



Non-Technical Summary
Distinctive changes in carbon isotope curves are used extensively alongside trilobite faunal turnover in the international correlation of Cambrian strata. One such isotopic signature, called “SPICE” (Steptoean Positive Isotope Carbon Excursion), is widely used, but in North America, the co-occurring trilobite fossils have never been illustrated. We here describe, discuss, and illustrate the 34 trilobite species (two new) that occur below, within, and above the SPICE from the same section where the carbon isotope data were collected in Utah. The illustration of the specimens, rather than just listing taxa, allows other scientists to evaluate the conclusions made here: the SPICE began in the Aphelaspis Biochron and ended within the Elvinia Biochron.
Introduction
The Steptoean Positive Isotope Carbon Excursion (SPICE) is an event broadly used for correlation of the Furongian Series worldwide (Saltzman et al., Reference Saltzman, Cowan, Runkel, Runnegar, Stewart and Palmer2004; Geyer, Reference Geyer2019; Peng et al., Reference Peng, Babcock, Ahlberg, Gradstein, Ogg, Schmitz and Ogg2020). The strata containing the SPICE record in northern Utah provides a continuous high-resolution δ13C stratigraphy for the upper Miaolingian to Furongian series along with a detailed trilobite biostratigraphy and U–Pb calibrated maximum depositional ages of <494 Ma (Cothren et al., Reference Cothren, Farrell, Sundberg, Dehler and Schmitz2022). This combination of features makes this SPICE record in northern Utah probably one of the most important SPICE records in Laurentia and worldwide and allows for testing ideas about correlation, biologic evolution, and the Cambrian timescale.
The Smithfield Canyon section of northern Utah is one of the fundamental locations used to establish the SPICE as an event for international correlation (Saltzman et al., Reference Saltzman, Cowan, Runkel, Runnegar, Stewart and Palmer2004; Geyer, Reference Geyer2019; Peng et al., Reference Peng, Babcock, Ahlberg, Gradstein, Ogg, Schmitz and Ogg2020), and the trilobites from this section are important for establishing the Laurentian biostratigraphic range of the SPICE and its international correlation. The base of the international Furongian, Paibian Stage, is determined by the first appearance of the Glyptagnostus reticulatus (Kobayashi, Reference Kobayashi1938) in Paibi, China. Biostratigraphic correlation of the base of the Furongian to Laurentia is based on the first appearance of G. reticulatus, which co-occurs with Aphelaspis fauna at the base of the Steptoean Stage of Laurentia (Palmer Reference Palmer1962; Peng et al., Reference Peng, Babcock, Robison, Lin, Rees and Saltzman2004). However, agnostids, which are crucial in international correlations (Peng et al., Reference Peng, Babcock, Ahlberg, Gradstein, Ogg, Schmitz and Ogg2020), are essentially absent from the lower part of the Smithfield Canyon section; thus, correlations require the use of polymeroid trilobites, which are less cosmopolitan.
The trilobites used for the original biostratigraphy in Smithfield Canyon were identified by Allison Palmer in Saltzman et al. (Reference Saltzman, Cowan, Runkel, Runnegar, Stewart and Palmer2004), but unfortunately they were never illustrated to allow re-evaluation of the identifications. The recent high-resolution δ13C stratigraphic study by Cothren et al. (Reference Cothren, Farrell, Sundberg, Dehler and Schmitz2022) re-documented the SPICE at this section and calibrated it using high-precision chemical abrasion–isotope dilution thermal ionization mass spectrometry (CA-ID-TIMS) of detrital zircons, yielding maximum depositional ages (MDAs) of the excursion and additional trilobite collections with preliminary results.
The goal of this study is to present the details of the trilobite paleontology of the new material collected for this geochemical analysis and provide refinement of the biostratigraphy originally reported by Saltzman et al. (Reference Saltzman, Cowan, Runkel, Runnegar, Stewart and Palmer2004). A parallel goal is to present new δ13C chemostratigraphy from a measured section that overlaps with the SPICE section from Cothren et al. (Reference Cothren, Farrell, Sundberg, Dehler and Schmitz2022) to better identify the end of the SPICE.
Geologic background
Regional geology of the Nounan and St. Charles formations
The Cambrian (Marjuman–Sunwaptan) Nounan and St. Charles formations are part of a thick Ediacaran–Devonian stratal package exposed in the Bear River Range of southeastern Idaho and northern Utah (e.g., Maxey, Reference Maxey1958; Fig. 1.1). The St. Charles Formation (300 m thick) is divided into the basal ~20 m thick, siliciclastic-rich Worm Creek Member and overlying informal upper carbonate member. In the northernmost exposures of these strata, in southeastern Idaho, the Worm Creek Member and the underlying Nounan Dolomite (300 m thick) contain two to four laterally discontinuous, overall upward-fining cycles of feldspathic arenite that transition to micrite and dolomicrite, wackestone, and packstone (Wakeley, Reference Wakeley1975; Todt, Reference Todt2014; Link et al., Reference Link, Todt, Pearson and Thomas2017). The upper Nounan and lower St. Charles formations are regionally correlative to the upper Frenchman Mountain Dolostone, Dunderberg, and Nopah formations in the central Great Basin, Utah, Nevada, and Arizona (Rowland and Korolev, Reference Rowland, Korolev, Hollingsworth, Sundberg and Foster2011; Karlstrom et al., Reference Karlstrom, Mohr, Schmitz, Sundberg, Rowland, Blakey and Hagadorn2020; Rowland et al., Reference Rowland, Korolev, Hagadorn and Ghosh2023); the Bonanza King and Nopah Formations in Death Valley, CA (Montañez et al., Reference Montañez, Osleger, Banner, Mack and Musgrove2000); and the Orr Formation in the House Range, Utah (Baker, Reference Baker2010; Fig. 2).
