


Turmeric (Curcuma longa L.) is a popular ancient herb that has been traditionally used in Asian countries for improving human health and beauty( Reference Gupta, Sung and Kim 1 ). The traditional medical system in China and India recommends the use of turmeric for several ailments related to the gut, liver, lungs and skin( Reference Gupta, Sung and Kim 1 ). Curcuma oil (C. oil), a lipophilic fraction from turmeric, exhibits several favourable effects on cell proliferation, inflammation, oxidation and platelet activation( Reference Singh, Jain and Misra 2 ). C. oil chiefly comprises ar-d-turmerone and α/β-turmerone. The other minor constituents are curcumene, zingiberene, germacrone, curcumerone, zedoarone, sedoarondiol, isozdedoaronidiol, curcumenone and curlone( Reference Jain, Prasad and Pal 3 ). Recent work from our laboratory has demonstrated an anti-hyperlipidaemic effect of C. oil in hamsters( Reference Singh, Jain and Misra 2 ). Hyperlipidaemia often leads to atherosclerosis, which is characterised by endothelial dysfunction, arterial macrophage accumulation, inflammation and vascular smooth cell proliferation( Reference Moore and Tabas 4 ). Advanced atherosclerotic plaques are rich in lipids, inflammatory cells, and matrix components such as proteoglycans, elastin and collagen( Reference Moore and Tabas 4 , Reference Kolodgie, Virmani and Burke 5 ). Arterial oxidised LDL (OxLDL) is internalised by vessel macrophages, leading to foam-cell formation and inflammation( Reference Tiwari, Singh and Barthwal 6 ). Peritoneal macrophages derived from hyperlipidaemic animals have been shown to enhance cellular lipids and mRNA transcripts of genes related to lipid homeostasis( Reference Saeed, Otsuka and Polavarapu 7 ). Anti-hyperlipidaemic drugs such as statins and fibrates reduce the progression of atherosclerosis by suppressing macrophage foam-cell formation and inflammation( Reference Li and Glass 8 ). In experimental animal models, the development of atherosclerosis is accelerated by combining a high-cholesterol (HC) diet with endothelial injury( Reference Singh, Tiwari and Dikshit 9 ). Oxidative stress in the vascular bed significantly contributes to the initiation and progression of atherosclerosis( Reference Heistad 10 ). The application of FeCl3 to the arterial adventitia induces oxidative stress, free radical generation, lipid peroxidation and endothelial dysfunction( Reference Eckly, Hechler and Freund 11 ). FeCl3-induced arterial oxidative injury (Ox-injury) is often used to induce neointimal proliferation and atherosclerosis in experimental animal models( Reference Tian, Hu and Sun 12 ). Recent reports have suggested that shear stress induced by partial carotid ligation (PCL) in hyperlipidaemic mice accelerated arterial lipid accumulation, endothelial dysfunction, inflammation and macrophage foam-cell formation( Reference Nam, Ni and Rezvan 13 ).
Hamsters do not develop extensive atherosclerotic lesions even after the treatment with a high-fat diet for 6–9 months( Reference Singh, Tiwari and Dikshit 9 ). Therefore, in the present study, we examine the effect of C. oil on the development of advanced atherosclerotic lesions and macrophage foam-cell formation in PCL- and FeCl3-induced hamster models. We demonstrate that C. oil confers a protective effect on advanced atherosclerotic lesions, extracellular matrix components, inflammation and macrophage foam-cell formation.
Experimental methods
Materials
Lipid estimation kits were obtained from Randox Laboratories. The Amplex Red Cholesterol Assay Kit was obtained from Invitrogen (Molecular Probes). The RevertAid™ H Minus First Strand cDNA Synthesis Kit and Maxima SYBR Green were obtained from Thermo Fisher Scientific, Fermentas, Inc. Cell-culture reagents and histological staining (Movat's pentachrome and Sirius Red) were procured from Sigma-Aldrich. Gene-specific primers were obtained from Integrated DNA Technology. All other fine chemicals used in the study were procured from Sigma.
Ethics statement
Animal experimental protocols were approved by the Institutional Animal Ethics Committee, and all procedures were carried out in strict accordance with the guidelines of the Committee for the Purpose of Control and Supervision of Experiments on Animals, which conforms to the international norms of the Indian National Science Academy. Ethical approval was granted by the institutional ethics committee (human research) of the Council of Scientific and Industrial Research-Central Drug Research Institute (CSIR-CDRI) for the collection of blood plasma samples from healthy subjects.
Experimental model of atherosclerosis and treatments
Male golden Syrian hamsters (110–115 g) maintained at the central animal house facility of CSIR-CDRI, Lucknow were used in the present study. After 1 week of HC diet feeding (chow diet supplemented with 1 % cholesterol and 10 % saturated fat from coconut oil)( Reference Singh, Jain and Prakash 14 ), hamsters were subjected to FeCl3-induced Ox-injury or PCL. For surgical procedures, hamsters were anaesthetised with an intraperitoneal injection of ketamine (80 mg/kg) and xylazine (10 mg/kg). To perform carotid ligation, the left carotid artery was carefully exposed under a dissecting microscope, and the external carotid artery, the internal carotid artery and the occipital artery except the superior thyroid artery were ligated using 6-0 silk sutures( Reference Nam, Ni and Rezvan 13 ).
To carry out Ox-injury, a parafilm strip was placed beneath the left carotid artery to prevent any leakage of FeCl3 into the surrounding tissues. Oxidative vascular injury was induced by placing a FeCl3 (10 %, w/v)-saturated Whatman no. 1 filter paper strip on the adventitia of the carotid artery for 60 s( Reference Tian, Hu and Sun 12 ). After removal of the filter paper, the site of the incision was washed three times with sterile normal saline.
After surgical incisions were sutured, hamsters were maintained on antibiotics (ampicillin, 30 mg/kg per d, intraperitoneally) and analgesics (pentazocine lactate, 10 mg/kg per d, intraperitoneally) for up to 3 d. At defined time intervals (at 1, 3 and 5 weeks following carotid injury), hamsters were euthanised under deep anaesthesia, and blood and tissue samples were collected for analysis.
C. oil was prepared and characterised as described previously( Reference Jain, Prasad and Pal 3 , Reference Ray, Singh and Khanna 15 ). Rhizomes of C. longa L. were procured from the local market at Lucknow, India. C. oil was extracted with hexane from the dried rhizomes of C. longa L. syn. Curcuma domestica Valeton (family Zingiberaceae) at the CSIR-CDRI( Reference Jain, Prasad and Pal 3 , Reference Ray, Singh and Khanna 15 ).
To evaluate the anti-atherogenic effect of C. oil, hamsters were randomly divided into four groups fed with the following diets: HC diet; HC diet with vehicle (0·25 % carboxymethylcellulose); HC diet with ezetimibe (1 mg/kg per d); HC diet with C. oil (100 or 300 mg/kg per d). Each group had a minimum of six animals. After 1 week of treatment, hamsters were subjected to PCL or Ox-injury followed by treatment with C. oil or ezetimibe for an additional 4 weeks. A carboxymethylcellulose suspension of C. oil and ezetimibe was administered orally (0·5 ml/animal per d).
