



The culprits, echinoderms, amphibians and mammals
Over recent decades, in addition to the echinoderms, two other groups, the amphibians and the mammals, have been used as models to study polyspermy prevention. The preparation of oocytes from these three groups, often for the convenience of the operator, has led to a series of artefacts that have altered the kinetics of this cell–cell interaction, leading to a misconception of what is really happening at fertilization. These include, not taking into account the cytoplasmic maturity of the oocytes, removing the extracellular coats from the oocytes, changing the physical–chemical nature of the egg surface and using unphysiological numbers of spermatozoa. An alternative explanation why oocytes are monospermic, even in relatively high sperm numbers in the laboratory, is that the fertilizing spermatozoon is intrinsically different to the other spermatozoa, either due to its physiological status or the status of its attachment site. As the fate of supernumerary spermatozoa is determined before the plasma membrane of the oocyte depolarizes it goes to say that transmembrane voltage does not regulate sperm entry.

Figure 1. Scanning electron microscopy of the surface of a sea urchin oocyte (Paracentrotus lividus) deprived of its jelly layer. Note that all four spermatozoa are attached well to the surface. |The image, fixed 5 s after insemination, gives the impression that the spermatozoa have the same capacity to enter the oocyte and/or the attachment sites are equal in receptivity. There is no evidence for either assumption.
Low sperm–oocyte ratios and gamete morphology across the animal kingdom
In the majority of animals, sperm–oocyte ratios and/or gamete morphology preclude the necessity for ‘polyspermy preventing mechanisms at the oocyte surface’. In addition, many authors have indicated that there is no evidence, nor indeed any need, for a fast electrical block to polyspermy in mammals and teleosts (Dale and Monroy, Reference Dale and Monroy1981; Dale and DeFelice, Reference Dale and DeFelice2011; Dale, Reference Dale2014, Reference Dale2016, Reference Dale and Skinner2018a, Reference Dale2018b; Wozniak and Carlson, Reference Wozniak and Carlson2020).
Mammals may produce up to 109 spermatozoa for every oocyte produced, however precise behavioural traits are required for successful fertilization. The vast majority of spermatozoa, deposited in the female tract in synchrony with the ovulatory cycle, are eliminated rapidly, with only a handful reaching the ampullae or the ampulla–isthmic junction. To avoid oocyte ageing, sperm ascent and oocyte descent must be synchronized. The mucus-filled cervix in humans is the first barrier to sperm progression, while contractile activity of the uterine wall aids spermatozoa to the lower isthmus. Migration from the isthmus to the ampullae appears to be due to both sperm motility and contractile activity of the oviduct. The spermatozoa in the tube are in close contact with the epithelium until they are released, probably triggered by the descending oocyte. In conclusion, in mammals, the number of spermatozoa reaching the site of fertilization is regulated by the female tract and, in any case, is a handful (see Dale, Reference Dale2018b, for references).
In insects, squid, and some teleosts, oocytes lack cortical granules and the extracellular coat is a thick impenetrable outer membrane called the chorion. In these species, the spermatozoa do not have an acrosome and the spermatozoon enters the oocyte through a preformed entry site, the micropyle (Dale and Monroy, Reference Dale and Monroy1981; Dale, Reference Dale1983). In fish, a sperm attractant, a glycol protein, has been identified as responsible for guiding the spermatozoa to the micropyle (Yanagimachi et al., Reference Yanagimachi, Cherr and Matsubara2013). In the anuran Discoglossus pictus, oocytes are highly polarized with the animal pole marking the position of the meiotic plate and organelles distributed in a gradient towards the vegetal pole. Here, the spermatozoa may only enter through a restricted depression at the animal pole, called the dimple, where the fine structural organization is different to the rest of the oocyte surface (Campanella, Reference Campanella1975). In tunicates, the spermatozoon enters the oocyte at a preferential site at the vegetal pole. Polarized sperm entry coincides with polarized oocyte activation events. The first event is the release of calcium from intracellular stores that traverses the oocyte from the point of sperm entry to the antipode in a wave (Dale et al., Reference Dale, Wilding, Coppola and Tosti2010). For example, in jellyfish and the anuran Xenopus laevis, in which the sperm enters the animal pole of the oocyte, the calcium wave starts at the animal pole and traverses the oocyte to the antipode (Busa et al., Reference Busa, Ferguson, Joseph, Williamson and Nuccitelli1985; Yamashita, Reference Yamashita1998), while in ascidian and nemertean oocytes, the wave initiates at the vegetal pole, the site of sperm entry (Dale and Monroy, Reference Dale and Monroy1981; Stricker, Reference Stricker1999; Wilding et al., Reference Wilding, Marino, Monfrecola and Dale2000). In teleosts, in which the spermatozoa are forced to enter the animal pole through the micropyle, the calcium wave starts at the animal pole (Gilkey et al., Reference Gilkey, Jaffe, Ridgway and Reynolds1978; Yoshimoto et al., Reference Yoshimoto, Iwamatsu, Hirano and Hiramoto1986; Ridgeway et al., Reference Ridgeway, Gilkey and Jaffe1997). Since in mammals and echinoderms, the calcium wave is also initiated at the point of sperm entry (Miyazaki et al., Reference Miyazaki, Hashimoto, Yoshimoto, Kishimoto, Igusa and Hiramoto1986; Swann and Ozil, Reference Swann and Ozil1994; Deguchi et al., Reference Deguchi, Shirakawa, Oda, Mohri and Miyazaki2000); more work is needed to determine whether there are also preferential sperm entry sites in these groups.