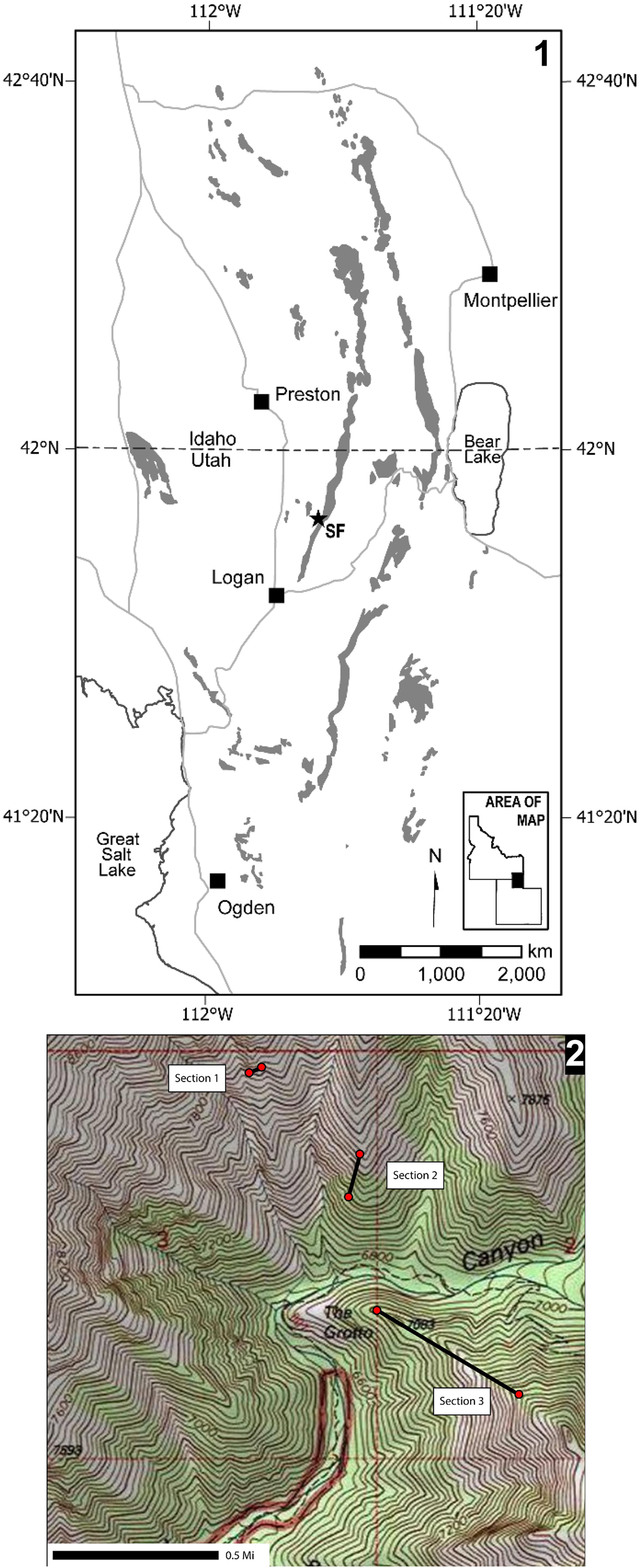
Figure 1. (1) Map showing the location of the Smithfield Canyon section (black star, “SF”) in regional context. Gray polygons represent the aerial extent of mapped Nounan and St. Charles formations. Modified from the Stage Geologic Map Compilation (Horton et al., Reference Horton, San Juan and Stoeser2017) and Wakeley (Reference Wakeley1975). (2) Portion of the Naomi Peak 7.5-foot quadrangle showing the locations of the three measured sections. Basemap: 2013 National Geographic USA topographic map, projection: NAD 1983 UTM Zone 12N.
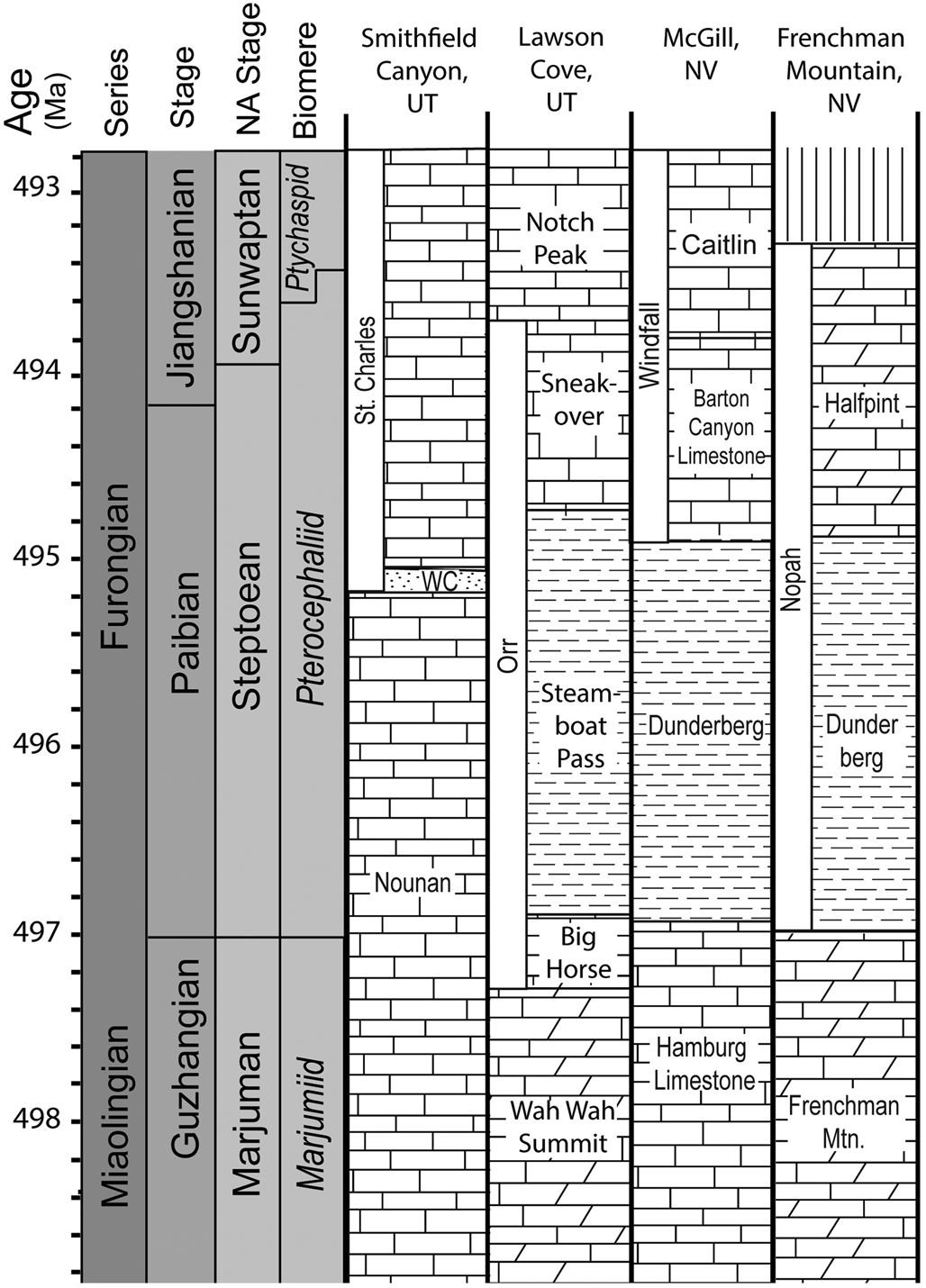
Figure 2. Working hypothesis for the correlation of the Nounan and St. Charles formations in northern Utah with formations in the Great Basin. Approximated biomere boundaries and their corresponding estimated time boundaries. NA = North American; WC = Worm Creek Member; Mtn = Mountain.
While for much of western Laurentia, the late Cambrian is a period of putative tectonic quiescence, this region experienced dynamic magmatism and uplift. A combination of normal faulting and thermal subsidence south (current geography) of the dextral-normal Snake River Transfer Fault created the Worm Creek depocenter (Lund, Reference Lund2008), which received clastic detritus from Cambrian plutons exhumed by uplift of the Lemhi Arch (Link et al., Reference Link, Todt, Pearson and Thomas2017). Quartzites of the Worm Creek Member are interpreted to represent the influx of siliciclastic sediment at the second-order Sauk II–III transition maximum lowstand within the greater transgressive regime across Laurentia during Cambrian time (Sloss, Reference Sloss1963; Saltzman et al., Reference Saltzman, Cowan, Runkel, Runnegar, Stewart and Palmer2004).