Peritoneal macrophage isolation
Macrophages were harvested from the peritoneal cavity of the hamsters fed either a chow or a HC diet with or without C. oil (300 mg/kg per d, orally) after treatment for 6 weeks. At 3 d before peritoneal macrophage isolation, hamsters were treated with sterile Brewer's thioglycollate medium (3 %, w/v, intraperitoneally, 100 μl per animal). On the day of the experiment, peritoneal fluid was collected in ice-cold sterile PBS and then centrifuged at 1200 rpm for 6 min to pellet the cells. Peritoneal cells were washed twice with PBS and suspended in Roswell Park Memorial Institute (RPMI)-1640 medium supplemented with 10 % fetal bovine serum, streptomycin (100 μg/ml) and penicillin (100 IU/ml). Subsequently, the cells were plated in six-well plates (2 × 106 cells/well) and incubated for 3 h at 37°C in a CO2 incubator to allow macrophage adherence( Reference Senokuchi, Matsumura and Sakai 16 ). After incubation, all non-adherent cells were removed, the wells were washed with PBS, and the adherent macrophages were processed for RNA or cholesterol extraction.
LDL isolation and oxidised LDL preparation
LDL was isolated from human plasma by sequential ultracentrifugation using an LE-80 ultracentrifuge (Beckman Coulter). LDL protein concentration was measured using the Bicinchoninic Acid Protein Assay Kit (Pierce Biotechnology). To prepare OxLDL, native LDL (200 μg/ml protein) was diluted in PBS and oxidised by exposure to 5 μm-CuSO4 in PBS at 37°C for 24 h. Oxidation was terminated by the addition of Na2-EDTA (0·2 mm) and butylated hydroxytoluene (50 μm). Oxidation of LDL was determined by measuring relative electrophoretic mobility and thiobarbituric acid-reactive substances( Reference Han, Hajjar and Tauras 17 ).
Cell culture and treatment
Human monocytic THP-1 cells (European Collection of Cell Cultures) were suspended in RPMI-1640 medium containing 10 % fetal bovine serum and 0·1 ml of pencillin–streptomycin/ml (catalogue no. P0781; Sigma), and then plated into six-well plates (2 × 106 cells/well). Monocytic cells were then differentiated into macrophages by the addition of phorbol 12-myristate 13-acetate (200 nm) for 72 h( Reference Tiwari, Singh and Singh 18 ). To evaluate the effect of C. oil on macrophage cholesterol accumulation and inflammation, THP-1 macrophages were pretreated for 18 h with C. oil (1, 3 and 10 μg/ml) followed by OxLDL (40 μg/ml) treatment for 48 h. Cell supernatant was collected for cytokine measurements, and the adhered cells were processed for cholesterol extraction or complementary DNA preparation( Reference Tiwari, Singh and Singh 18 ).
Plasma lipid analysis
Blood samples from overnight fasted animals were collected by cardiac puncture from anaesthetised animals and placed in a tube containing 3·8 % tri-sodium citrate. Whole blood was centrifuged at 4000 g for 10 min at 4°C to obtain plasma. Total cholesterol (TC), TAG, LDL-cholesterol (LDL-C) and HDL-cholesterol (HDL-C) concentrations were estimated by the automated analyser using commercial kits( Reference Singh, Jain and Misra 2 ).
Cytokine estimation using ELISA
Concentrations of TNF-α, IL-1β, transforming growth factor-β (TGF-β) and IL-10 were measured in the supernatant obtained from the control and OxLDL-stimulated THP-1 macrophages with or without C. oil pretreatment (1, 3 and 10 μg/ml). Cytokine levels were measured by conventional ELISA (BD OptEIA set; BD Biosciences) as described earlier( Reference Tiwari, Singh and Singh 18 ). In brief, samples and standards were added to specific capture-antibody pre-coated wells and incubated for 2 h at room temperature. Subsequently, the wells were washed with buffer and incubated with detection antibody for 1 h. Following washing and incubation with enzyme reagent, 3,3′,5,5′-tetramethylbenzidine substrate reagent was added and absorbance was read at 450 and 570 nm on an ELISA plate reader (Biotek Instrument, Inc.). The resulting standard curve was used for the calculation of absolute cytokine levels.
Aortic and cellular cholesterol estimation
Cholesterol in the carotid artery was quantified as described previously( Reference Singh, Jain and Misra 2 ). In brief, the carotid artery was removed, cleaned and weighed, and lipids were extracted with hexane–isopropanol (3:2). The extracted lipids were dried and resuspended in reagent-grade ethanol containing Nonidet P-40 (9:1).
Macrophages (THP-1 and peritoneal) were fixed with 2 % paraformaldehyde for 15 min and subsequently washed with PBS. Cholesterol was extracted by incubating the cells with 300 μl of reagent-grade ethanol for 30 min at 4°C( Reference McLaren, Calder and McSharry 19 ). After incubation, lipid-rich ethanol was collected for determination of cholesterol concentration. The cholesterol-extracted macrophages were incubated with SDS (0·1 %, w/v) and NaOH (0·2 m) for 30 min at room temperature, and total cellular protein contents were measured using the Pierce Bicinchoninic Acid Assay (Thermo Scientific).
TC and free cholesterol (FC) in the samples were measured using the Amplex Red Cholesterol Assay Kit (Invitrogen (Molecular Probes)). In brief, 50 μl of the diluted samples and standards were incubated with 50 μl of working reagent containing cholesterol oxidase, cholesterol esterase, horseradish peroxidase and Amplex Red reagent overnight for 30 min. After incubation, the plate was read on a fluorescence plate reader (BMG LABTECH GmbH) at an excitation wavelength of 540 nm and an emission wavelength of 590 nm. Cholesteryl ester was calculated after subtracting FC from TC. The cholesterol standard curve was generated using the cholesterol reference standard (0–8 μg/ml).
Histology
After 1, 3 and 5 weeks following PCL or Ox-injury, carotid arteries were isolated, cleaned and preserved in 10 % neutral buffered formalin solution for 24 h. Fixed arterial segments were then embedded in paraffin and consecutively cut into 5 μm-thick sections and subjected to Movat's pentachrome and Sirius Red staining. Alterations in the lesion of ground substances (such as mucin, glycosaminoglycans, proteoglycans and collagen) and plaque composition were determined using Movat's pentachrome staining( Reference Khanna, Jain and Singh 20 , Reference Finn, Nakano and Narula 21 ). Intimal thickness (μm), medial thickness (μm), lumen area (mm2), plaque area (PA, mm2) and internal elastic lamina (IEL) area (mm2) were measured in at least six arterial sections from each hamster using the computer-assisted image analysis software (Leica Qwin version 3.5.1). The intra-observer variation was less than 5 %. The PA was calculated by subtracting the lumen area from the IEL area. Cross-sectional narrowing (CSN) was defined as the extent to which the PA occupied the potential lumen area, and was calculated as follows:
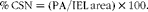 $$\begin{eqnarray} \%\,CSN = (PA/IEL\,area)\times 100. \end{eqnarray}$$
$$\begin{eqnarray} \%\,CSN = (PA/IEL\,area)\times 100. \end{eqnarray}$$
Type I and type III collagens in carotid arteries were quantified using Picrosirius Red staining( Reference Aikawa, Rabkin and Okada 22 ). In brief, deparaffinised and hydrated sections were stained with Picrosirius Red solution (Sirius Red (0·1 %) in the saturated aqueous solution of picric acid) for 1 h. After subsequent washing with acidified (0·5 % glacial acetic acid in distilled water) and tap water, sections were counter-stained with Mayer's haematoxylin solution. Dibutyl phthalate xylene-mounted sections were visualised under a polarised light microscope, and the image was acquired using the Leica Qwin image acquisition software (Leica Microsystems)( Reference Aikawa, Rabkin and Okada 22 ). Each section was photographed with the same exposure time. Type I collagen fibres exhibited yellow to orange colour, and type III collagen fibres exhibited green colour under polarised light.
