


Dietary microparticles are defined as non-biological particles of the gastrointestinal lumen that are of appropriate size and shape (typically 0·2–1·0 μm diameter), for uptake by mucosal phagocytes, in much the same way as biological particles (i.e. bacteria) can be. It should be more widely appreciated that (i) two distinct types of microparticle exist, namely endogenous and exogenousReference Lomer, Thompson and Powell1 (ii) gastrointestinal exposure is substantial (e.g. 10Reference Sernka and Borle12 exogenous particles per person per day in the UK)Reference Lomer, Hutchinson, Volkert, Greenfield, Catterall, Thompson and Powell2 (iii) specific mechanisms exist in the gut for the absorption of non-biological particles so tissue uptake is also largeReference Powell, Ainley, Harvey, Mason, Kendall, Sankey, Dhillon and Thompson3 and (iv) such particles are most likely to adsorb or trap soluble molecules of the gut lumen so that cells are exposed to “conjugates” of non-biological particle plus associated biomoleculesReference Ellingsen4–Reference Van der Meer, Welberg, Kuipers, Kleibeuker, Mulder, Termont, Vonk, De Vries and De Vries7. Over the past ten years our group has aimed to characterise these particles and to define their origins, route of gastrointestinal uptake, cellular fate and immuno-modulatory properties. We have also sought to test the hypothesis that microparticles are involved in the aetiopathogenesis of Crohn's disease. Here, we briefly overview exposure to and fate of dietary microparticles and then concentrate on what is known of their immune modulating properties.
Exogenous or Endogenous?
During early work, aimed at characterising the chemical behaviour of ingested metals in the gut lumen, Powell and colleagues noted that classically insoluble metal ions at intestinal pH (e.g. iron, aluminium, etc) were in fact rendered soluble by factors of the intestinal juices (ligands and mucin) whereas calcium, uniquely, was found in great abundance co-precipitated with phosphateReference Powell, Whitehead, Ainley, Kendall, Nicholson and Thompson8. These calcium phosphate precipitates had been well reported beforeReference Schedl, Osbaldiston and Mills9, Reference Favus10, but, what had not been well appreciated, is that a fraction of the precipitate is reasonably homogenous sub-micron sized microparticles-making these candidates for gut tissue uptakeReference Powell, Ainley, Harvey, Mason, Kendall, Sankey, Dhillon and Thompson3, Reference Sass, Dreyer and Seifert11. In fact the same group had already reported that when similarly sized particles are ingested, a proportion is taken up by the intestinal mucosaReference Powell, Ainley, Harvey, Mason, Kendall, Sankey, Dhillon and Thompson3. So, why might the gut “manufacture” microparticles of calcium phosphate in the lumen (i.e. endogenous particles) that could then be scavenged by mucosal phagocytes? And what would the impact be of exposing this same system to new, man-made microparticles (i.e. exogenous particles)?
Even on calcium and phosphorus-poor diets, active secretion of Ca2+ and PO43 − ions, from the intestinal mucosa and into the lumen, still occurs and co-precipitation ensuesReference Schedl, Osbaldiston and Mills9, Reference Favus10, Reference Sernka and Borle12. In spite of the observation that calcium phosphate microparticles (generally less than 1 μm) form in the gastrointestinal lumen, their translocation into the intestinal mucosa is yet to be shown. This is mainly due to technical challenges in preserving and observing freshly precipitated and thus labile, mineral structures in situ. Nonetheless, their presence can be invoked from the ubiquitous nature of calcium phosphate microparticles in the lumen and the exceptional capacity of M cells, which chiefly overlay intestinal lymphoid aggregates (Peyer's patches in the small bowel), in the uptake of non-biological particlesReference Sass, Dreyer and Seifert11, Reference Pappo and Ermak13, Reference Smith, Thomas, Jenkins, Miller, Cremaschi and Porta14.