In some animals it is normal for more than one spermatozoon to enter the oocyte cytoplasm; however, only one sperm nucleus interacts with the maternal nucleus to form the zygote nucleus, while the others degenerate. Physiologically polyspermic animals include ctenophores, many insects, elasmobranchs, some amphibians, reptiles, and birds (Monroy, Reference Monroy1963; Rothschild, Reference Rothschild1954; Dale and Monroy, Reference Dale and Monroy1981; Snook et al., Reference Snook, Hosken and Karr2011).
The most classical studies include those on the urodeles by Fankhauser (Reference Fankhauser1925, Reference Fankhauser1932, Reference Fankhauser1948), and those in birds by Harper (Reference Harper1904). In birds, hundreds of millions of sperms are inseminated, but only a few hundred reach the ovum (Bakst et al., Reference Bakst, Wishart and Brillard1994). The vast majority are ejected by the female tract early after copulation. Recently, it has been shown in the zebra finch and domestic fowl that the female tract regulates the number of sperm reaching the site of fertilization and it has been suggested that, although one or few spermatozoa are sufficient to activate the oocyte, the presence of several supernumerary spermatozoa in the cytoplasm of the oocyte is a prerequisite for embryogenesis (Hemmings and Birkhead, Reference Hemmings and Birkhead2015).
In some insects and nematode worms, sperm utilization is very efficient. In Drosophila, Lefevre and Jonsson (Reference Lefevre and Jonsson1962) discovered a 1:1 ratio between the progeny produced and the number of sperms in the seminal receptacles. In the nematode worm Caenorhabditis elegans, every spermatozoon fertilizes an oocyte; however, not all oocytes are fertilized because in fact oocytes are produced in excess (Ward and Carroll, Reference Ward and Carroll1979). This high efficiency of sperm utilization in insects and nematodes may be an important adaptation as it enables a minimal volume and nutrients for the stored sperm (Parker, Reference Parker1970). Low numbers reduce genetic variability, which may be offset by sperm displacement from second matings (Parker, Reference Parker1970; Ward and Carroll, Reference Ward and Carroll1979).
In ∼0.1% of all animal species, reproduction is achieved without the participation of spermatozoa, a phenomenon called parthenogenesis. In aphids, for example, parthenogenetic generations alternate with those generated by fertilization and in bees an egg may be parthenogenetic or fertilized. The creation of female parthenogenetic offspring is widespread among insects such as some Phasmida, Diptera and Lepidoptera, while in the crustacea the best known example of parthenogenesis is the brine shrimp Artemia salina. Nematodes, rotifers, snails and flatworms also include a few parthenogenetic species, whereas in the vertebrates the most common form of parthenogenesis and female-only species are found in the lizards (Dale, Reference Dale and Skinner2018a).