Smithfield Canyon, Utah
In Smithfield Canyon, the upper 160 m of the Nounan Formation is characterized by digitate stromatolites, thrombolites (including a prominent 7 m stacked thrombolite complex), sandy trilobite grainstones, imbricated intraclastic conglomerates, oolitic packstone to grainstones, and a 0.25 m thick calcareous sandstone horizon (Fig. 3). Hummocky cross-stratified to swaley-bedded, plane-bedded, rippled sandy grainstones and imbricated intraclastic conglomerates indicate storm deposition at or below fair-weather wave base. Ptygmoidal injectites and associated convolute laminae may suggest local, syndepositional seismic activity (Pratt, Reference Pratt1998; Kahle, Reference Kahle2002; Pratt and Ponce, Reference Pratt and Ponce2019). Causal seismicity is likely linked to the emplacement and uplift of the Lemhi arch and/or movement along the Snake River Transform Fault to the Northeast (Lund, Reference Lund2008; Link et al., Reference Link, Todt, Pearson and Thomas2017; Pratt, Reference Pratt2021).
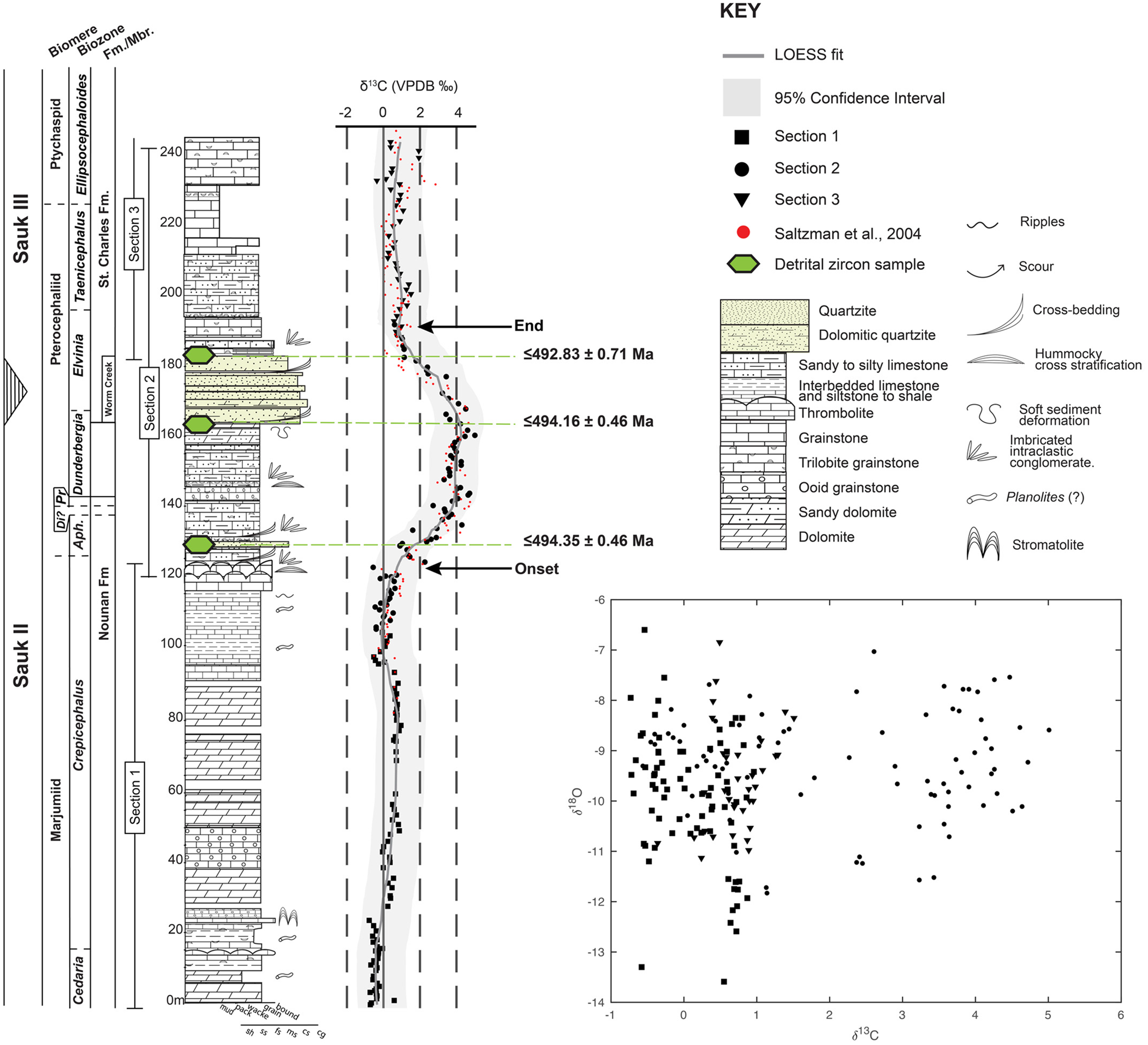
Figure 3. Composite lithostratigraphy of the Smithfield Canyon sections, estimated Sauk II–III boundary, biozones, and biomeres. Solid lines indicate well-defined biostratigraphic boundaries (e.g., first occurrence and last occurrence of biozone fauna are <1 m), and dashed lines indicate estimated biozone boundary position; δ13C stratigraphy: black points are from this study, and red points are from Saltzman et al. (Reference Saltzman, Cowan, Runkel, Runnegar, Stewart and Palmer2004); δ13C and δ18O cross plot indicating a lack of co-variation between δ13C and δ18O values and arguing that δ13C values are likely primary. Aph = Aphelaspis; Di = Dicanthopyge; Pr = Prehousia; Fm. = Formation; Mbr. = Member; VPBD = Vienna PeeDee Belemnite. Modified from Cothren et al. (Reference Cothren, Farrell, Sundberg, Dehler and Schmitz2022).
The upper contact between the Nounan Formation and the overlying Worm Creek Member is seemingly gradational with an increase in siliciclastic material up-section. The Worm Creek Member comprises tan to pale-pink variably dolomitic orthoquartzites that are fine- to medium-grained, trough-cross bedded, and plane bedded. At the top of the member, orthoquartzites are interbedded with thin cryptalgal carbonate beds, suggesting a gradational contact. The lower part of the overlying informal upper member of the St. Charles Formation is lithologically similar to the uppermost Nounan Formation at this locality, dominated by sandy trilobite grainstone.
Carbon isotopes
The δ13C stratigraphy in Smithfield Canyon covers the uppermost 160 m of the Nounan Formation, the entire 20 m Worm Creek Member, and the lower 60 m of the upper member of the St. Charles Formation, coinciding with nine trilobite biozones (Figs. 3, 4; Cothren et al., Reference Cothren, Farrell, Sundberg, Dehler and Schmitz2022; this paper). A prominent +5‰ excursion in δ13Ccarb has been identified as the SPICE (Saltzman et al., Reference Saltzman, Cowan, Runkel, Runnegar, Stewart and Palmer2004; Cothren et al., Reference Cothren, Farrell, Sundberg, Dehler and Schmitz2022). In the upper Cedaria and Crepicephalus biozones, δ13C values range from –0.5‰ to +1‰, the excursion rises to peak values of +5‰ in the Aphelaspis, Prehousia, and Dunderbergia biozones, and exhibits a plateau over 35 m (136–171 m) within the Dunderbergia and Elvinia biozones before returning to near background values within the Elvinia Biozone. Analyses of detrital zircons from three siliciclastic horizons in this section give MDAs of 494.35 ± 0.46 Ma (n = 6), 494.16 ± 0.46 Ma (n = 10), and 492.84 ± 0.73 Ma (n = 3; Cothren et al., Reference Cothren, Farrell, Sundberg, Dehler and Schmitz2022).