Real-time RT-PCR
Quantitative gene expression analysis was performed using the LightCycler 480II Real-time PCR System (Roche Applied Science) along with Maxima SYBR Green reagents. Total RNA was extracted from freshly isolated carotid arteries of different groups using the Trizol isolation procedure as described previously(
Reference Singh, Jain and Misra
2
), and cDNA was synthesised using the cDNA Synthesis Kit (Thermo Scientific) according to the manufacturer's protocol. The mRNA expression of lipid-related and inflammatory genes involved in atherosclerosis and macrophage foam-cell formation was quantified using specific primers (Table 1). Amplification conditions used in the present study consisted of an initial pre-incubation step at 94 or 95°C for 10 min followed by amplification of the target DNA for forty-five cycles (95°C for 10 s and 54–60°C (as applicable) for 10 s). Melting curve analysis was carried out immediately after amplification using the manufacturer's protocol. The relative fold difference between an experimental and calibrator sample was calculated using the Q-Gene software (comparative C
p (
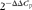 $$2^{ - \Delta \Delta C _{p}} $$
) method; BioTechniques Software Library: www.BioTechniques.com). β-Actin was used as the internal control to calculate the relative expression level(
Reference Tiwari, Singh and Singh
18
).
$$2^{ - \Delta \Delta C _{p}} $$
) method; BioTechniques Software Library: www.BioTechniques.com). β-Actin was used as the internal control to calculate the relative expression level(
Reference Tiwari, Singh and Singh
18
).
Table 1 List of the primers used in the study

CD68, cluster of differentiation 68; MMP-2 and MMP-9, matrix metalloproteinase-2 and matrix metalloproteinase-9; CD45, cluster of differentiation 45; IFN-γ, interferon-γ; TGF-β, transforming growth factor-β; LXRα, liver X receptor-α; ABCA1, ATP-binding cassette A1; ABCG1, ATP-binding cassette G1; CD36, cluster of differentiation 36.
Statistical analysis
Data are presented as means with their standard errors. A two-tailed, unpaired Student's t test was used to calculate the significant difference between two groups. Comparison of more than two groups was made using a one-way ANOVA followed by Dunnett's post hoc test. P≤ 0·05 was considered statistically significant. All statistical analyses were performed with the GraphPad Prism 5.0 program (GraphPad, Inc.).
Results
Curcuma oil reduces aortic macrophage and lipid accumulation in the partial carotid ligation and oxidative-injury hamster model
Plasma lipid concentrations were significantly higher in hamsters analysed at 1, 3 and 5 weeks following carotid injury than in their matched chow-fed control group (Figs. 1 and 2). As the chow-fed group analysed at different days had similar lipid levels, the data from hamsters of the different groups were pooled (Figs. 1 and 2). Plasma lipid levels were equivalent in the HC-fed group with or without arterial injury (data not shown). As expected, plasma levels of TC, TAG, LDL-C and HDL-C increased progressively at 1 and 3 weeks after PCL or Ox-injury in the HC-fed group (Figs. 1(a) and 2(a)). However, no further increase was observed after 3 weeks, and lipid levels at 5 weeks were almost similar to those at 3 weeks (Figs. 1(a) and 2(a)).

Fig. 1 Curcuma oil (C. oil) reduces aortic lipid and macrophage accumulation following partial carotid ligation (PCL) in hyperlipidaemic hamsters. (a) Plasma lipids (n 8), (b) total cholesterol (TC, n 6), (c) free cholesterol (FC, n 6), (d) cholesteryl ester (CE, n 6) and (e) cluster of differentiation 68 (CD68) mRNA expression (n 5) in the aorta at 1, 3 and 5 weeks following PCL. Values are means, with their standard errors represented by vertical bars. Mean value was significantly different from that of the chow diet-fed group: * P< 0·05, ** P< 0·01, *** P< 0·001 (one-way ANOVA). Mean value was significantly different from that of the uninjured control group (right common carotid artery of the PCL experimental animal): † P< 0·05, †† P< 0·01, ††† P< 0·001 (two-tailed, unpaired Student's t test). Mean value was significantly different from that of the high-cholesterol (HC)+PCL group: ‡‡ P< 0·01, ‡‡‡ P< 0·001. Mean value was significantly different from that of the HC+PCL-5 weeks group: § P< 0·05, §§ P< 0·01, §§§ P< 0·001 (one-way ANOVA). (a) ![]() , Chow-fed;
, Chow-fed; ![]() , HC+PCL-1 week;
, HC+PCL-1 week; ![]() , HC+PCL-3 weeks;
, HC+PCL-3 weeks; ![]() , HC+PCL-5 weeks;
, HC+PCL-5 weeks; ![]() , HC+PCL-5 weeks+C. oil-100 mg/kg;
, HC+PCL-5 weeks+C. oil-100 mg/kg; ![]() , HC+PCL-5 weeks+C. oil-300 mg/kg;
, HC+PCL-5 weeks+C. oil-300 mg/kg; ![]() , HC+PCL-5 weeks+ezetimibe-1 mg/kg. (b–d)
, HC+PCL-5 weeks+ezetimibe-1 mg/kg. (b–d) ![]() , Uninjured control;
, Uninjured control; ![]() , HC+PCL;
, HC+PCL; ![]() , HC+PCL+C. oil-300 mg/kg;
, HC+PCL+C. oil-300 mg/kg; ![]() , HC+PCL+ezetimibe-1 mg/kg. (e)
, HC+PCL+ezetimibe-1 mg/kg. (e) ![]() , Uninjured control;
, Uninjured control; ![]() , HC+PCL-1 week;
, HC+PCL-1 week; ![]() , HC+PCL-3 weeks;
, HC+PCL-3 weeks; ![]() , HC+PCL-5 weeks;
, HC+PCL-5 weeks; ![]() , HC+PCL-5 weeks+C. oil-300 mg/kg;
, HC+PCL-5 weeks+C. oil-300 mg/kg; ![]() , HC+PCL-5 weeks+ezetimibe-1 mg/kg. LDL-C, LDL-cholesterol; HDL-C, HDL-cholesterol. To convert TC, LDL-cholesterol (LDL-C) and HDL-cholesterol (HDL-C) from mg/dl to mmol/l, multiply by 0·02586. To convert TAG from mg/dl to mmol/l, multiply by 0·01129.
, HC+PCL-5 weeks+ezetimibe-1 mg/kg. LDL-C, LDL-cholesterol; HDL-C, HDL-cholesterol. To convert TC, LDL-cholesterol (LDL-C) and HDL-cholesterol (HDL-C) from mg/dl to mmol/l, multiply by 0·02586. To convert TAG from mg/dl to mmol/l, multiply by 0·01129.