Bottlenecks reducing the number of spermatozoa reaching the oocyte
Successful fertilization in all animals requires the progressive and reciprocal activation of both gametes as they interact with each other. Those that are not activated in a correct temporal sequence fall by the wayside. Activation starts at spawning or ejaculation and involves several steps triggered by ionic signals in the environment, chemotactic triggers and, finally, receptors in the oocyte coats. Both the outer layers, the cumulus oophorus in mammals and the jelly layer in echinoderms and amphibians, and the inner coat, the zona pellucida in mammals and the vitelline coat in echinoderms, have receptors that allow the progression of the successful spermatozoon. The cumulus oophorus for example is a matrix of cells and hyaluronic acid, which is a polymer of alternating N-acetylglucosamine and glucuronic acid with several protein components. In the starfish, ARIS, Co-Aris and asterosap, high molecular weight glycoconjugates, trigger the acrosome reaction in the successful spermatozoa. In the sea urchin, a 30,000 mwt polypeptide called bindin, exposed during the acrosome reaction, is responsible for sperm binding to the vitelline coat; whereas a glycoprotein, ZP3, in the mammalian zona pellucida causes the acrosome reaction. Finally, IZUMO1 on the spermatozoan plasma membrane, and JUNO on the egg plasma membrane, mediate gamete adhesion in mammals and humans, while a putative fusion protein Tmem95 may favour fusion in the mammals (see Dale, Reference Dale2018b for a fuller explanation of sperm–oocyte interaction and full references). Van Blerkom and Caltrider (Reference Van Blerkom and Caltrider2013) and Van Blerkom and Zimmermann (Reference Van Blerkom and Zimmermann2016) have also shown that the distribution of ganglioside-enriched microdomains on the mammalian oocyte plasma membrane restricts the areas available for sperm–oocyte interaction.
There is, to date, no precise explanation why some spermatozoa progress through the extracellular coats and others do not. Whether the receptors in the coats are topographically limited and sparse and/or the spermatozoa are not all capable of responding to the signal is a matter for conjecture.
Where the controversy lies: the echinoderms and amphibians
The fertilization success of sea urchins in nature depends on the spawning behaviour of the animals, population size, current velocity, oocyte size, sperm swimming capacity, and many other factors. Sea urchins produce ∼10,000 spermatozoa for every oocyte produced, which is one-hundredth of the typical concentration used in the laboratory (106/ml); however, data collected from natural spawning show a low fertilization rate, with fertilization success in free-spawning benthic organisms often less than 1% (Levitan, Reference Levitan1993). Therefore, in the environment, sperm–oocyte collisions in sea urchins may be rare, and the availability of sperm may affect female reproductive success (see review on sperm limitation, Levitan and Petersen, Reference Levitan and Petersen1995). If indeed, under natural conditions, sperm–oocyte ratios are low then selective pressures may have favoured the achievement of monospermy, rather than the evolution of mechanisms to prevent polyspermy.
Without considering sperm–oocyte ratios or indeed fertilization behaviour in the natural habitat, Rothschild and Swann in the 1940s and 1950s studied sperm–oocyte interactions in sea urchins in the laboratory, deciding concentrations and conditions, and came up with the idea that the fertilizing spermatozoon induced a fast, yet partial, change in the oocyte surface that preceded the cortical reaction and that reduced sperm receptivity by 1/20th.
Observing sea urchin oocytes under dark field illumination, the authors noted a change in the scattering of light properties that covered the oocyte cortex in ∼20 s at 18°C:
‘At the site of spermatozoon entry, there is a localized and transient decrease in light scattering, the elevation of a fertilization cone and it is here that the fertilization membrane starts to elevate.’
Rothschild then made the assumption that a suspension of spermatozoa was analogous to an assembly of gas molecules, and calculated the number of sperm–oocyte collisions at 105/ml to be 1.6, at 106/ml to be 16, and at 107/ml to be 160, assuming the translator speed of sperm to be 190 microns/s.
Using immature oocytes [the germinal vesicle (GV) stage], Rothschild and Swann demonstrated that, from an expected 4500 collisions at a sperm density of 107/ml, in a 5-min period, <100 sperm actually entered the oocyte. The authors concluded that:
‘only a fraction of the spermatozoa which collide with the egg surface are able to initiate activation’
and
‘attachment of the spermatozoon to an egg is not followed by fertilization unless there exists a particular orientation on a molecular scale, between the egg and sperm surfaces, and provided there has been no previous interaction between spermatozoa and Gynogamone II’
(an agglutinating substance released from eggs). In later experiments, Rothschild and Swann, mixed oocytes and sperm at known densities for varying periods of time and, by treating the fertilization reaction as a first-order chemical reaction, found that the fraction of monospermic oocytes increased in time according to the relationship, M(t) = 1 − e− αt below sperm densities of 3 × 106/ml.