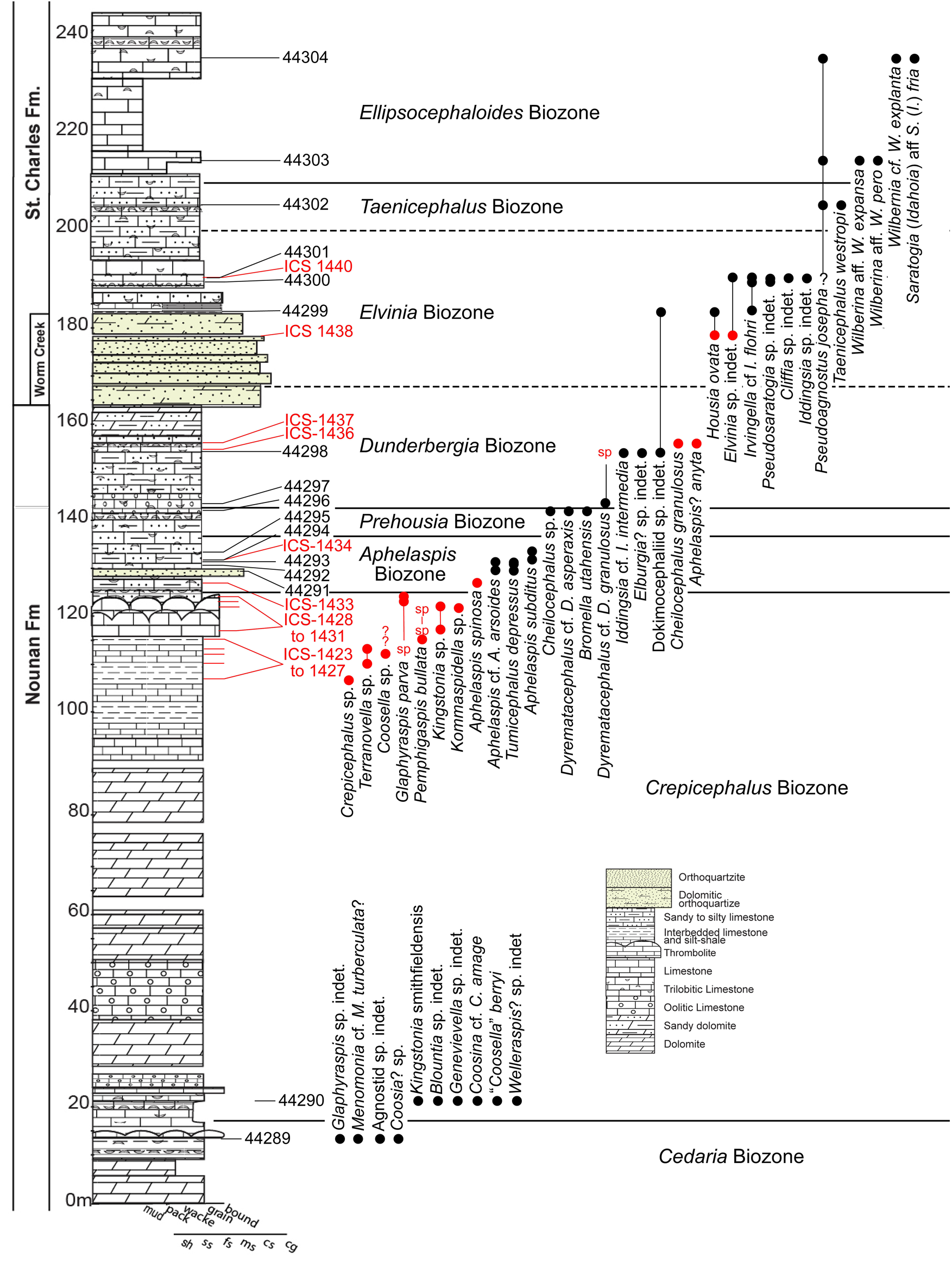
Figure 4. Composite stratigraphic column and trilobite ranges of the upper Nounan and lower St. Charles formations from Smithfield Canyon, Utah. Taxa occurrences and localities listed in red represent data from Saltzman et al. (Reference Saltzman, Cowan, Runkel, Runnegar, Stewart and Palmer2004); those listed in black represent new data from this study. sp = no specific identification by Saltzman; ? = questionable occurrence of the taxon. Section modified from Cothren et al. (Reference Cothren, Farrell, Sundberg, Dehler and Schmitz2022).
The δ13C values in the strata overlying those presented by Cothren et al. (Reference Cothren, Farrell, Sundberg, Dehler and Schmitz2022) gradually decrease toward background values within the Elvinia, Taenicephalus, and Ellipsocephaloides biozones (193–245.5 m), capturing apparent carbon cycle stabilization following the SPICE. The δ13C values range from –0.36‰ to +1.5‰, with a mean value of +0.71‰. The δ18O values have a weak correlation with δ13C values (R2 = 0.35), indicating that the δ13C values are likely primary (Fig. 3). The end of the SPICE is defined as the inflection point in which the rate of change (first derivative) of the LOESS fit of δ13C values returns to 0, indicating no change. From our analysis, the SPICE concludes at 191 m, within the Elvinia Biozone, stratigraphically above the last occurrence (LO) of Irvingella.
These new data allow a complete profile of the SPICE to be captured. The end of the SPICE within the Elvinia Biozone, stratigraphically above the LO of Irvingella, follows similar trends in the Great Basin, in which the SPICE concludes in the upper Elvinia Biozone, above the I. major subzone, with δ13C values stabilizing toward near background values in the Taenicephalus Biozone (Saltzman et al., Reference Saltzman, Runnegar and Lohmann1998). We do not observe the small +3‰ δ13C excursion near the Pterocephaliid–Ptychaspid biomere boundary recorded in Smithfield Canyon by Saltzman et al. (Reference Saltzman, Cowan, Runkel, Runnegar, Stewart and Palmer2004). While two points, at 241 m and 243 m, depart from background values at ~2‰, this is not a well-defined “excursion” as it lacks a rising or falling limb. The post-SPICE positive excursion captured by Saltzman et al. (Reference Saltzman, Cowan, Runkel, Runnegar, Stewart and Palmer2004) may not be recorded in this work due to lower sampling resolution and/or uncertainty regarding the section location and lack of data availability from Saltzman et al. (Reference Saltzman, Cowan, Runkel, Runnegar, Stewart and Palmer2004). Alternatively, rock may be missing due to a subtle unconformity or hiatal surface; however, biostratigraphy points toward a near-continuous section.