Fig. 2 Curcuma oil (C. oil) reduces aortic lipid and macrophage accumulation following ferric chloride-induced arterial oxidative injury (Ox-injury) in hyperlipidaemic hamsters. (a) Plasma lipids (n 8), (b) total cholesterol (TC, n 6), (c) free cholesterol (FC, n 6), (d) cholesteryl ester (CE, n 6) and (e) cluster of differentiation 68 (CD68) mRNA expression (n 5) in the aorta at 1, 3 and 5 weeks after Ox-injury. Values are means, with their standard errors represented by vertical bars. Mean value was significantly different from that of the chow diet-fed group: * P< 0·05, ** P< 0·01, *** P< 0·001 (one-way ANOVA). Mean value was significantly different from that of the uninjured control group (right common carotid artery of the Ox-injury experimental animal): † P< 0·05, †† P< 0·01, ††† P< 0·001 (two-tailed, unpaired Student's t test). ‡‡‡ Mean value was significantly different from that of the high-cholesterol (HC)+Ox-injury group (P< 0·001; one-way ANOVA). Mean value was significantly different from that of the HC+Ox-injury-5 weeks group: § P< 0·05, §§ P< 0·01, §§§ P< 0·001 (one-way ANOVA). (a) ![]() , Chow-fed;
, Chow-fed; ![]() , HC+Ox-injury-1 week;
, HC+Ox-injury-1 week; ![]() , HC+Ox-injury-3 weeks;
, HC+Ox-injury-3 weeks; ![]() , HC+Ox-injury-5 weeks;
, HC+Ox-injury-5 weeks; ![]() , HC+Ox-injury-5 weeks+C. oil-100 mg/kg;
, HC+Ox-injury-5 weeks+C. oil-100 mg/kg; ![]() , HC+Ox-injury-5 weeks+C. oil-300 mg/kg;
, HC+Ox-injury-5 weeks+C. oil-300 mg/kg; ![]() , HC+Ox-injury-5 weeks+ezetimibe-1 mg/kg. (b–d)
, HC+Ox-injury-5 weeks+ezetimibe-1 mg/kg. (b–d) ![]() , Uninjured control;
, Uninjured control; ![]() , HC+Ox-injury;
, HC+Ox-injury; ![]() , HC+Ox-injury+C. oil-300 mg/kg;
, HC+Ox-injury+C. oil-300 mg/kg; ![]() , HC+Ox-injury+ezetimibe-1 mg/kg. (e)
, HC+Ox-injury+ezetimibe-1 mg/kg. (e) ![]() , Uninjured control;
, Uninjured control; ![]() , HC+Ox-injury-1 week;
, HC+Ox-injury-1 week; ![]() , HC+Ox-injury-3 weeks;
, HC+Ox-injury-3 weeks; ![]() , HC+Ox-injury-5 weeks;
, HC+Ox-injury-5 weeks; ![]() , HC+Ox-injury-5 weeks+C. oil-300 mg/kg;
, HC+Ox-injury-5 weeks+C. oil-300 mg/kg; ![]() , HC+Ox-injury-5 weeks+ezetimibe-1 mg/kg. LDL-C, LDL-cholesterol; HDL-C, HDL-cholesterol. To convert TC, LDL-cholesterol (LDL-C) and HDL-cholesterol (HDL-C) from mg/dl to mmol/l, multiply by 0·02586. To convert TAG from mg/dl to mmol/l, multiply by 0·01129.
, HC+Ox-injury-5 weeks+ezetimibe-1 mg/kg. LDL-C, LDL-cholesterol; HDL-C, HDL-cholesterol. To convert TC, LDL-cholesterol (LDL-C) and HDL-cholesterol (HDL-C) from mg/dl to mmol/l, multiply by 0·02586. To convert TAG from mg/dl to mmol/l, multiply by 0·01129.
Treatment with C. oil (300 mg/kg) significantly reduced the plasma levels of TC, TAG and LDL-C and increased plasma HDL-C concentration in the HC-fed group exposed to PCL or Ox-injury when compared with those exposed to PCL or Ox-injury alone (Figs. 1(a) and 2(a)). Treatment with ezetimibe (1 mg/kg) reduced the plasma levels of TC, TAG and LDL-C (Figs. 1(a) and 2(a)).
The lower dose of C. oil (100 mg/kg) reduced only the plasma levels of TC and LDL-C. No effect was observed on the plasma levels of TAG and HDL-C in the groups exposed to both PCL (Fig. 1(a)) and Ox-injury (Fig. 2(a)). No significant difference in plasma lipid levels was observed between the groups exposed to PCL (Fig. 1(a)) or Ox-injury (Fig. 2(a)).
Carotid arteries of the PCL- or Ox-injury-induced groups showed a time-dependent increase in the levels of TC, FC and CE when compared with the uninjured control group (Figs. 1 (b)-(d) and 2(b)-(d)). Interestingly, aortic macrophage foam-cell accumulation also progressively increased as the mRNA expression level of cluster of differentiation 68 (CD68) elevated 1, 3 and 5 weeks following PCL or Ox-injury when compared with the uninjured control group (Figs. 1(e) and 2(e)). An increase in this pan-macrophage marker indicates an increase in the number of such cells in the lesion, and demonstrates their important role in disease progression.
The C. oil (300 mg/kg) treatment prevented lipid accumulation in the carotid artery of groups exposed to PCL or Ox-injury as evident by the significantly reduced levels of TC, FC and CE (Figs. 1 (b)-(d) and 2(b)-(d)), compared with those exposed to PCL or Ox-injury alone. The ezetimibe (1 mg/kg) treatment significantly decreased the aortic levels of TC, FC and CE (Figs. 1 (b)-(d) and 2(b)-(d)).
Accordingly, mRNA expression of CD68 was also significantly decreased after treatment with C. oil or ezetimibe in the PCL- or Ox-injury-induced group (Figs. 1(e) and 2(e)).
The lower dose of C. oil (100 mg/kg) did not inhibit aortic lipid accumulation and mRNA expression of CD68 in both PCL- and Ox-injury-induced groups (data not shown).
Curcuma oil prevents partial carotid ligation- and oxidative-injury-induced arterial narrowing and matrix metalloproteinase-2 and -9 expression
Differential staining with Movat's pentachrome revealed that atherosclerotic plaque resulting from PCL or Ox-injury composed of ground substances and mucin (blue stain), muscles (red stain) and elastin (black stain) (Figs. 3(a) and 4(a)). The collagen-stained areas (yellow) were hardly apparent in the aortic sections of the PCL-induced group (Fig. 3(a)), while intense staining was observed in the aortic sections of the Ox-injury-induced group (Fig. 4(a)). Further exploration of collagen content in the aortic sections with Picrosirius Red staining revealed that there was no significant change in the collagen content of the groups exposed to PCL when compared with the uninjured control group (Fig. 3(b)). Moreover, arterial mRNA expression of type-1 and type-3 collagen was also not significantly altered after exposure to PCL (Fig. 3(c)). However, in the Ox-injury group, Picrosirius Red staining showed that collagen content, specifically type I (yellow to orange colour under polarised light), was increased in the intimal area of the artery after exposure to Ox-injury (Fig. 4(b)). The mRNA expression of type-1 and type-3 collagen in the carotid artery was elevated at 1, 3 and 5 weeks in the groups exposed to Ox-injury when compared with the uninjured control group (Fig. 4(c)). A qualitative assessment of atherosclerotic plaque in the aorta of the PCL group showed intimal thickening, which is rich in muscle content (Fig. 3(a)). The Ox-injury group exhibited fibro-fatty lesions, which are rich in lipids, ground substances and collagen (Fig. 4(a)). Additionally, the mRNA expression of matrix metalloproteinase-2 (MMP-2) and matrix metalloproteinase-9 (MMP-9) was increased at 5 weeks after exposure to PCL or Ox-injury (Figs. 3(c) and 4(c)). Morphometric analysis showed a progressive increase in the intima:media thickness ratio and the percentage of CSN at 1, 3 and 5 weeks after exposure to PCL or Ox-injury when compared with the uninjured control group (Figs. 3 and (e) and 4). Since intimal thickening was not observed in the arterial sections of the uninjured control group, the intima:media thickness ratio and the percentage of CSN were not assessed in these groups (Figs. 3 and (e) and 4).