Endogenous microparticles
Our interest lies in the biological role of luminally precipitated calcium phosphate in the gut and, specifically, in the potential immunological benefit(s) of the microparticulate fraction. Calcium phosphate, due to its surface properties, is able to adsorb organic molecules and/or luminal componentsReference Govers, Termont, Van Aken and Van der Meer5–Reference Van der Meer, Welberg, Kuipers, Kleibeuker, Mulder, Termont, Vonk, De Vries and De Vries7 and therefore may play a role in luminal detoxification (through adsorption of faecal toxins). On quantitative grounds, however, a detoxification role seems unlikely for the microparticulate fraction. Instead, we recently proposed that calcium and phosphate ions may co-precipitate with luminal antigen forming a microparticle sized hydroxy-apatite-antigen complex which is then scavenged by M cells of Peyer's patches and passed to underlying phagocytic lymphoid cells for processingReference Pele, Powell, Colombel, Gasche, Scholmerich and Vucelic15. The presumption that the particles are hydroxy-apatite, rather than any other form of calcium phosphate, is based mainly upon in vitro models and needs to be carefully tested in vivo. If this mechanism is proven then we suggest the term “constitutive antigen transfection”. This could represent a mechanism of immuno-surveillance by continual sampling of gut luminal contents and may lead to promotion of immunological tolerance.
How may calcium phosphate microparticles promote an immuno-tolerant response? It is now well recognised that dietary calcium, possibly through binding to luminal free fatty acids and bile acids, may confer protection against colonic carcinogenesisReference Govers, Termont, Van Aken and Van der Meer5–Reference Van der Meer, Welberg, Kuipers, Kleibeuker, Mulder, Termont, Vonk, De Vries and De Vries7, Reference Lupton, Steinbach and Chang16. However, it has also recently been shown that dietary calcium modulates enteric infection and inflammationReference Bovee-Oudenhoven, Lettink-Wissink, Van Doesburg, Witteman and Van Der Meer17–Reference Bovee-Oudenhoven, Wissink, Wouters and Van der Meer19, which is not easily explained through the binding of bile acids or fatty acids. Other studies have since provided convincing explanations. High levels of dietary calcium have been shown to increase apoptosis of the murine mid and distal colonic epithelial cellsReference Hambly, Saunders, Rijken and Rowland20, Reference Penman, Liang, Bode, Eastwood and Arends21 and to protect against intestinal weight loss and peripheral TNF-α production in an induced-colitis modelReference Zhu, Mahon, Froicu and Cantorna22. In agreement with these findings, we also demonstrated in a recent study that low dietary calcium levels modulate mucosal caspase expression and increase disease activity in mice with dextran sulfate sodium-induced colitis (unpublished results). Thus (by analogy to dietary calcium effects on intestinal epithelial cells), the first possible effect of calcium phosphate microparticles on intestinal mononuclear phagocytes is the induction of apoptosis. Indeed previous data indicate that luminal calcium levels of the gut are proportional to dietary levelsReference Lomer, Thompson and Powell1 and that the uptake of calcium phosphate microparticles by intestinal mononuclear cells can directly trigger cell deathReference Evans, Ashwood, Warley, Berisha, Thompson and Powell23. Thereafter tolerance could be established towards exogenous antigen contained within the apoptotic cells, as this would now be regarded as “self antigen”Reference Ferguson, Herndon, Elzey, Griffith, Schoenberger and Green24–Reference Blander and Medzhitov26. A second possibility is that calcium phosphate-antigen conjugates are taken up by mononuclear cells and the calcium phosphate acts as an adjuvant, promoting antigen presentation, while the intestinal mononuclear cell provides tolerogenic signals as these cells are known to be hyporesponsive compared to similar peripheral cellsReference Smythies, Sellers, Clements, Mosteller-Barnum, Meng, Benjamin, Orenstein and Smith27. A third possibility is that the calcium phosphate microparticle directly induces tolerogenic signalling in the antigen-presenting cell through an unknown mechanism. It is now important that this putative pathway of constitutive antigen transfection is established and that the role of luminal calcium phosphate microparticles is delineated.
