ISI}_{{{\rm composite}}} \,\colon\,{\rm 1}0\,000\,/\,\sqrt {{\rm fasting}\,{\rm glucose}\,\left( {{\rm mg}\,/\,{\rm dl}} \right){\times}{\rm fasting}\,{\rm insulin}$$
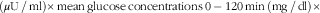 $$({\rm \mu U}\,/\,{\rm ml}){\times}{\rm mean}\,{\rm glucose}\,{\rm concentrations}\,0-{\rm 12}0\,{\rm min}\,\left( {{\rm mg}\,/\,{\rm dl}} \right){\times}$$
$$({\rm \mu U}\,/\,{\rm ml}){\times}{\rm mean}\,{\rm glucose}\,{\rm concentrations}\,0-{\rm 12}0\,{\rm min}\,\left( {{\rm mg}\,/\,{\rm dl}} \right){\times}$$
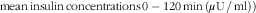 $${\rm mean}\,{\rm insulin}\,{\rm concentrations}\,0-{\rm 12}0\,{\rm min}\,({\rm \mu U}\,/\,{\rm ml}))} } $$
(
Reference Matsuda and DeFronzo
25
,
Reference DeFronzo and Matsuda
26
). The determination of the Matsuda indices was modified with respect to analysing postprandial concentrations for insulin and glucose after consuming standardised WWB based on 50 g rapidly available starch instead of 75 g glucose.
$${\rm mean}\,{\rm insulin}\,{\rm concentrations}\,0-{\rm 12}0\,{\rm min}\,({\rm \mu U}\,/\,{\rm ml}))} } $$
(
Reference Matsuda and DeFronzo
25
,
Reference DeFronzo and Matsuda
26
). The determination of the Matsuda indices was modified with respect to analysing postprandial concentrations for insulin and glucose after consuming standardised WWB based on 50 g rapidly available starch instead of 75 g glucose.
Significant differences in test variables after the test and reference products were assessed with ANOVA (general linear model) using Minitab Statistical Software (release 14–16, Minitab; Minitab Inc.). In the cases of unevenly distributed residuals (tested using the Anderson–Darling test and considered unevenly distributed when P<0·05), Box–Cox transformations were performed on the data before ANOVA. Differences between the products at different time points were evaluated using a mixed model (PROC MIXED in SAS release 9.2 and 9.3; SAS Institute Inc.) with repeated measures and an autoregressive covariance structure. Due to previous suggestions that SCFA may be involved in the release of gut hormones, relationships between total SCFA and gut hormones (PYY, GLP-1 and GLP-2) were investigated using Pearson’s correlation in Minitab Statistical Software (release 14–16). Randomisation of the order in which the test or reference products were taken and power calculations were performed in Minitab Statistical Software (release 14–16). If a value from a test subject was missing for one of the products, the test subject was excluded from that specific calculation. Values of P≤0·05 were considered significant. Data are expressed as mean values with their standard errors of the mean; n 20.
Power calculation
The primary outcome measure for power calculations was change in b-glucose iAUC 0–120 min after the standardised breakfast. Number of test subjects required for the study was determined in Minitab, using previous results of ‘overnight’ effects of barley kernel meals on glucose iAUC 0–120 min( Reference Nilsson, Östman and Holst 16 ). Assuming a difference of 70 mmol/l per min between the WWB and the BB intervention, and an sd of 82 mmol/l per min, with α=0·05 and 1–β=0·8, about thirteen to seventeen test subjects were required (two-tailed test). We decided to increase the number of test subjects to twenty due to the lack of information from previous similar studies regarding the cohort included in the present study (middle-aged adults).
Results
Gastrointestinal hormones (peptide YY, glucagon-like peptide-1, glucagon-like peptide-2, oxyntomodulin and ghrelin)
A main effect of the test products was observed on p-PYY concentrations in the morning following the 3-d intervention, revealing higher levels following BB compared with WWB (P=0·042). When calculating a mean of p-PYY for the time period 0–150 min, increased concentrations were seen after BB (18 %; P=0·043) compared with WWB (Table 2). Consumption of BB for 3 d resulted in significantly increased concentrations of p-GLP-1 at fasting on day 4 (56 %; P=0·006). The mean p-GLP-2 concentrations were increased during the standardised breakfast on day 4 (13 %; P=0·042) after BB intake for 3 d, as compared with WWB, and a tendency (P=0·06) towards increased fasting concentrations of p-GLP-2 was observed after BB (Table 2).