Fig. 3 Effect of curcuma oil (C. oil) on histomorphometric and biochemical changes induced by partial carotid ligation (PCL). Atherosclerotic lesion components and mRNA expression of various genes were analysed in the carotid artery at 1, 3 and 5 weeks after PCL and 5 weeks after PCL plus C. oil treatment. (a) Representative images of Movat's pentachrome-stained section of all groups (scale bar 50 μm, n 5). (b) Representative images of Picrosirius Red-stained sections of all groups under polarised light (scale bar 50 μm, n 5). (c) Aortic mRNA expression of collagen and matrix metalloproteinase (MMP, n 5), (d) intima:media thickness ratio (n 5) and (e) percentage of cross-sectional narrowing (CSN, n 5). Values are means, with their standard errors represented by vertical bars. Mean value was significantly different from that of the uninjured control group: * P< 0·05, ** P< 0·01 (one-way ANOVA). Mean value was significantly different from that of the high-cholesterol (HC)+PCL-1 week group: ††† P< 0·001 (one-way ANOVA). Mean value was significantly different from that of the HC+PCL-3 weeks group: ‡‡‡ P< 0·001 (two-tailed, unpaired Student's t test). Mean value was significantly different from that of the HC+PCL-5 weeks group: § P< 0·05, §§ P< 0·01, §§§ P< 0·001 (two-tailed, unpaired Student's t test). (c) ![]() , Uninjured control;
, Uninjured control; ![]() , HC+PCL-1 week;
, HC+PCL-1 week; ![]() , HC+PCL-3 weeks;
, HC+PCL-3 weeks; ![]() , HC+PCL-5 weeks;
, HC+PCL-5 weeks; ![]() , HC+PCL-5 weeks+C. oil-300 mg/kg. (d, e)
, HC+PCL-5 weeks+C. oil-300 mg/kg. (d, e) ![]() , HC+PCL-1 week;
, HC+PCL-1 week; ![]() , HC+PCL-3 weeks;
, HC+PCL-3 weeks; ![]() , HC+PCL-5 weeks;
, HC+PCL-5 weeks; ![]() , HC+PCL-5 weeks+C. oil-300 mg/kg. A colour version of this figure can be found online at http://www.journals.cambridge.org/bjn.
, HC+PCL-5 weeks+C. oil-300 mg/kg. A colour version of this figure can be found online at http://www.journals.cambridge.org/bjn.

Fig. 4 Effect of curcuma oil (C. oil) on histomorphometric and biochemical changes induced by arterial oxidative injury (Ox-inj). Atherosclerotic lesion components and mRNA expression of various genes were analysed in the carotid artery at 1, 3 and 5 weeks after Ox-inj and 5 weeks after Ox-inj plus C. oil treatment. (a) Representative images of Movat's pentachrome-stained section of all groups (scale bar 50 μm, n 5). (b) Representative images of Picrosirius Red-stained sections of all groups under polarised light (scale bar 50 μm, n 5). (c) Aortic mRNA expression of collagen and matrix metalloproteinase (MMP, n 5), (d) intima:media thickness ratio (n 5) and (e) percentage of cross-sectional narrowing (CSN, n 5). Values are means, with their standard errors represented by vertical bars. Mean value was significantly different from that of the uninjured control group: * P< 0·05, ** P< 0·01 (one-way ANOVA). Mean value was significantly different from that of the high cholesterol (HC)+Ox-inj-1 week group: ††† P< 0·001 (one-way ANOVA). Mean value was significantly different from that of the HC+Ox-inj-3 weeks group: ‡ P< 0·05, ‡‡‡ P< 0·001 (two-tailed, unpaired Student's t test). Mean value was significantly different from that of the HC+Ox-inj-5 weeks group: § P< 0·05, §§ P< 0·01 (two-tailed, unpaired Student's t test). (c) ![]() , Uninjured control;
, Uninjured control; ![]() , HC+Ox-inj-1 week;
, HC+Ox-inj-1 week; ![]() , HC+Ox-inj-3 weeks;
, HC+Ox-inj-3 weeks; ![]() , HC+Ox-inj-5 weeks;
, HC+Ox-inj-5 weeks; ![]() , HC+Ox-inj-5 weeks+C. oil-300 mg/kg. (d, e)
, HC+Ox-inj-5 weeks+C. oil-300 mg/kg. (d, e) ![]() , HC+Ox-inj-1 week;
, HC+Ox-inj-1 week; ![]() , HC+Ox-inj-3 weeks;
, HC+Ox-inj-3 weeks; ![]() , HC+Ox-inj-5 weeks;
, HC+Ox-inj-5 weeks; ![]() , HC+Ox-inj-5 weeks+C. oil-300 mg/kg. A colour version of this figure can be found online at http://www.journals.cambridge.org/bjn.
, HC+Ox-inj-5 weeks+C. oil-300 mg/kg. A colour version of this figure can be found online at http://www.journals.cambridge.org/bjn.
Treatment with C. oil (300 mg/kg) prevented the PCL- or Ox-injury-induced increase in the expression of MMP-2 and MMP-9 (Figs. 3(c) and 4(c)). A significant reduction in the intima:media thickness ratio and the percentage of CSN was observed in the C. oil-treated groups when compared with those subjected to PCL or Ox-injury alone (Figs. 3 and (e) and 4).
Qualitative comparison of atherosclerotic lesions after exposure to PCL or Ox-injury revealed that lesions formed after exposure to Ox-injury were rich in lipids and exhibited more stenosis when compared with the PCL-induced group. Moreover, Movat's staining revealed that enhanced extracellular matrix accumulation, rather than cell proliferation, largely contributed to vascular stenosis in the atherosclerotic arteries of the Ox-injury-induced group. The C. oil treatment reduced arterial narrowing in both PCL- and Ox-injury-induced groups.
Curcuma oil prevents partial carotid ligation- and oxidative-injury-induced aortic inflammation
Accumulation of immune cells and inflammation is a hallmark of atherosclerosis. Therefore, we analysed the mRNA transcripts of pro- and anti-inflammatory cytokines in atherosclerotic arteries. Elevated expression of cluster of differentiation 45 (CD45) in the carotid artery was observed at 1, 3 and 5 weeks after exposure to PCL or Ox-injury when compared with the uninjured control group (Fig. 5(a) and (b)). Gene expression analysis of cytokines using real-time RT-PCR showed that the mRNA transcripts of TNF-α, interferon-γ (IFN-γ), IL-1β, IL-6 and IL-10 increased progressively with time after exposure to PCL or Ox-injury when compared with the uninjured control group (Fig. 5(a) and (b)). The mRNA expression of TGF-β was significantly increased at 1, 3 and 5 weeks after exposure to Ox-injury (Fig. 5(b)).

Fig. 5 Effect of curcuma oil (C. oil) on gene expression levels of cytokines in atherosclerotic plaques. mRNA expression of cluster of differentiation 45 (CD45), TNF-α, interferon-γ (IFN-γ), IL-1β, IL-2, IL-6, transforming growth factor-β (TGF-β) and IL-10 in the carotid artery from hamsters at 1, 3 and 5 weeks after exposure to (a) partial carotid ligation (PCL, n 5) and (b) oxidative injury (Ox-injury, n 5). Values are means, with their standard errors represented by vertical bars. Mean value was significantly different from that of the uninjured control group: * P< 0·05, ** P< 0·01 (one-way ANOVA). Mean value was significantly different from that of the high-cholesterol (HC)+PCL-5 weeks or HC+Ox-injury-5 weeks group: † P< 0·05, †† P< 0·01, ††† P< 0·001 (two-tailed, unpaired Student's t test). (a) ![]() , Uninjured control;
, Uninjured control; ![]() , HC+PCL-1 week;
, HC+PCL-1 week; ![]() , HC+PCL-3 weeks;
, HC+PCL-3 weeks; ![]() , HC+PCL-5 weeks;
, HC+PCL-5 weeks; ![]() , HC+PCL-5 weeks+C. oil-300 mg/kg. (b)
, HC+PCL-5 weeks+C. oil-300 mg/kg. (b) ![]() , Uninjured control;
, Uninjured control; ![]() , HC+Ox-inj-1 week;
, HC+Ox-inj-1 week; ![]() , HC+Ox-inj-3 weeks;
, HC+Ox-inj-3 weeks; ![]() , HC+Ox-inj-5 weeks;
, HC+Ox-inj-5 weeks; ![]() , HC+Ox-inj-5 weeks+C. oil-300 mg/kg.