The relationship is similar for polyspermic oocytes at densities between 7 × 107/ml and 3 × 108/ml giving a rate of appearance of polyspermic oocytes as α1. They argued that as α1 (the refertilization rate) was found to be much less than α (the monospermic rate), a rapidly acting partial block reduced the probability of successful reactions after the first had occurred. To perform these calculations of refertilization rates (to fix a T = 0), the authors used a totally unnatural sperm concentration of 108/ml, which is slightly less than that found in the testis before dilution. Rothschild and Swann (Reference Rothschild and Swann1951, Reference Rothschild and Swann1952) point out the drawbacks of their own experiments: as α1 is the rate of increase of polyspermic eggs, it is perforce an underestimate of the successful collision rate because it does not take into account the actual number of sperm per polyspermic egg. In addition, the nonlinear dependence of α on n may be due to sperm–sperm interactions at higher densities, while we have not taken into account chemotaxis from egg-exuded components, the presence of the jelly or the declining fertilizing capacity of spermatozoa in time. Although an exercise to be studied by all biology students, the Rothschild hypothesis has fundamental flaws. Fertilization is not a first-order chemical reaction and spermatozoa are not analogous to gas molecules. Second, it does not consider the possibility that there is a limited number of sperm entry sites on the oocyte surface. Last, it assumes that all spermatozoa are equal in their capacity to penetrate the cell. This is incorrect. Only competent spermatozoa that encounter and respond to the correct sequence of triggering events as they progress through the oocyte investments are successful.
The hypothesis of the fast electrical block to polyspermy
Scientists, some 20 years after Rothschild – again ignoring both the fertilization dynamics in nature and the flaws in the Rothschild kinetic experiments – hypothesized the concept of a fast electrical block to polyspermy in sea urchin oocytes (Jaffe, Reference Jaffe1976). First, electrical measurements across the oocyte plasma membrane were recorded (Jaffe, Reference Jaffe1976). Subsequently, ion channels responsible for these currents were identified (Tosti and Ménézo, Reference Tosti and Ménézo2016). Finally, the authors insinuated that these channels evolved as polyspermy-blocking mechanisms (Wozniak and Carlson, Reference Wozniak and Carlson2020). There is no evidence that these ion channels in the oocyte plasma membrane can, in any way, regulate sperm entry. This would require the identification of a transmembrane mechanism that is sensitive to voltage and regulates sperm entry.
In a recent review, Wozniak and Carlson (Reference Wozniak and Carlson2020) describe electrical changes that occur at activation in oocytes and the ion channels underlying them, referring to some publications, but omitting many others (Dale, Reference Dale2014). Wozniak and Carlson (Reference Wozniak and Carlson2020) do not give evidence for a fast block to polyspermy. Instead they cite circumstantial experiments to try to support the hypothesis of a fast electrical block. They state:
‘In 1976, Laurinda Jaffe voltage clamped sea urchin eggs and demonstrated the depolarization of their membranes dictated whether sperm could enter.’
Both statements are incorrect. The oocytes in question were not voltage clamped and sperm entry was not directly observed. Polyspermy was inferred from abnormal cleavage patterns in this paper (Jaffe, Reference Jaffe1976) and in a later study in which external Na+ was reduced to interfere with the membrane depolarization (Jaffe, Reference Jaffe1980). Limatola et al. (Reference Limatola, Vasilev, Chun and Santella2019) showed that abnormal cleavage patterns in sea urchin oocytes fertilized in low Na+ seawater were due to alterations in the actin cytoskeleton and not to multiple sperm entry. These latter authors were careful not to adulterate the sea urchin oocytes before experimentation, that is, their jelly layers were intact. Perhaps ion substitution experiments in fertilization experiments from other phyla should be re-visited. In an attempt to mimic the depolarization of the oocyte plasma membrane at fertilization, Jaffe (Reference Jaffe1976) artificially depolarized the membrane of the unfertilized oocyte by injecting current through the intracellular electrode. To do this, jelly-free oocytes that adhered to plastic dishes were selected for recording. These impaired eggs were then inseminated. Jaffe (Reference Jaffe1976) noted that, although positive membrane voltages prevented all attached spermatozoa from activating the oocyte, when ‘the current was turned off, only one sperm had entered the egg’. Depolarization induced by current injection is due to cations being forced into the cytoplasm of the oocytes from the recording microelectrode that contains high molarity KCl. This is not what happens at fertilization. The fertilizing spermatozoon induces depolarization of the oocyte plasma membrane by opening ion channels and therefore increasing membrane conductance (Dale, Reference Dale2014). Correct voltage clamp experiments were, however, carried out by authors in later papers. Shen and Steinhardt (Reference Shen and Steinhardt1984) demonstrated that, while positive voltages blocked the entry of all the spermatozoa attached to the surface of the oocyte, a negative repolarization permitted the entry of only one spermatozoon. The authors concluded:
‘We were somewhat surprised to observe a low rate of polyspermy after the application of a window of negativity. Despite the presence of numerous sperm, all presumably blocked at the same point, nearly all the fertilizations were monospermic.’