Exogenous microparticles
Considerably more is known of exogenous microparticles than it is of endogenous ones. Oral exposure has been reported by Lomer et al Reference Lomer, Hutchinson, Volkert, Greenfield, Catterall, Thompson and Powell2, and amounts to approximately 40 mg/person/day (or about 10Reference Sernka and Borle12 particles) in the UK. Uptake by the Peyer's patch phagocytes has been shown by a number of groupsReference Shepherd, Crocker, Smith and Levison28, Reference Urbanski, Arsenault, Green and Haber29 and best characterised by Powell et al Reference Powell, Ainley, Harvey, Mason, Kendall, Sankey, Dhillon and Thompson3. In particular these particles are sub-micron sized and highly resistant to chemical breakdown so, following ingestion and mucosal uptake, they survive both gastrointestinal digestion and cellular processing. They appear to be almost exclusively additives to food, pharmaceuticals and toothpaste and are chiefly aluminosilicates and titanium dioxide, although a small proportion of non-aluminium containing silicates are also found in human gut tissueReference Powell, Ainley, Harvey, Mason, Kendall, Sankey, Dhillon and Thompson3, Reference Mazumder, Bode, McIntyre and Ghosh30. The distribution of intakes in the UK is shown in Table 1.
Table 1 *Mean total daily intakes per person in the UK of microparticulate silicates (including aluminosilicates) and titanium dioxide derived from food, pharmaceuticals and toothpaste
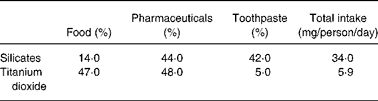
* Data from Lomer et al. 2
In human gut tissue their dominant occurrence is in basal phagocytes of the Peyer's patch (Fig. 1) where they occur in large numbers.

Fig. 1 Dark field microscopy of human small bowel showing an area (in the white box) of particle-containing cells or “pigment cells” (each cell appearing bright white due to reflectance by the pigment in dark field microscopy) located at the base of a human Peyer's Patch (original × 200).
We hypothesize that exogenous microparticles “hijack” the putative route for endogenous microparticle uptake but, due to their chemically-resistant nature, they cannot then contribute to tolerance induction and may even, in susceptible individuals, establish abnormal immune responsivenessReference Lomer, Thompson and Powell1, Reference Powell, Harvey and Thompson31. Based upon the capacity of exogenous microparticles for surface adsorption of organic molecules we also anticipate that conjugation of luminal components will occur in the gastrointestinal tract prior to uptake and that this will influence cell responsivenessReference Ashwood, Thompson and Powell32.
Luminal conjugation
One important aspect of the gastrointestinal tract is the extent of its secretions and bacterial burden. Secretions are rich in endogenous proteins including immunoglobulins, cations and anions, mucin and other glycoproteins, amino acids and short chain fatty acids. In addition, secretions contain whole and partially digested bacteria from the symbiotic flora and some non-absorbed food antigens. The bathing of microparticles in such complex fluids surely leads to adsorptive interactions: these particles, which typically have charged, large-area surfaces, could adsorb luminal constituents, facilitated by both ionic and entropic forces. In particular, cations such as calcium can provide bridging charges for larger proteins and glycoproteins to absorb to the surface of particles Reference Govers, Termont, Van Aken and Van der Meer5–Reference Van der Meer, Welberg, Kuipers, Kleibeuker, Mulder, Termont, Vonk, De Vries and De Vries7. In model systems we have been able to show that adsorption of a typical luminal constituent (namely bacterial lipopolysaccharide, LPS) by exogenous microparticles (either titanium dioxide or aluminosilicates) occurs in the presence of sufficient calcium ionsReference Evans, Ashwood, Warley, Berisha, Thompson and Powell23, Reference Ashwood, Thompson and Powell32, Reference Powell, Harvey, Ashwood, Wolstencroft, Gershwin and Thompson33. Calcium ions, present in the distal gut lumen, would therefore provide excellent bridging potential for components such as LPS or peptidoglycansaccharide (PGPS), to adsorb to ingested microparticles. It is likely that these LPS-calcium-microparticle conjugates are not all pure single particle entities because calcium itself may partially precipitate and complex aggregates may then formReference Ashwood, Thompson and Powell32. Nonetheless the aggregates still fall well within the microparticle range and are avidly taken up by phagocytic cellsReference Ashwood, Thompson and Powell32. Further work is needed to (a) incubate such particles with real intestinal secretions and gut contents to see which materials are preferentially adsorbed and (b) to look in vivo and confirm this model although, analytically, this is a significant challenge. Nonetheless, it seems most likely that ingested dietary microparticles will adsorb certain luminal materials and, as such, be able to carry them into the intestinal tissue via the M-cell portal in this conjugated fashion. Our subsequent work has investigated what the cellular responses are to native particles or conjugated particles.