Table 2 Gut hormone responses following 3 d of intervention with reference or test products, respectively (Mean values with their standard errors)

WWB, white wheat bread; BB, barley kernel-based bread; p, plasma; GLP-1 and 2, glucagon-like peptides 1 and 2; PYY, peptide YY; OXM, oxyntomodulin.
Significantly different from WWB:
* P < 0>05,
** P < 0>01 (ANOVA; general linear model, Minitab).
† Percentage of differences in concentrations of test variables after BB compared with WWB.
‡ P=0·06.
No significant differences were observed in the concentrations of p-ghrelin or p-OXM following the standardised breakfast, depending on intervention product (Table 2).
Breath hydrogen and serum SCFA
Inclusion of the BB test product resulted in increased breath H2 levels at fasting on day 4 compared with the WWB (146 %; P=0·007). The increased levels of H2 were maintained during the standardised breakfast (363 %; P=0·000) (Fig. 1; Table 3). Intake of BB for 3 d increased fasting concentrations of s-acetate and total s-SCFA (18 %; P=0·013 and 16 %; P=0·012, respectively, Table 3). A tendency (P=0·1) towards an increase in s-butyrate levels was observed after BB compared with WWB.

Fig. 1. Breath hydrogen excretion during the experimental day. Mean postprandial breath hydrogen concentrations at a standardised breakfast following 3 d of intervention with barley kernel-based bread (BB) or white wheat bread (WWB), respectively. Dotted lines at the y-axis indicate mean values for BB (32·5 (sem 6·77) parts per million (ppm)) and WWB (7·01 (sem 1·18) ppm). Values are means with their standard errors; n 20 (repeated measures; mixed model in SAS). ![]() , WWB;
, WWB; ![]() , BB.
, BB.
Table 3 Serum (s)-SCFA and breath hydrogen following 3 d of intervention with reference or test products, respectively (Mean values with their standard errors)

WWB, white wheat bread; BB, barley kernel-based bread; ppm, parts per million.
Significantly different from WWB:
* P < 0>05,
** P < 0>01,
*** P < 0>001 (ANOVA; general linear model, Minitab).
† Percentage of differences in concentrations of test variables after BB compared with WWB.
‡ P=0>1.
Blood glucose and serum insulin
B-glucose and s-insulin responses at the standardised breakfast are presented in Fig. 2. In comparison with the intervention with WWB, the 3-d intervention with BB beneficially affected the b-glucose response at the following standardised breakfast in terms of lower iPeak (BB and WWB: 2·8 (sem 0·2) and 3·4 (sem 0·2) mmol/l, respectively, –17 %; P=0·014) and lower iAUC 0–120 min (BB and WWB: 152 (sem 12) and 194 (sem 18) mmol/l per h, respectively, –22 %; P=0·018). In addition, in comparison with WWB, BB lowered the s-insulin iPeak (BB and WWB: 0·21 (sem 0·02) and 0·24 (sem 0·02) nmol/l per min, respectively, –11 %; P=0·024) and s-insulin iAUC 0–120 min (BB and WWB: 11·4 (sem 1·2) and 13·8 (sem) 1·3 nmol/l per min, respectively, –17 %; P=0·002). Further, the ISIcomposite was increased (25 %; P=0·005), indicating a higher insulin sensitivity on day 4 after 3 d of BB intervention compared with WWB. No effects were observed for fasting insulin resistance as estimated using HOMA-IR. Table 4 displays the results regarding insulin sensitivity variables and fasting glucose and insulin concentrations.

Fig. 2 Incremental blood (b)-glucose and serum (s)-insulin responses during the experimental day. Incremental b-glucose (a) and s-insulin (b) response at a standardised breakfast following 3 d of intervention with barley kernel-based bread (BB) or white wheat bread (WWB), respectively. Values are means with their standard errors; n 20 (repeated measures; mixed model in SAS). ![]() , WWB;
, WWB; ![]() , BB.
, BB.
Table 4 Whole blood (b)-glucose and serum (s)-insulin responses including measures of insulin resistance and insulin sensitivity at a standardised breakfast following 3 d of intervention with reference or test products, respectively (Mean values with their standard errors)

WWB, white wheat bread; BB, barley kernel-based bread; HOMA-IR, homoeostatic model assessment insulin resistance; ISIcomposite, insulin sensitivity index.