, HC+Ox-inj-5 weeks+C. oil-300 mg/kg.
Treatment with C. oil (300 mg/kg) significantly attenuated the aortic mRNA expression of CD45, TNF-α, IFN-γ, IL-1β and IL-6, while the expression of TGF-β and IL-10 was significantly increased when compared with the groups exposed to PCL or Ox-injury alone (Fig. 5(a) and (b)).
Curcuma oil attenuates peritoneal macrophage foam-cell formation by regulating genes involved in cholesterol homeostasis
To explore the effect of hyperlipidaemia on foam-cell formation in vivo, cellular lipid and CD68 mRNA expression were quantified in macrophages from the peritoneal cavity of the chow- or HC-fed group. Peritoneal macrophages of the HC-fed group showed increased cellular accumulation of TC, FC and CE, indicating cellular lipid enrichment following hyperlipidaemia (Fig. 6(a)). The mRNA expression of CD68 was also elevated in the peritoneal macrophages of the HC-fed group when compared with the chow-fed control group (Fig. 6(b)).

Fig. 6 Curcuma oil (C. oil) attenuates peritoneal macrophage foam-cell formation. Peritoneal macrophages were collected after 5 weeks of high-cholesterol (HC) diet feeding with or without C. oil (300 mg/kg) treatment. (a) Cellular total cholesterol (TC), free cholesterol (FC) and cholesteryl ester (CE) (n 5) and (b) mRNA expression of PPARα, liver X receptor-α (LXRα), ATP-binding cassette A1 (ABCA1), ABCG1, cluster of differentiation 36 (CD36) and CD68 (n 6), and (c) TNF-α, IL-1β, IL-6, interferon-γ (IFN-γ), transforming growth factor-β (TGF-β) and IL-10 (n 6) in the peritoneal macrophages of the chow- or HC-fed group with or without C. oil treatment. Values are means, with their standard errors represented by vertical bars. Mean value was significantly different from that of the chow-fed group: * P< 0·05, ** P< 0·01 (two-tailed, unpaired Student's t test). Mean value was significantly different from that of the chow-fed group: † P< 0·05, †† P< 0·01 (one-way ANOVA). Mean value was significantly different from that of the HC-5 weeks group: ‡ P< 0·05, ‡‡ P< 0·01, ‡‡‡ P< 0·001 (two-tailed, unpaired Student's t test). ![]() , Chow;
, Chow; ![]() , HC-5 weeks;
, HC-5 weeks; ![]() , HC-5 weeks+C. oil-300 mg/kg.
, HC-5 weeks+C. oil-300 mg/kg.
The C. oil (300 mg/kg) treatment significantly reduced the cellular accumulation of TC, FC and CE in peritoneal macrophages (Fig. 6(a)). Moreover, C. oil suppressed the mRNA expression of CD68 in peritoneal macrophages when compared with the HC-fed group (Fig. 6(b)), suggesting that C. oil prevents in vivo foam-cell formation in hyperlipidaemic hamsters.
To explore the possible mechanisms underlying the preventive effect of C. oil on macrophage foam-cell formation, the mRNA expression of lipid-related genes was evaluated. Peritoneal macrophages of the HC-fed group displayed increased mRNA expression of PPARα, liver X receptor-α (LXRα), ATP-binding cassette A1 (ABCA1), ATP-binding cassette G1 (ABCG1) and cluster of differentiation 36 (CD36) when compared with the chow-fed group (Fig. 6(b)).
Furthermore, C. oil (300 mg/kg) treatment in the HC-fed group increased the mRNA expression of PPARα, LXRα, ABCA1 and ABCG1 in peritoneal macrophages when compared with the untreated HC-fed group (Fig. 6(b)). However, the mRNA expression of CD36 was suppressed in the peritoneal macrophages of the C. oil-treated group.
Since treatment with C. oil reduced the gene expression of pro-inflammatory cytokines in atherosclerotic arteries, its effect on the mRNA expression of inflammatory cytokines was evaluated in peritoneal macrophages. Unexpectedly, peritoneal macrophages of the HC-fed group exhibited reduced mRNA transcripts of TNF-α, IL-6 and IFN-γ when compared with the chow-fed group (Fig. 6(c)). The mRNA expression of TGF-β was increased in the HC-fed group, while that of IL-1β and IL-10 remained unchanged (Fig. 6(c)).
C. oil treatment in the HC-fed group further reduced mRNA expression of TNF-α, IL-1β, IL-6 and IFN-γ in the peritoneal macrophages as compared with HC fed alone (Fig. 6(c)). Importantly, C. oil augmented mRNA expression of TGF-β in peritoneal macrophages of the HC-fed group; however, the mRNA expression of IL-10 remained unchanged (Fig. 6(c)).
Curcuma oil prevents oxidised LDL-induced macrophage foam-cell formation and inflammation
To determine whether the preventive effect of C. oil on peritoneal macrophage foam-cell formation and inflammation is due to its direct effect on macrophages or to its lipid-lowering effect, further studies were carried out in the human monocytic cell line THP-1. OxLDL (40 μg/ml) stimulation for 48 h in THP-1 macrophages significantly increased the cellular accumulation of TC, FC and CE after OxLDL treatment (Fig. 7(a)). Furthermore, an increase in the mRNA expression of CD68 was also observed in OxLDL-stimulated macrophages (Fig. 7(b)).

Fig. 7 Curcuma oil (C. oil) inhibits oxidised LDL (OxLDL)-induced macrophage foam-cell formation and inflammation. To evaluate the effect of C. oil on OxLDL-induced cholesterol accumulation, mRNA expression of lipid-related genes, and cytokine production, THP-1 macrophages were pre-incubated with C. oil (1, 3 and 10 μg/ml) for 18 h followed by OxLDL (40 μg/ml) stimulation for 48 h. After the stimulation, the supernatant was collected for cytokine estimation by ELISA, and the cells were processed for complementary DNA synthesis or cholesterol extraction. (a) Cellular total cholesterol (TC), free cholesterol (FC) and cholesteryl ester (CE, n 5), (b) mRNA expression of PPARα, liver X receptor-α (LXRα), ATP-binding cassette A1 (ABCA1), ABCG1, cluster of differentiation 36 (CD36) and CD68 (n 5) and (c) cytokine levels (ng/ml) in the cell-culture supernatant (n 5). Values are means, with their standard errors represented by vertical bars. Mean value was significantly different from that of the THP-1 control macrophages: *** P< 0·001 (two-tailed, unpaired Student's t test). Mean value was significantly different from that of the THP-1 control macrophages: † P< 0·05, †† P< 0·01, ††† P< 0·001 (one-way ANOVA). Mean value was significantly different from that of the THP-1+OxLDL-40 μg/ml group: ‡ P< 0·05, ‡‡ P< 0·01, ‡‡‡ P< 0·001 (two-tailed, unpaired Student's t test). Mean value was significantly different from that of the THP-1+OxLDL-40 μg/ml group: § P< 0·05, §§ P< 0·01, §§§ P< 0·001 (one-way ANOVA). (a) ![]() , THP-1 control;
, THP-1 control; ![]() , THP-1+OxLDL-40 μg/ml;
, THP-1+OxLDL-40 μg/ml; ![]() , THP-1+C. oil-1 μg/ml+OxLDL-40 μg/ml;
, THP-1+C. oil-1 μg/ml+OxLDL-40 μg/ml; ![]() , THP-1+C. oil-3 μg/ml+OxLDL-40 μg/ml;
, THP-1+C. oil-3 μg/ml+OxLDL-40 μg/ml; ![]() , THP-1+C. oil-10 μg/ml+OxLDL-40 μg/ml. (b)
, THP-1+C. oil-10 μg/ml+OxLDL-40 μg/ml. (b) ![]() , THP-1 control;
, THP-1 control; ![]() , THP-1+C. oil-10 μg/ml;
, THP-1+C. oil-10 μg/ml; ![]() , THP-1+OxLDL-40 μg/ml;
, THP-1+OxLDL-40 μg/ml; ![]() , THP-1+C. oil-10 μg/ml+OxLDL-40 μg/ml. (c)
, THP-1+C. oil-10 μg/ml+OxLDL-40 μg/ml. (c) ![]() , THP-1 control;
, THP-1 control; ![]() , THP-1+OxLDL-40 μg/ml;
, THP-1+OxLDL-40 μg/ml; ![]() , THP-1+C. oil-1 μg/ml+OxLDL-40 μg/ml;
, THP-1+C. oil-1 μg/ml+OxLDL-40 μg/ml; ![]() , THP-1+C. oil-3 μg/ml+OxLDL-40 μg/ml;
, THP-1+C. oil-3 μg/ml+OxLDL-40 μg/ml; ![]() , THP-1+C. oil-10 μg/ml+OxLDL-40 μg/ml.