Dietary microparticles and cellular activity
Certain bacterial motifs that are specifically recognised by cells of the immune system are referred to as pathogen associated molecular patterns (PAMPs) and include, for example, LPS (recognised by toll like receptor 4 (TLR 4)), bacterial flagellin (recognised by TLR 5) and PGPS which contains motifs that are recognised by a number of receptors including the intracellular nuclear oligomerisation domains (NODS). NOD2 for example recognises muramyl dipeptide while NOD1 recognises MurNAc-L-Ala-D-Glu-meso-diaminopimelic acidReference Netea, Ferwerda and de Jong34. Recognition involves specific ligation and cellular responsiveness which typically is an increase in the secretion of pro- inflammatory cytokines such as IL-8, IL-6, TNF-α and IL-1-β coupled with potential changes to cell functions such as antigen presentation and maturation/partial differentiation. Recently Blander et al have shown that dendritic cell uptake of antigen-conjugated particles, co-delivered with LPS (i.e. within the same phagolysosomes), led to presentation of the antigen at the cell surface for induction of T cell proliferative responsesReference Blander and Medzhitov26. In contrast, in the absence of LPS from this specific compartment, the antigen was not presented. Other PAMPs were not investigated but such work provides a theoretical basis for the immune system to differentiate between “inert particles” and biologically active particles such as invasive bacteria. Moreover, it provides a basis for distinction between particulate self-antigen such as apoptotic cells and particulate bacterial antigen such as in early infection. Thus the immune system can determine, based on recognition of these PAMPs, whether to mount a response through T cell activation or not. We thus recognise the importance in understanding how particles and PAMPs engage with the acquired immune system in our work but have so far restricted our studies to outcomes of the innate immune system (i.e. cytokines). The following is a brief review of these works.
In initial work we used intestinal organ cultures (human gut biopsies in culture) as a model for assessment of dietary microparticle effects on stimulation of the pro-inflammatory cytokine IL1-βReference Powell, Harvey, Ashwood, Wolstencroft, Gershwin and Thompson33. Microparticles alone (titanium dioxide) had no effect, neither did high dose LPS. It is important to recognize that immune cells of the gut are hypo-responsive to LPS as indeed they are to PAMPs in general from what is understoodReference Smythies, Sellers, Clements, Mosteller-Barnum, Meng, Benjamin, Orenstein and Smith27. So unlike with peripheral cells, for example, this lack of gut cell responsiveness to LPS is expected. In contrast when LPS and dietary microparticles were added together in the presence of excess calcium, a small (1·5 to 4 fold) increase in IL1-β secretion was observed. The same pattern was observed using peripheral blood mononuclear cells except this time the increase was dramatic with the LPS-calcium-titanium dioxide conjugate (30 to 60 fold above base line)Reference Ashwood, Thompson and Powell32. While this again emphasizes the tolerant nature of intestinal cells it also demonstrates that such particles may have immune potentiating possibilities when bound with PAMPs from the gastrointestinal lumen. Further work really needs to dissect out the effects of calcium in the system because we have subsequently shown that this is an important, even major, part of the activity of the conjugate/agglomerateReference Evans, Ashwood, Warley, Berisha, Thompson and Powell23. Nonetheless this is still likely to represent in vivo situations.