** Significantly different from WWB (P < 0>01; ANOVA; general linear model, Minitab).
† Percentage of differences in concentrations of test variables after BB Swap the † and ** footnotes.
Subjective appetite sensation, serum NEFA and markers of inflammation (serum-IL-6, serum-IL-18 and serum adiponectin)
No significant differences in appetite sensations (satiety, hunger and desire to eat) were observed at the standardised breakfast meal depending on previous interventions (data not shown). However, a tendency for a main effect appeared for desire to eat (P=0·08, repeated measures; mixed model in SAS), with less desire to eat in the case of the 3-d intervention period with BB.
No differences were observed in s-NEFA, s-IL-6, s-IL-18 or s-adiponectin levels depending on the intervention product (P>0·05, data not shown).
Relationships between markers of gut fermentation and gut hormones
In the morning on day 4 in both the intervention periods, the fasting concentrations of total s-SCFA were positively associated with fasting concentrations of p-PYY (r 0·51, P=0·038 and r 0·57, P=0·01, after WWB and BB, respectively). Fasting concentrations of s-SCFA were similarly associated with postprandial p-PYY concentrations (mean 0–150 min; r 0·51, P=0·032 and r 0·55, P=0·014, after WWB and BB, respectively). In addition, fasting s-SCFA after BB intervention was positively associated with postprandial (mean 0–150 min) concentrations of p-GLP-2 (r 0·53, P=0·020).
Discussion
The present study investigated the impact of indigestible carbohydrates (NSP and RS) present in BB on endogenous release of gut hormones, metabolic risk markers and perceived appetite in healthy, normal-to-slightly overweight middle-aged subjects. We showed that intake of BB for 3 d markedly increased gut fermentation activity, manifested as increased breath H2 excretion and s-SCFA concentrations at a standardised breakfast on day 4, as compared with intake of the WWB reference. Further, circulating concentrations of gut hormones p-GLP-1, p-GLP-2 and p-PYY were significantly elevated following consumption of BB for 3 d. In parallel, the present study demonstrates not only a lowered b-glucose response (iPeak and iAUC 0–120 min) at the standardised breakfast challenge after BB in comparison with the WWB, but also a reduced insulin response (iPeak and iAUC 0–120 min). Although no specific analysis of insulin sensitivity was performed using, for example, the clamp technique, the present study indicates improved insulin sensitivity as determined by an increased ISIcomposite after the BB intervention. The results are in accordance with studies showing improved insulin sensitivity in healthy subjects after a meal tolerance test in the morning, following intake of 60 g DF from Hi-maize over a period of 24 h the previous day( Reference Robertson, Currie and Morgan 27 ). The authors suggested a mechanism related to increased rate of colonic fermentation. Similarly, insulin resistance, assessed using the euglycemic–hyperinsulinemic clamp technique, was improved in healthy subjects after 4 weeks intervention with Hi-maize RS (Hi-maize 260; 30 g/d; National Starch and Chemical)( Reference Robertson, Bickerton and Dennis 28 ).
The underlying mechanisms are, however, not clarified, and other authors have, for example, argued that the beneficial effects of DF on insulin resistance are due to the interference of DF with the digestion and/or absorption of dietary proteins in the small intestine( Reference Weickert, Roden and Isken 29 ). Consequently, effects of DF on insulin sensitivity was previously investigated in obese subjects with the metabolic syndrome after 18 weeks of DF supplementation and compared with a diet high in proteins, or a diet rich in both DF and proteins, and a control diet neither enriched in proteins nor in DF( Reference Weickert, Roden and Isken 29 ). Insulin sensitivity was assessed using the euglycemic–hyperinsulinemic clamp technique. The results showed no improvement on insulin sensitivity after DF supplementation compared with the control diet. However, the high-protein diet impaired insulin sensitivity, resulting in significantly better insulin sensitivity after DF supplementation in comparison with the high-protein diet. In addition, when DF was added to the high-protein diet, the deleterious effect on insulin resistance was diminished. The DF supplement source in the obese study population( Reference Weickert, Roden and Isken 29 ) was composed of DF isolates consisting of non-soluble and poorly fermentable DF from oat husk, where the β-glucans were removed. This is in contrast with the test product included in the present study, which instead was rich in highly fermentable soluble DF made up mostly of β-glucans. The results, thus, suggest that several mechanisms may be involved in eliciting beneficial effects and that different sources of DF may target different metabolic pathways.