Biostratigraphy
Saltzman et al. (Reference Saltzman, Cowan, Runkel, Runnegar, Stewart and Palmer2004) presented the initial biostratigraphy of the SPICE in Smithfield Canyon on the basis of material identified by Allison Palmer (in Saltzman et al., Reference Saltzman, Cowan, Runkel, Runnegar, Stewart and Palmer2004). Unfortunately, none of the trilobite specimens that this biostratigraphy was based on have been illustrated. This paper integrates the list of taxa identified by Palmer present in the Institute of Cambrian Studies (ICS; data provided by M. Webster, personal communication, 2022) and the new collections made for this study.
Trilobite materials collected for this study occur in the Nounan and St. Charles formations and range from the Cedaria to Ellipsocephaloides biozones. These biozones represent the upper Marjuman, Steptoean, and lower Sunwaptan stages of Laurentia and the international upper Guzhangian, Paibian, and lower Jiangshanian stages of the Miaolingian and Furongian series.
The Cedaria Biozone is presently poorly documented on the basis of the occurrence of Glaphyraspis sp. indet., Coosia? sp. indet., and Menomonia cf. M. turberculata Rasetti, Reference Rasetti1965 at USNM loc. 44289. The biozone diagnostic species is Menomonia cf. M. turberculata, which is known from a single librigena with the characteristic ornamentation and shape of M. turberculata. This taxon is from the Cedaria Biozone of the Maryville Limestone of the East Coast (Rasetti, Reference Rasetti1965). By contrast, Glaphyraspis sp. indet., represented here by small cranidia, is similar to an unnamed species typically found in the lower portion of the Crepicephalus Biozone in the Great Basin (Eby, Reference Eby1981). The small cranidia identified as Coosia? sp. indet. could belong to meraspides of any related taxon to Coosia. No pygidia are associated with the cranidia.
The Crepicephalus Biozone is well documented in the section on the basis of fauna from USNM loc. 44290 that include Welleraspis? sp. indet., Genevievalla sp. indet., Coosina cf. C. amage (Walcott, Reference Walcott1916b), “Coosella” berryi (Lochman, Reference Lochman1940), Blountia sp., and Kingstonia smithfieldensis n. sp. Palmer in Saltzman et al. (Reference Saltzman, Cowan, Runkel, Runnegar, Stewart and Palmer2004) reported in ascending order Crepicephalus sp. (ICS-1423), Terranovella sp. (ICS-1424, 1426), Coosella? sp. (ICS-1425, 1426, 1428?), Glaphyraspis sp. 1 (ICS-1426), Pemphigaspis bullata Hall, Reference Hall1863, (ICS-1427), Pemphigaspis sp. (ICS-1428, 1429), Kingstonia sp. (ICS-1428, 1429), and Komaspidella? sp. (ICS-1429). Crepicephalus is diagnostic of the biozone, and Coosina amage occurs in the Crepicephalus Biozone of Tennessee (Rasetti, Reference Rasetti1965). “Coosella” berryi is known from the upper Cedaria and/or lower Crepicephalus biozones in Missouri (Lochman, Reference Lochman1940), Virginia (Rasetti, Reference Rasetti1965), Pennsylvania (Tasch, Reference Tasch1951), and Utah (Robison, Reference Robison1960; Eby, Reference Eby1981), and Genevievalla sp. indet. from the Nounan Formation is found in the upper Cedaria Biozone of the House Range (Eby, Reference Eby1981). These two taxa suggest that USNM loc. 44290 may represent the lowermost portion of the Crepicephalus Biozone.
The Aphelaspis Biozone is well documented in the section on the basis of fauna from USNM locs. 44291–44294 that include Aphelaspis subditus Palmer, Reference Palmer1962 and Aphelaspis cf. A. arsoides Rasetti, Reference Rasetti1965. Aphelaspis subditus, which is found in the upper two horizons, Palmer (Reference Palmer1965, p. 60) reported from the upper part of the Aphelaspis Biozone. Palmer (Reference Palmer1965, p. 90) stated that Tumicephalus depressus Palmer, Reference Palmer1965 is commonly found in the Dicanthopyge Biozone; however, its occurrence in the Nounan with Aphelaspis cf. A. arsoides and below A. subditus puts that in question. Palmer in Saltzman et al. (Reference Saltzman, Cowan, Runkel, Runnegar, Stewart and Palmer2004) identified Glaphyraspis parva (Walcott, Reference Walcott1899) from a 2 m interval (ICS-1430–1432), which he believed represents the lower Aphelaspis Biozone and possibly the crisis interval at the base of the Steptoean. Above these samples, Palmer reported Aphelaspis spinosa Palmer, Reference Palmer1954 (ICS-1433), Aphelaspis sp. 1 (ICS-1434), and Bromella? sp. (ICS-1434). MDAs associated within the Aphelaspis Biozone in Smithfield Canyon indicate that the Furongian Epoch, Paibian Age, and Steptoean Age must be ≤494.4 ± 0.5 Ma (Cothren et al., Reference Cothren, Farrell, Sundberg, Dehler and Schmitz2022).
The Dicanthopyge Biozone has not been documented in the section by either Saltzman et al. (Reference Saltzman, Cowan, Runkel, Runnegar, Stewart and Palmer2004) or this study. However, Tumicephalus depressus, which occurs with Aphelaspis spp. in the Nounan Formation, is typically found in the Dicanthopyge Biozone of Nevada and Utah (see Palmer, Reference Palmer1965).
The Prehousia Biozone is documented in the section on the basis of fauna from USNM locs. 44298–44296 that contains Bromella utahensis n. sp., Cheilocephalus sp. indet., and Dytremacephalus cf. D. asperaxis Palmer, Reference Palmer1965. Palmer (Reference Palmer1965, p. 85) reported Bromella veritas Palmer, Reference Palmer1965 and D. asperaxis from the Dicanthopyge and Prehousia biozones. He also reported a Dytremacephalus species similar to D. granulosus Palmer, Reference Palmer1954 from the Prehousia Biozone. The Cheilocephalus sp. indet. pygidia are similar to Cheilocephalus brachyops Palmer, Reference Palmer1965, C. brevilobus (Walcott, Reference Walcott1916b), and C. granulosus Palmer, Reference Palmer1965. These taxa occur in the Aphelaspis to Elvinia biozones according to Palmer; thus, their occurrence here in the Prehousia Biozone is reasonable. Palmer in Saltzman et al. (Reference Saltzman, Cowan, Runkel, Runnegar, Stewart and Palmer2004) did not report any taxa from the Prehousia Biozone.
The Dunderbergia Biozone is documented in the section on the basis of fauna from USNM locs. 44297 and 44298 that contain Dytremacephalus cf. D. granulosus? Palmer, Reference Palmer1954, Elburgia? sp. indet., and Iddingsia cf. I. intermedia Palmer, Reference Palmer1965. Dytremacephalus granulosus is known from the basal Dunderbergia Biozone, Elburgia spp. occurs in the middle to upper Dunderbergia Biozone, and Iddingsia intermedia Palmer, Reference Palmer1965 are found in the upper Dunderbergia Biozone according to Palmer (Reference Palmer1965). Palmer in Saltzman et al. (Reference Saltzman, Cowan, Runkel, Runnegar, Stewart and Palmer2004) identified Dytremacephalus sp. (ICS-1436), Aphelaspis? anyta (Hall and Whitfield, Reference Hall and Whitfield1877; ICS-1437 [originally reported as Dunderbergia anyta]), and Cheilocephalus granulosus Palmer, Reference Palmer1965 (ICS-1437), which he suggested may represent the lower Dunderbergia Biozone. These samples occur below the Worm Creek Quartzite in the top of the Nounan Formation.