There are a number of problems with organ cultures of colonic biopsies including viability and surface area for full exposure in particle-based studies. Thus, subsequently, we continued this work using isolated human intestinal cells from the lamina propria. We also undertook time course studies. The results were very similar in that there was a small but significant immune potentiating effect of the conjugates versus the individual agents, namely titanium dioxide and lipopolysaccharide aloneReference Evans, Ashwood, Warley, Berisha, Thompson and Powell23. However, the importance of the calcium was especially emphasized because this alone was able to induce some significant secretion of IL1-β, albeit not as protracted as for the conjugate. Overall, the effects were associated with the induction of cell death in the phagocytic population suggesting that caspase activity had been increasedReference Evans, Ashwood, Warley, Berisha, Thompson and Powell23, Reference Ashwood, Thompson and Powell32. Caspase 1, for example, is required for the cleavage of Pro IL1-β and using inhibition-based studies we have been able to show that activation of this caspase was required for the IL1-β release. Moreover, unlike in the previous work with organ cultures, we were able to show significant differences in the response of the cells from patients with inflammatory bowel disease (IBD) versus controls. This was clearly related to the baseline inflammatory status of the patients and probably was not therefore a specific hyper-responsiveness in IBD. It is unclear why this was not observed in the organ culture work but may represent the different genotypes (which were not tested in either works) that were present in the relatively small populations tested or due to the different techniques used.
In further studies we were able to confirm that the results described above were a specific effect of particulate uptake, which was mediated via the scavenger receptor, and was associated with increases in secretion of a large number of pro-inflammatory cytokines albeit only with IL-1β did we see a synergistic effect between particle plus calcium plus LPS compared to the additive effect of the individual componentsReference Evans, Ashwood, Warley, Berisha, Thompson and Powell23, Reference Ashwood, Thompson and Powell32. These data suggested to us that such ingested microparticle-conjugates/agglomerates may exacerbate inflammation in the permeable and hyper responsive gut of subjects with inflammatory bowel disease.
In an initial small pilot study we were able to show that withdrawal of microparticles from the diet of patients with Crohn's disease was associated with marked improvement of symptomsReference Lomer, Harvey, Evans, Thompson and Powell35. This stimulated a large multi-centre double blind trial, which was carefully undertaken. In this work however we were able to show no effect of microparticle withdrawal on disease activity or objective biochemical measures of diseaseReference Lomer, Grainger and Ede36. Patients had to be concomitantly treated with immunosuppressant drugs (corticosteroids in this case) so the differences observed were between corticosteroids plus the normal microparticle-containing diet versus corticosteroids plus a low microparticle diet. It is possible that the corticosteroids masked any differences but, in which case, the differences must have been subtle. A second problem was that despite careful choice of drugs and dietary advice, patients were still being exposed to some microparticles and it may be that a microparticle-free diet, rather than a low microparticle diet, is what is required. However, the more likely interpretation is that microparticles do not significantly influence the pathogenesis of Crohn's disease. Whether they are involved as aetiological factors in triggering the disease is a different matter and something we continue to investigate.
Our further work will look at the effects of microparticles on the acquired immune system and in patients of different genotypic status- for example, in those carrying NOD2 variants. We cannot get away from the fact that Crohn's disease is a modern, Western disease, the first signs of which are seen overlying intestinal lymphoid aggregates and that there is clearly a gene-environment interaction in the triggering of the disorder. The presence of these microparticles in the modern, Western diet, and their uptake and large quantitative presence within the intestinal lymphoid aggregates of the gut still makes them tantalizing candidates for involvement in this complex polygenic disorder. Evidence however remains elusive.
Conflict of interest statement
The authors have no conflict of interest to report. The authors contributed equally in the preparation and redaction of the manuscript.