In humans, supplementation with different DF has shown different effects on second meal overnight GLP-1 concentrations. Consequently, in healthy subjects, 4-week supplementation with RS (30 g/d) did not affect GLP-1 concentrations at fasting or postprandially at breakfast the following day( Reference Robertson, Bickerton and Dennis 28 ), whereas an increase in GLP-1 was observed after 2 weeks of prebiotic supplementation containing oligofructose-enriched inulin (16 g/d)( Reference Cani, Lecourt and Dewulf 15 ). Further, 24-h supplementation with a mixture of insoluble DF from oat extracts (depleted from β-glucans), wheat DF extracts and RS (Hi-maize 1043; National Starch and Chemical) did not increase GLP-1 concentrations the following day at fasting or following a standardised breakfast( Reference Weickert, Mohlig and Koebnick 30 ). The differences in outcome on GLP-1 with different DF sources is not known but could be related to the differences in capacity to stimulate specific gut bacteria. The incretin GLP-1 has been ascribed both anti-diabetic and anti-obesity features, and during the last few years there has been an increased interest regarding the possibility to stimulate endogenous GLP-1 release as a means to improve postprandial glucose metabolism( Reference Mansour, Hosseini and Larijani 31 , Reference Holst and McGill 32 ). In rats, dietary supplementation with oligofructose increased the number of L-cells in the proximal colon with a concomitant increase in endogenous GLP-1 concentrations( Reference Cani, Hoste and Guiot 14 ). In addition, specific key differentiation factors, involved in endocrine cell maturation, were up-regulated after oligofructose treatment. In this context, it was suggested that the promotion of L-cell development was possibly mediated through SCFA formation( Reference Cani, Hoste and Guiot 14 , Reference Robertson, Bickerton and Dennis 28 ). It can be speculated that the increase in circulating gut hormones observed in the present study may be a result from a shift in the microbial flora and an increased number of L-cells after BB intervention. However, the intervention time in the present study was 3 d, and the regeneration time for enteroendocrine cells is usually 5–7 d( Reference Egerod, Engelstoft and Grunddal 33 ). Thus, probably additional mechanisms may be operative in the shorter time perspective of the present work, possibly involving increased activation of already-existing L-cells, and increased SCFA receptor signalling( Reference Kaji, Karaki and Kuwahara 34 ). Tolhurst et al.( Reference Tolhurst, Heffron and Lam 6 ) suggested that the colonic L-cells may be directly influenced by SCFA (acetate and propionate), with a concomitant stimulation of GLP-1 and PYY release. Accordingly, rectal administration of acetate in hyperinsulinaemic overweight subjects increased circulating GLP-1 and PYY 1 h after administration( Reference Freeland and Wolever 35 ). The possibility to activate the L-cells by such a mechanism is a plausible hypothesis behind the results presented in this study, supported by an increased level of circulating s-SCFA (total of acetate, propionate and butyrate) and increased concentrations of gut hormones – p-GLP-1, p-PYY and p-GLP-2 – all co-secreted from the L-cells( Reference Schwartz and Holst 12 ). The observed positive relationships between s-SCFA and p-PYY or p-GLP-2, respectively, further strengthen the possibility for such a mechanism. The results regarding effects of the specific indigestible carbohydrates present in the barley kernel product on release of gut hormones in a 11- to 14-h perspective, and the relationships between the gut hormones and colonic fermentation metabolites – that is, SCFA – are novel. However, studies regarding mechanisms of gut flora-mediated benefits on host metabolism are scarce and more studies are needed comparing efficacy of different DF as presented in foods, focusing on the relationships between gut fermentation and cardiometabolic benefits. A tentative mechanism for the metabolic benefits seen in the present study relates to the activation of specific gut bacteria by the intrinsic DF substrates in BB, causing stimulation of L-cells and/or generation of SCFA. Consequently, ingestion of BB for 3 d increased abundance of specific gut microbes in faeces, providing evidence for a mechanism related to gut fermentation (P Kovatcheva-Datchary, A Nilsson, R Akrami, Y Shiuan Lee, F De Vadder, T Arora, A Hallen, E Martens, I Björck and F Bäckhed, unpublished results).