Using a fluorescent dye to indicate cell–cell fusion, Hinkley et al. (Reference Hinkley, Wright and Lynn1986) showed that, in sea urchin oocytes voltage clamped at −20 mV, only one spermatozoon fused with the oocyte. In addition, Lynn et al. (Reference Lynn, McCulloh and Chambers1988), using clamp voltages of −20 to +17 mV, showed that normal monospermic entry was associated with a typical biphasic fertilization current and quoted:
‘As many as 5–25 sperm may attach before a successful encounter occurs that results in an electrophysiological response.’
There is only one explanation for these voltage clamp experiments: the fertilizing spermatozoon is intrinsically different to the supernumerary spermatozoa, either due to its physiological status or the status of its attachment site. The supernumerary spermatozoa are not excluded by changes in the membrane potential of the oocyte. They are already excluded beforehand (Figure 2).
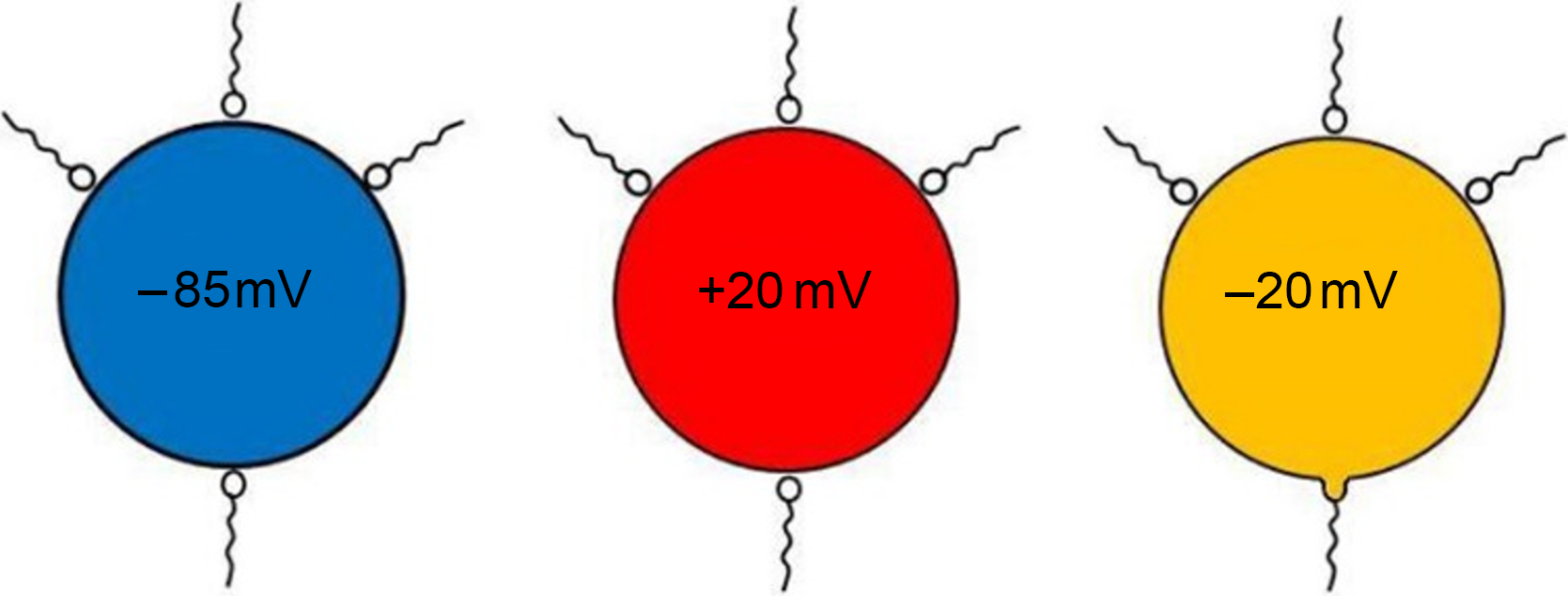
Figure 2. A schematic representation of inseminated voltage clamped sea urchin oocytes. Voltage clamping the oocyte at −85 mV or +20 mV (where large amounts of current need to be injected to hold the voltage at these values) stops all spermatozoa attached to the surface from fusing and entering the oocyte. Voltage clamping the oocyte at −20 mV, near to its natural resting potential (which requires small amounts of holding current), allows the entry of only one spermatozoon. This spermatozoon must be intrinsically different from the others and/or it is attached to a preferential entry site.