The Elvinia Biozone is documented in the section on the basis of fauna from USNM locs. 44299 and 44301 that contain Cliffia sp. indet., Elvinia sp. indet., Housia ovata Palmer, Reference Palmer1960, Iddingsia sp. indet., Irvingella cf. I. flohri Resser, Reference Resser1942, and Pseudosaratogia sp. indet. Cliffia spp., Housia ovata, Elvinia spp., Iddingsia spp. Irvingella flohri, and Pseudosaratogia spp. are known from the Elvinia Biozone (Wilson, Reference Wilson1951; Palmer, Reference Palmer1965; Westrop, Reference Westrop1986; Pratt, Reference Pratt1992). Palmer in Saltzman et al. (Reference Saltzman, Cowan, Runkel, Runnegar, Stewart and Palmer2004) identified Elvinia? sp. (ICS-1438), Housia ovata (ICS-1438), Irvingella sp. (ICS-1439), Kindbladia sp. (ICS-1439), and Irvingella cf. I. flohri (ICS-1440), which he suggested may represent the lower to mid-Elvinia Biozone. These samples occur above the Worm Creek Quartzite in the base of the overlying upper member of the St. Charles Formation. MDA in the Elvinia Biozone, below the first occurrence of Irvingella, suggests the base of the Jiangshanian Stage is ≤492.83 ± 0.71 Ma (Cothren et al., Reference Cothren, Farrell, Sundberg, Dehler and Schmitz2022). This correlation is based on the co-occurrence of Agnostotes orientalis (Kobayashi, Reference Kobayashi1935) and Irvingella angustilimbus (Kobayashi, Reference Kobayashi1938) at the Jiangshanian Global Stratotype Section and Point (Palmer, Reference Palmer1965; Peng et al., Reference Peng, Babcock, Zuo, Zhu, Lin, Yang and Wang2012).
The Taenicephalus Biozone is documented in the section on the basis of fauna from USNM loc. 44302 that contains Taenicephalus westropi Chatterton and Gibb, Reference Chatterton and Gibb2016, Kendallina sp. indet., and Pseudagnostus josepha (Hall, Reference Hall1863). Kendallina species are known from the Taenicephalus Biozone of Alberta (Westrop, Reference Westrop1986; Chatterton and Gibb, Reference Chatterton and Gibb2016). Palmer in Saltzman et al. (Reference Saltzman, Cowan, Runkel, Runnegar, Stewart and Palmer2004) did not report any taxa from this biozone.
The Ellipsocephaloides Biozone is documented in the section on the basis of fauna from USNM locs. 44303 and 44304 that contain Saratogia (Idahoia) aff. S. (I.) fria Lochman and Hu, Reference Lochman and Hu1959, Wilbernia cf. W. explanata (Whitfield, Reference Whitfield1880), Wilbernia aff. W. expansa Frederickson, Reference Fredrickson1949, Wilbernia aff. W. pero (Walcott, Reference Walcott1890), and/or Pseudagnostus josepha. These or closely related species are known from the upper Taenicephalus to Ellipsocephaloides biozones (Frederickson, Reference Fredrickson1949; Lochman and Hu, Reference Lochman and Hu1959; Grant, Reference Grant1962; Westrop, Reference Westrop1986). Palmer in Saltzman et al. (Reference Saltzman, Cowan, Runkel, Runnegar, Stewart and Palmer2004) did not report any taxa from this biozone.
Materials and methods
Materials
All specimens were found as isolated sclerites and for the most part in poorly fossiliferous strata. As a result, the association of different sclerites is presumed to represent a single taxon if similar associations have been previously reported (e.g., Palmer, Reference Palmer1965; Westrop, Reference Westrop1986; Sundberg, Reference Sundberg1999). However, at localities with multiple taxa present, association of the different sclerites is difficult. As a result, several taxa are herein left in open nomenclature following Bergstrom (Reference Bergstrom1988), with “cf.” representing that the material is not abundant or well-enough preserved to demonstrate that it firmly belongs to the assigned taxon and “aff.” used to designate a probable new species affiliated with the species, but the samples and or preservation of the specimens are not enough to justify naming a new species.
Illustrated specimens have been coated with colloidal graphite followed by ammonium chloride sublimate. Specimen orientation for photography and measurements is primarily with the cranidial anterior border and/or palpebral lobes, librigenal border, or pygidial border in a horizontal plane. Some photographs of specimen counterparts (negative relief) have been digitally inverted using Adobe Photoshop and are labeled in the figure captions as “inverted.” This inversion included changing positive images to negatives (changing black to white and vice versa) and flipping the image horizontally.
Some materials are not preserved well enough to justify illustration, but they are mentioned so their occurrence can be noted (e.g., Agnostid sp. indet.).
Carbon isotopes
Sample preparation and carbon-isotope analyses were conducted using the methods outlined by Cothren et al. (Reference Cothren, Farrell, Sundberg, Dehler and Schmitz2022, supplemental data).
Repositories and institutional abbreviations
Specimens discussed within this paper are housed in the United States Natural History Museum (USNM). Other abbreviations include American Museum of Natural History (AMNH) and Institute of Cambrian Studies, University of Chicago (ICS).
Systematic paleontology
Sundberg is responsible for all taxonomic assignment of the new material from Smithfield Canyon.
Phylum Arthropoda von Siebold, Reference von Siebold, von Siebold and Stannius1848
Remarks
Citation of the authorship of the phylum has been variable; however, Hegna et al. (Reference Hegna, Legg, Møller, van Roy and Lerosey-Aubril2013) discussed this inconsistency in the authorship, demonstrating that Arthropoda von Siebold, Reference von Siebold, von Siebold and Stannius1848 is the correct citation.
Class Uncertain
Order Agnostida Salter, Reference Salter1864
Family Agnostidae M'Coy, Reference M'Coy1849
Agnostid sp. indet.
Remarks
A single, very small cephalon was found from the Nounan Formation (USNM loc. 44289), which cannot be identified to generic level and is left in open nomenclature.
Subfamily Pseudagnostinae Whitehouse, Reference Whitehouse1936
Genus Pseudagnostus Jaekel, Reference Jaekel1909
Type species
Agnostus cyclopyge Tullberg, Reference Tullberg1880 from the Alum Shale, Andrarum, southern Sweden (by original designation).
Pseudagnostus josepha (Hall, Reference Hall1863)
Figure 5
- Reference Hall1863
Agnostus josepha Hall, p. 178, pl. 6, figs. 54, 55.
- Reference Peng and Robison2000
Pseudagnostus josepha Peng and Robison (see for previous synonymy; also see remarks that follow).
- Reference Sundberg, Kurkewicz and Rooks2007
Pseudagnostus josepha; Sundberg et al., p. 795, fig. 3.3.