, THP-1+C. oil-10 μg/ml+OxLDL-40 μg/ml.
To evaluate the effect of C. oil on in vitro macrophage foam-cell formation, THP-1 macrophages were pretreated with C. oil (1, 3 and 10 μg/ml) for 18 h followed by OxLDL stimulation for 48 h. Treatment with C. oil (3 and 10 μg/ml) prevented the cellular accumulation of TC, FC and CE in OxLDL-stimulated THP-1 macrophages (Fig. 7(a)). However, the lower dose of C. oil (1 μg/ml) was ineffective in reducing cellular cholesterol content. Pretreatment with C. oil in THP-1 macrophages alone did not alter cellular cholesterol levels (data not shown). C. oil (10 μg/ml) pretreatment in THP-1 macrophages alone or in OxLDL-stimulated cells significantly reduced the mRNA transcript of CD68 when compared with their respective controls (Fig. 7(b)).
To explore the possible mechanisms underlying C. oil-induced reduction in cellular cholesterol accumulation, the mRNA expression of different genes involved in cellular cholesterol uptake and efflux was monitored in THP-1 macrophages with or without C. oil treatment using quantitative RT-PCR.
OxLDL stimulation in THP-1 macrophages significantly increased the mRNA expression of PPARα, LXRα, ABCA1, ABCG1 and CD36 when compared with THP-1 control macrophages (Fig. 7(b)).
Overnight pretreatment with C. oil (10 μg/ml) in THP-1 macrophages significantly elevated the expression of PPARα, LXRα, ABCA1 and ABCG1 when compared with THP-1 control macrophages (Fig. 7(b)). Similarly, in OxLDL-stimulated THP-1 macrophages, pretreatment with C. oil (10 μg/ml) increased the mRNA expression of PPARα, LXRα, ABCA1 and ABCG1 compared with OxLDL alone. The mRNA expression of CD36 was significantly decreased after C. oil treatment in control macrophages as well as in OxLDL-stimulated macrophages when compared with their respective controls.
Furthermore, the effect of C. oil was evaluated on OxLDL-induced inflammation in THP-1 macrophages. OxLDL stimulation for 48 h increased the production of TNF-α and IL-1β in THP-1 macrophages (Fig. 7(c)), while TGF-β and IL-10 remained unchanged when compared with the control macrophages (Fig. 7(c)). Pretreatment with C. oil (1, 3 and 10 μg/ml) in THP-1 macrophages prevented OxLDL-induced TNF-α and IL-1β production in a dose-dependent manner compared with OxLDL alone (Fig. 7(c)). The higher dose of C. oil (10 μg/ml) increased the production of TGF-β in OxLDL-stimulated macrophages when compared with OxLDL alone (Fig. 7(c)). However, the lower doses of C. oil were ineffective in modulating the production of TGF-β. The production of IL-10 remained unchanged in OxLDL-stimulated THP-1 macrophages in the presence or absence of C. oil (Fig. 7(c)). Cytokine production in THP-1 macrophages with or without C. oil pretreatment was similar (data not shown).
Discussion
The purpose of the present study was to explore the anti-atherosclerotic effects and underlying protective mechanisms of C. oil in hamster models of accelerated atherosclerosis and in THP-1 macrophages.
The present results showed that C. oil prevented arterial injury-induced accelerated atherosclerosis and macrophage foam-cell formation. In addition to inhibiting macrophage and lipid accumulation in hamster models, C. oil significantly attenuated OxLDL-induced foam-cell formation and inflammation in THP-1 macrophages. Therefore, in vivo anti-atherosclerotic effect of C. oil can be partly attributed to its direct effect on macrophages.
Since C. oil at the dose of 300 mg/kg effectively lowered lipid levels in a previous study( Reference Singh, Jain and Misra 2 ), this dose was chosen for monitoring anti-atherosclerotic effects in the present study. Ezetimibe has been shown to prevent accelerated atherosclerosis in the rabbit( Reference Gomez-Garre, Munoz-Pacheco and Gonzalez-Rubio 23 ). This finding is consistent with the effects observed in the present study, therefore justifying the use of ezetimibe in the validation of anti-atherosclerotic models.
Ease of access to the common carotid artery and its branches allows for the induction of vascular injury that accelerates the development of atherosclerosis. Disturbed flow with low and oscillatory shear stress induced by PCL leads to rapid endothelial dysfunction recruitment of leucocytes including macrophages, and atherosclerosis( Reference Nam, Ni and Rezvan 13 ). Similarly, we observed that PCL in hyperlipidaemic hamsters accelerated aortic lipid and macrophage foam-cell accumulation and vascular proliferation. In contrast, FeCl3 induces endothelial injury. The adaptive response to this injury involves infiltration of leucocytes, lipid accumulation, and enhanced neointimal and medial fibrosis( Reference Tian, Hu and Sun 12 ). Movat's pentachrome staining revealed that muscle content primarily accounted for PCL-induced arterial stenosis. In contrast, extracellular matrix accumulation largely contributed to arterial stenosis after exposure to Ox-injury. Moreover, the early increase in the relative mRNA expression of collagen-1 and collagen-3 indicates its contribution to the progression of atherosclerosis after exposure to Ox-injury. However, few collagen-stained areas were observed in the intimal portion of the carotid artery exposed to PCL. The dissimilarity observed between the atherosclerotic lesions could be due to the difference in the type of arterial injury: PCL activates vascular endothelium, while FeCl3-induced Ox-injury denudes it. Following Ox-injury to the carotid artery, relatively more fold induction in the mRNA expression of pro-atherogenic cytokines was observed in comparison to the PCL-induced group. This may also explain the enormous vascular stenosis observed in the Ox-injury-induced group.
In contrast to hamsters, apoE− / − mice has been shown to develop rapid atherosclerotic plaque that comprises enormous aortic lipids, inflammation and intra-plaque neovessels following exposure to PCL( Reference Nam, Ni and Rezvan 13 ). The characteristic difference in atherosclerotic lesions in these two species may be associated with the difference in physiological characteristics such as mean arterial pressure and heart rate, which regulate the level of oscillatory shear stress exerted on the arterial wall after exposure to PCL( Reference Mattson 24 , Reference Thomas, Artwohl and Suzuki 25 ).