Additional evidence that the fertilizing spermatozoon is intrinsically different to its competitors is evident from the time course of the early electrical events. Sea urchin oocytes are large and often transparent, making it easy to follow fertilization events without resorting to fixation. As noted by Dale and de Santis (Reference Dale and de Santis1981), and later by Lynn et al. (Reference Lynn, McCulloh and Chambers1988), many spermatozoa attach to the oocyte, before one, the fertilizing spermatozoon, distinguishes itself by gyrating around its point of attachment. Approximately 3 s later, a small electrical step depolarization occurs with no change in the morphology of the oocyte surface or sperm behaviour until a further 9 s later when a larger bell-shaped depolarization starts. The successful spermatozoon then stiffens, stops gyrating, and the cortical reaction is initiated at this point (Figure 3). A protuberance of the oocyte cortex (the fertilization cone) is formed and the sperm head slowly disappears into this cone, while the cortical reaction spreads across the oocyte to the antipode. The cortical reaction leads to the elevation of the fertilization membrane and is completed during the repolarizing phase of the fertilization potential. Unsuccessful spermatozoa, that is, those that do not enter the oocyte, attach to the oocyte surface, but do not generate either electrical step depolarizations or fertilization cones, and continue gyrating around their point of attachment for many seconds until their energy resources are depleted and they fall limp to the oocyte surface (Dale, Reference Dale2016).

Figure 3. Schematic voltage recordings from a mature sea urchin oocyte (often called an egg) and an immature GV stage sea urchin oocyte showing the correlation between sperm behaviour and electrical events at the oocyte plasma membrane. In eggs, of the many spermatozoa that attach to the surface, one, the fertilizing spermatozoon, distinguishes itself by gyrating around its point of attachment. At 3 s later this pre-destined spermatozoon generates a small step-like depolarization across the egg plasma membrane which lasts for 9 s until a larger bell-shaped depolarization starts. The successful spermatozoon then stiffens, stops gyrating, and the cortical reaction is initiated at this point 3. Unsuccessful spermatozoa, that is, those that do not enter the oocyte, attach to the oocyte surface, but do not generate either electrical step depolarizations or fertilization cones, and continue gyrating around their point of attachment for many seconds until their energy resources are depleted and they fall limp to the oocyte surface. In GV stage oocytes, successful spermatozoa, which may number up to 20, gyrate around their point of attachment and cause a long step depolarization, which accumulates in a staircase fashion, whereas the step depolarization generated by spermatozoa that do not enter reverses after ∼50 s.
Wozniak and Carlson (2019) also acknowledge that the hypothetical ‘fast electrical block depolarization’ is preceded by a small step current, but fail to interpret its significance. As only one of all the attached spermatozoa is capable of generating the small step current and this occurs well before the larger later depolarization, we arrived at the same conclusion as that drawn from the voltage clamp experiments. The fate of supernumerary spermatozoa is determined well before the plasma membrane of the oocyte depolarizes. Transmembrane voltage does not serve to regulate sperm entry.
Conclusion
In this short review, I have given examples across the animal kingdom showing that sperm–oocyte ratios at the site of fertilization in nature are very modest, often approaching unity, and this is modulated in externally fertilizing species by dilution and by the female reproductive tract in those species with internal fertilization. This is then followed by a bottleneck created by the oocyte extracellular coats that further reduces the effective number of sperm reaching the oocyte surface. To progress to the oocyte plasma membrane, the fertilizing spermatozoon must encounter and respond to a correct sequence of signals from the oocytes extracellular coats. Those that do not, are halted in their progression by defective signalling and fall to the wayside. Final success and entry may be finely tuned by the spermatozoon anchoring to a predetermined site on the plasma membrane.