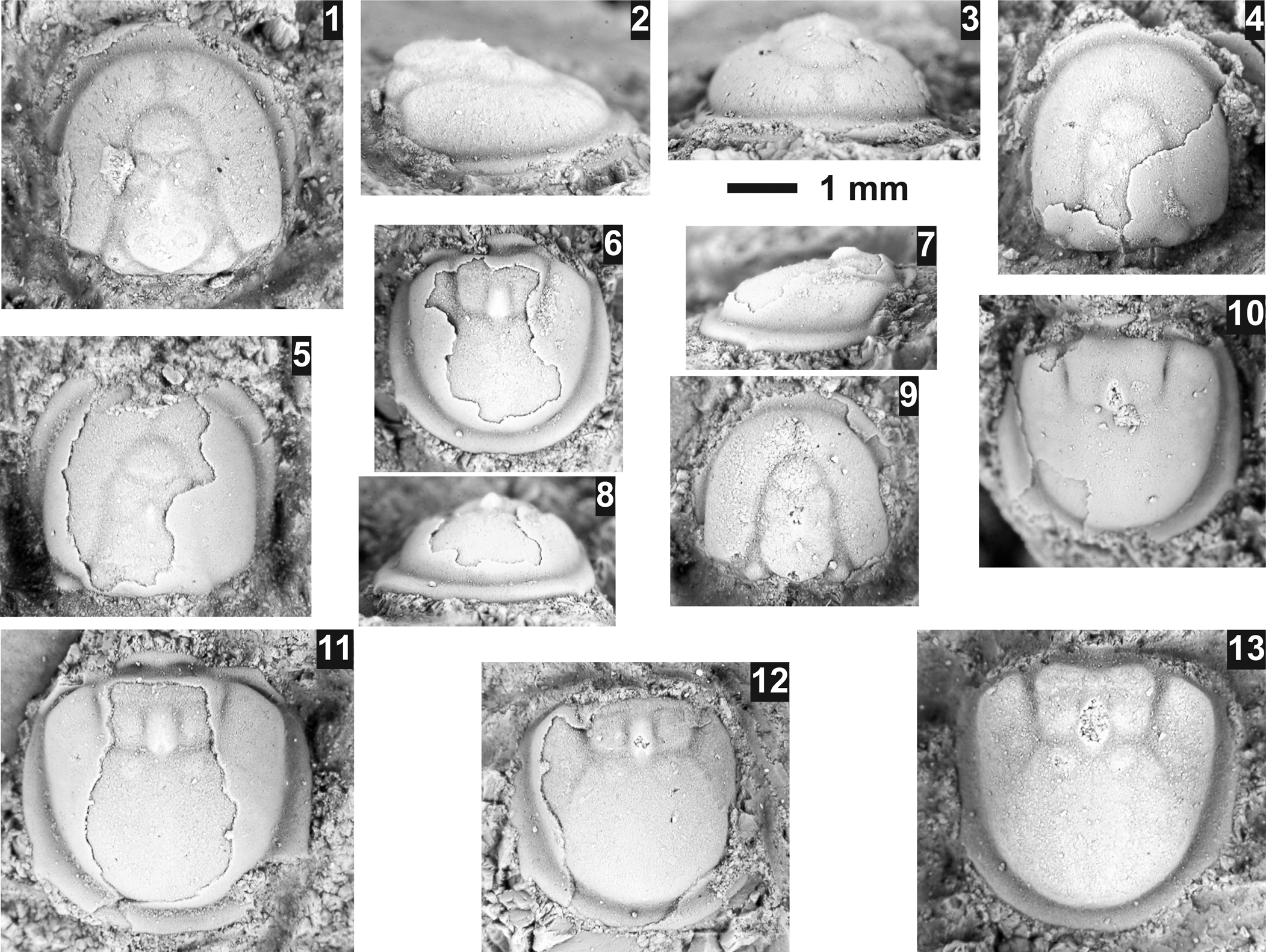
Figure 5. Pseudagnostus josepha (Hall, Reference Hall1863) from the Sunwaptan Stage, St. Charles Formation, Smithfield Canyon, Utah. All specimens are from USNM loc. 44304. (1–3) Exfoliated cephalon USNM 775702: (1) dorsal view; (2) lateral view; (3) anterior view. (4) Partially testate cephalon USNM 775706. (5) Mostly testate cephalon USNM 775705. (6–8) Mostly testate pygidium USNM 775700: (6) dorsal view; (7) lateral view; (8) anterior view. (9) Exfoliated cephalon USNM 775703. (10) Testate pygidium USNM775704. (11) Mostly testate pygidium USNM 775707. (12) Mostly exfoliated pygidium USNM 775699. (13) Exfoliated pygidium USNM 775701.
Holotype
Cotypes AMNH 311 cranidium and pygidium preserved as sandstone molds Agnostus josepha Hall, Reference Hall1863 (p. 178, pl. 6, figs. 54, 55; see Shergold, Reference Shergold1977, pl. 15, figs. 9, 10).
Occurrence
St. Charles Formation (Elvinia? to Ellipsocephaloides biozones), Smithfield Canyon, Utah (see Appendix).
Remarks
There are conflicting views as to which species of Pseudagnostus are valid or definable. Peng and Robison (Reference Peng and Robison2000) and Chatterton (Reference Chatterton2020) viewed P. josepha as a variable species that includes several previously named species. By contrast, Westrop and Eoff (Reference Westrop and Eoff2012) viewed the taxa of Pseudagnostus as more constrained in morphology and removed some taxa that Peng and Robison (Reference Peng and Robison2000) included in their synonymy. Of note is Westrop and Eoff's (Reference Westrop and Eoff2012, p. 208) removal of P. communis (Hall and Whitfield, Reference Hall and Whitfield1877) from P. josepha that was proposed by Peng and Robison (Reference Peng and Robison2000). Westrop and Eoff (Reference Westrop and Eoff2012, p. 209) pointed out that the type material of P. communis has never been photographically illustrated and is poorly known from the type area. Chatterton and Gibb (Reference Chatterton and Gibb2016; Chatterton Reference Chatterton2020) noted that P. josepha is widespread in the McKay Group, Canada, but is designated as Pseudagnostus cf. P. josepha due to its quality of preservation.
The limited samples preserved in limestone from the St. Charles Formation show a combination of features of P. josepha preserved in limestone from Alberta (Westrop, Reference Westrop1986) and China (Peng and Robison, Reference Peng and Robison2000) and Pseudagnostus cf. P. communis preserved in limestone from Newfoundland (Westrop and Eoff, Reference Westrop and Eoff2012). In terms of cephalon shape (cephalon length/width ratio), the grouping of the St. Charles Formation specimens with the China and Alberta specimens of P. josepha (Fig. 6.1) and their separation from Pseudagnostus cf. P. communis from Newfoundland suggest their placement within P. josepha. This ratio changes during ontogeny from around 97% to 106% cephalic length for P. josepha. By contrast, Pseudagnostus cf. P. communis from Newfoundland show a gradual decrease from 94% to 90% of cephalic length, suggesting Westrop and Eloff's (Reference Westrop and Eoff2012) separation of the Newfoundland specimens from the Alberta and China specimen. Glabellar widths (tr., measured at the termination of the basal lobes) of the samples from the St. Charles Formation are a bit wider than P. josepha from China and Alberta (Fig. 6.2); however, these three localities form an overall trend of glabellar width around 35% to 40% of cephalon width from 2.0 to 4.5 mm in cephalon length. By contrast, the material of Pseudagnostus cf. P. communis from Newfoundland centers around 33% of cephalon width regardless of cephalon length (1.5 to 5.0 mm), again justifying the separation of the Newfoundland specimens of P. josepha. Other cephalic differences between the St. Charles Formation specimens from the Newfoundland specimens include a larger and subcircular M3 delineated with a distinct F2 similar to the China specimens and a shallower median glabellar furrow, although this feature is variable in both the Newfoundland and China specimens. Overall, the St. Charles Formation specimens are most like P. josepha.