This study is to our knowledge among the first to observe a simultaneous increase in several gut hormones in humans in response to intrinsic DF in a composite food. As a particularly interesting observation, there was an increase in p-GLP-2 concentrations after the intervention with BB. To our knowledge, only one report is available showing a similar increase in endogenous GLP-2 following intake of brown beans using a similar experimental design( Reference Nilsson, Johansson and Ekstrom 18 ). GLP-2 appears to be involved in epithelial cell proliferation and intestinal growth, and its importance for gut barrier functions has been increasingly acknowledged( Reference Orskov, Hartmann and Poulsen 36 ). LPS, released by unfavourable intestinal bacteria and absorbed into the systemic circulation, may promote a state of metabolic endotoxemia, and act to trigger sub-clinical inflammation and development of insulin resistance( Reference Cani, Amar and Iglesias 37 ), and even obesity( Reference Everard and Cani 38 ). Therefore, the gut epithelium provides an important protective barrier against translocation of endotoxins( Reference Cani 39 ). Interestingly, prebiotic supplementation increased circulating concentrations of GLP-2, as well as decreased intestinal permeability and concentrations of p-LPS in obese mice, altogether implicating a GLP-2-dependent mechanism( Reference Cani, Possemiers and Van de Wiele 10 ).
The present study has certain limitations. For example, instead of using the golden standard method to determine insulin resistance – that is, the hyperinsulinemic–euglycemic clamp – HOMA-IR, and the Matsuda index were used to estimate insulin sensitivity. Matsuda index has been shown to correlate well (r 0·73, P<0·0001) with the rate of whole-body glucose disposal during the euglycemic insulin clamp( Reference Matsuda and DeFronzo 25 ). In our study, we used a modified method for calculations of the Matsuda index. Typically, the postprandial determinations of glucose and insulin concentrations are performed after a 75 g glucose load. We instead assessed the Matsuda index after a standardised WWB breakfast, yielding 50 g available carbohydrates from starch. As a potential additional study limitation it should be noted that the relationship between s-SCFA and p-GLP-2 was obtained between fasting concentrations of p-SCFA and mean concentrations of p-GLP-2 during the intervention period (0–150 min), and it cannot be ruled out that the discrepancy in the time may have affected the results. Regarding test subjects, an unbalanced sex distribution was present, providing only three male compared with seventeen female subjects. We did not investigate all the data only for women; however, re-calculating glucose and insulin data without men did not affect the outcome. Another possible study limitation was that the study design (food products consumed at home) did not allow for controlling compliance. However, the apparent difference in breath H2 at day 4 between test BB and reference WWB intervention indicates that the test subjects followed instructions regarding the test products. Finally, several response variables were investigated, and we cannot rule out that some of the detected associations were due to chance. However, we did not adjust for multiple testing as several of the investigated response variables were correlated; such adjustment would, therefore, be unduly conservative.
Conclusions
The present study demonstrates the metabolic benefits of ingestion of BB for 3 d in healthy middle-aged subjects, as manifested by increased concentrations of GLP-1, PYY and GLP-2, facilitated b-glucose regulation and improved insulin sensitivity. In addition, positive relationships were observed between total SCFA and gut hormones PYY and GLP-2, indicating a causal relationship between SCFA production and gut hormone secretion. These new findings suggest anti-inflammatory, anti-diabetic and anti-obesogenic potential of foods rich in intrinsic indigestible carbohydrates (NSP/RS approximately 13/11 %, DM), as present in the BB. It is suggested that the mechanisms are related to gut fermentation of the DF fraction in BB. These findings are novel and encouraging, and it is put forward that such foods may add to the preventive value of a healthy diet. However, more studies are needed to clarify the complex mechanisms underlying the relationships between colonic fermentation of certain DF and cardiometabolic benefits.
Acknowledgements
This study was funded by the Lund University Antidiabetic Food Centre, a VINNOVA VINN Excellence Centre (grant no. 2013/46).
The authors’ contributions were as follows: E. V. J.-B., I. M. E. B. and A. C. N. designed the study; E. V. J.-B. and A. C. N. conducted the study; E. V. J.-B. and A. C. N. analysed the data; E. V. J.-B, I. M. E. B. and A. C. N. evaluated the results and wrote the paper; and A. C. N. and I. M. E. B. had primary responsibility for the final content. All the authors read and approved the final manuscript.
The authors declare that they have no conflicts of interest.