Early increases in the mRNA expression of CD45 in response to both PCL and Ox-injury suggest that vascular injury triggers leucocyte infiltration, which can substantially contribute to vascular inflammation( Reference Alberts-Grill, Rezvan and Son 26 ). The inhibitory effect of C. oil on the aortic expression of CD45, as well as TNF-α, IFN-γ, IL-6 and IL-1β, may also contribute to its anti-atherosclerotic effect. Curcumene, a component of C. oil, is known to inhibit leucocyte migration( Reference Nogueira de Melo, Grespan and Fonseca 27 ), which might explain the reduced aortic expression of the leucocyte marker CD45 in the C. oil-treated group.
The aortic expression of MMP-2 and MMP-9 was increased in the later stages of atherosclerosis progression. As reported previously, MMP-2 and MMP-9 are mainly expressed in advanced atherosclerotic plaque in a mouse model( Reference Wagsater, Zhu and Bjorkegren 28 ), and have been shown to induce carotid plaque instability( Reference Toutouzas, Synetos and Nikolaou 29 ).
The inhibitory effect of C. oil on aortic MMP and pro-atherogenic cytokines suggests that it may reduce the propensity of the rupture of atherosclerotic plaque; however, this hypothesis needs to be tested in experimental animal models.
It is widely known that macrophages are abundantly present in the peritoneal cavity of rodents( Reference Ghosn, Cassado and Govoni 30 ). In the present study, we observed that peritoneal macrophages accumulated significant amounts of cellular cholesterol mainly in the form of CE in response to hyperlipidaemia. This is quite similar to atherosclerotic lesions where CE mainly contribute to the foamy appearance of macrophages and to plaque vulnerability( Reference Chen, Ichetovkin and Kurtz 31 ). C. oil attenuated hyperlipidaemic-induced peritoneal macrophage lipid accumulation and enhanced the expression of lipid-related genes involved in lipid metabolism and efflux. Therefore, reduced lipid accumulation in peritoneal macrophages may be attributed to both plasma lipid-lowering and enhanced mRNA expression of genes involved in cholesterol metabolism and efflux. To further test whether C. oil can directly inhibit macrophage lipid accumulation, experiments were carried out in THP-1 cells.
In THP-1 macrophages, C. oil exhibited dose-dependent inhibition of OxLDL-induced cholesterol accumulation and enhanced the mRNA expression of lipid-related genes involved in cellular cholesterol metabolism and efflux. More importantly, C. oil reduced the expression of the scavenger receptor CD36 in both peritoneal and THP-1 macrophages. Consequently, it can be speculated that C. oil prevents lipid accumulation in macrophages by regulating the expression of these genes. The overexpression of PPARα, LXRα, ABCA1 and ABCG1 in macrophages has been shown to prevent the development of atherosclerosis( Reference Rigamonti, Chinetti-Gbaguidi and Staels 32 , Reference Li, Binder and Gutierrez 33 ), suggesting that these genes may be involved in decreased atherosclerotic plaque formation in C. oil-treated hamsters. Furthermore, C. oil inhibited the OxLDL-induced secretion of pro-inflammatory cytokines in a dose-dependent manner, thus demonstrating its anti-inflammatory effect. Although an increase in the expression of TGF-β may confer some beneficial effects in atherosclerosis due to its anti-proliferative and apoptotic effects on fibrotic cells( Reference Toma and McCaffrey 34 ), it can also affect cell migration, matrix synthesis, wound contraction, calcification and immune response( Reference Toma and McCaffrey 34 ). In the present study, the protective effect of TGF-β might be explained by the modulation of such pathways. However, the ultimate effect of TGF-β probably depends on the microenvironment, ageing and disease progression, since young and old cells respond differently to TGF-β ( Reference Toma and McCaffrey 34 ). In contrast, peritoneal macrophages from hyperlipidaemic hamsters exhibited reduced expression of pro-atherogenic cytokines, a phenomenon that may be explained by the accumulation of desmosterol( Reference Spann, Garmire and McDonald 35 ).
The present study aimed to explore the preventive effect of C. oil as the treatment was started before the manifestation of the disease. The preventive regimen employed in the present study may offer some insight, albeit speculated, into the possible cardiovascular benefits to individuals who consume curcumin in their daily diet. However, the precise preventative effect of C. oil can only be ascertained in the models of atherosclerosis regression.
Extreme caution should be exercised while deriving exact human doses by using mathematical formulas( Reference Spann, Garmire and McDonald 35 ). According to a commonly used body surface area-based dose calculation method( Reference Reagan-Shaw, Nihal and Ahmad 36 ), C. oil doses of 100 and 300 mg/kg in hamsters will be equivalent to doses of about 800 mg and 2·4 g, respectively, for a human adult( Reference Reagan-Shaw, Nihal and Ahmad 36 ). Although the preparation of C. oil has not currently been tested in human subjects, turmeric oil (600 mg/d) with turmeric (3 g/d) has been shown to exert a beneficial effect in patients with oral submucous fibrosis in a previous study( Reference Hastak, Lubri and Jakhi 37 ). In another human study, intake of turmeric oil at doses of 600 mg and 1 g/d for 1 and 3 months, respectively, has been found to be safe by using haematological, renal and hepatotoxicity parameters( Reference Joshi, Ghaisas and Vaidya 38 ). In a previous study in hamsters, C. oil has not been shown to exhibit hepatotoxicity at the highest dose (300 mg/kg)( Reference Singh, Jain and Misra 2 ), and, in fact, it has been shown to exhibit beneficial effects as reflected by the improvement in liver function and oxidative stress( Reference Singh, Jain and Misra 2 ). Dietary surveys in the Asian population have revealed a positive correlation between improved cognitive function and decreased incidence of cancer with the regular dietary intake of turmeric for extended periods of time( Reference Ng, Chiam and Lee 39 , Reference Hutchins-Wolfbrandt and Mistry 40 ). However, controlled trials have yet to establish a proven correlation. Since the percentage of C. oil is only 1·4 % in C. longa, it will require a daily intake of at least 25 g of turmeric in order to achieve a dose of 300 mg/kg. Therefore, it is quite possible that a regular intake of C. oil in humans at a similar or lower dose for a longer duration might produce anti-atherosclerotic effects. However, a long-term study with lower doses of C. oil in animals, accompanied by detailed toxicity and safety evaluations, must be conducted to support the translation of C. oil for human use.
Acknowledgements
The authors gratefully acknowledge the CSIR, New Delhi, India for the award of research fellowships to V. S., and the Indian Council of Medical Research, New Delhi, India to M. R. and M. J. The authors greatly appreciate Mr P. K. Srivastava, Medicinal and Process Chemistry Division, for providing support for the preparation of C. oil. The authors are grateful to Dr M. P. S. Negi, Biometry and Statistics Division, CSIR-CDRI, for helping with the statistical analysis of the data.
The present study was financially supported by the CSIR, New Delhi, India and the Network Project BSC0102 and BSC0103.
The authors’ contributions are as follows: V. S. contributed to the planning and execution of the experimental animals; V. S. performed the surgery and lipid profiling and gene expression studies; M. R. and V. S. carried out aortic and cellular cholesterol estimations; M. R. was responsible for OxLDL preparation and treatments in THP-1 cells; M. J. performed RNA isolation and cDNA synthesis; N. S. and V. S. carried out Movat's pentachrome and Picrosirius Red staining; R. M., A. N. and A. K. D. were responsible for the preparation, isolation and characterisation of the C. oil extract and helped in the data analysis and writing of the manuscript; M. D. provided critical inputs for the experiments; M. K. B. was responsible for the planning, execution and troubleshooting of the experiments and the preparation of the manuscript.
The present paper is CSIR-CDRI communication no. 8827.
The authors declare that there are no conflicts of interest.