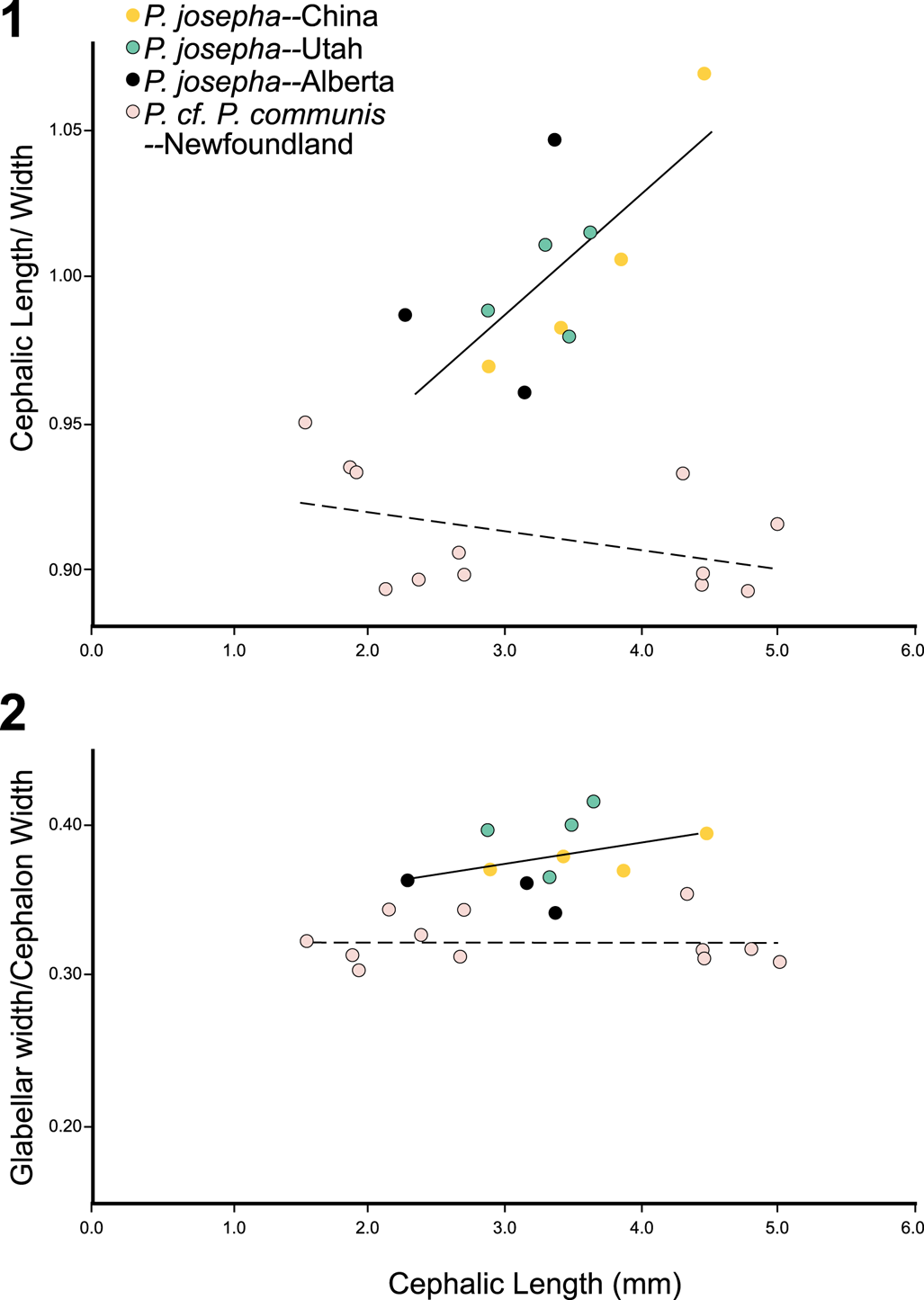
Figure 6. Bivariate plots of limestone specimens of Pseudagnostus josepha (Hall, Reference Hall1863) from China (Peng and Robison, Reference Peng and Robison2000), Alberta (Westrop, Reference Westrop1986), and Utah (this study) and Pseudagnostus cf. P. communis from Newfoundland (Westrop and Eoff, Reference Westrop and Eoff2012). (1) Cephalic length/width ratio versus cephalic length (mm), illustrating a different trend of P. cf. P. communis (dashed line) compared with specimens of P. josepha. (2) Glabellar width/cephalon width versus cephalic length (mm), again showing different trends between the two taxa.
Four partial cephala from USNM loc. 44289 are similar to P. josepha, but without associated pygidia, they cannot be firmly assigned to this species.
Class Trilobita Walch, Reference Walch1771
Order “Ptychopariida” Swinnerton, Reference Swinnerton1915
Suborder “Ptychopariina” Richter, Reference Richter1933
Remarks
See Sundberg and Webster (Reference Sundberg and Webster2022) for comments on the order and suborder of ptychopariidids.
Superfamily Olenacea Burmeister, Reference Burmeister1843
Family Aphelaspididae Palmer, Reference Palmer1960
Subfamily Aphelaspidinae Palmer, Reference Palmer1960
Remarks
Lee and Chatterton (Reference Lee and Chatterton2005) discussed the placement of this subfamily on the basis of the ontogeny of several species of Aphelaspis. They suggested that Aphelaspidinae (Housia and Aphelaspis) is closely related to Olenidae and should be placed into the superfamily Olenacea, but not in the family Pterocephaliidae. By contrast, Hopkins (Reference Hopkins2011) illustrated that Housiinae and Pterocephaliinae are sister clades when compared with the Aphelaspidinae.
Genus Aphelaspis Resser, Reference Resser1935
Type species
Aphelaspis walcotti Resser, Reference Resser1938, Nolichucky Formation, Virginia (by original designation).
Aphelaspis subditus Palmer, Reference Palmer1962
Figure 7.1–7.11
- Reference Palmer1962
Aphelaspis subditus Palmer, p. 35, pl. 4, figs. 20–22.
- Reference Lee and Chatterton2005
Aphelaspis subditus; Lee and Chatterton, p. 1360, pl. 5, figs. 1–6, 10, 21, 26 (see for further synonymy).
- Reference Cothren, Farrell, Sundberg, Dehler and Schmitz2022
Aphelaspis subditus; Cothren et al., figs. 2.9, 2.11.
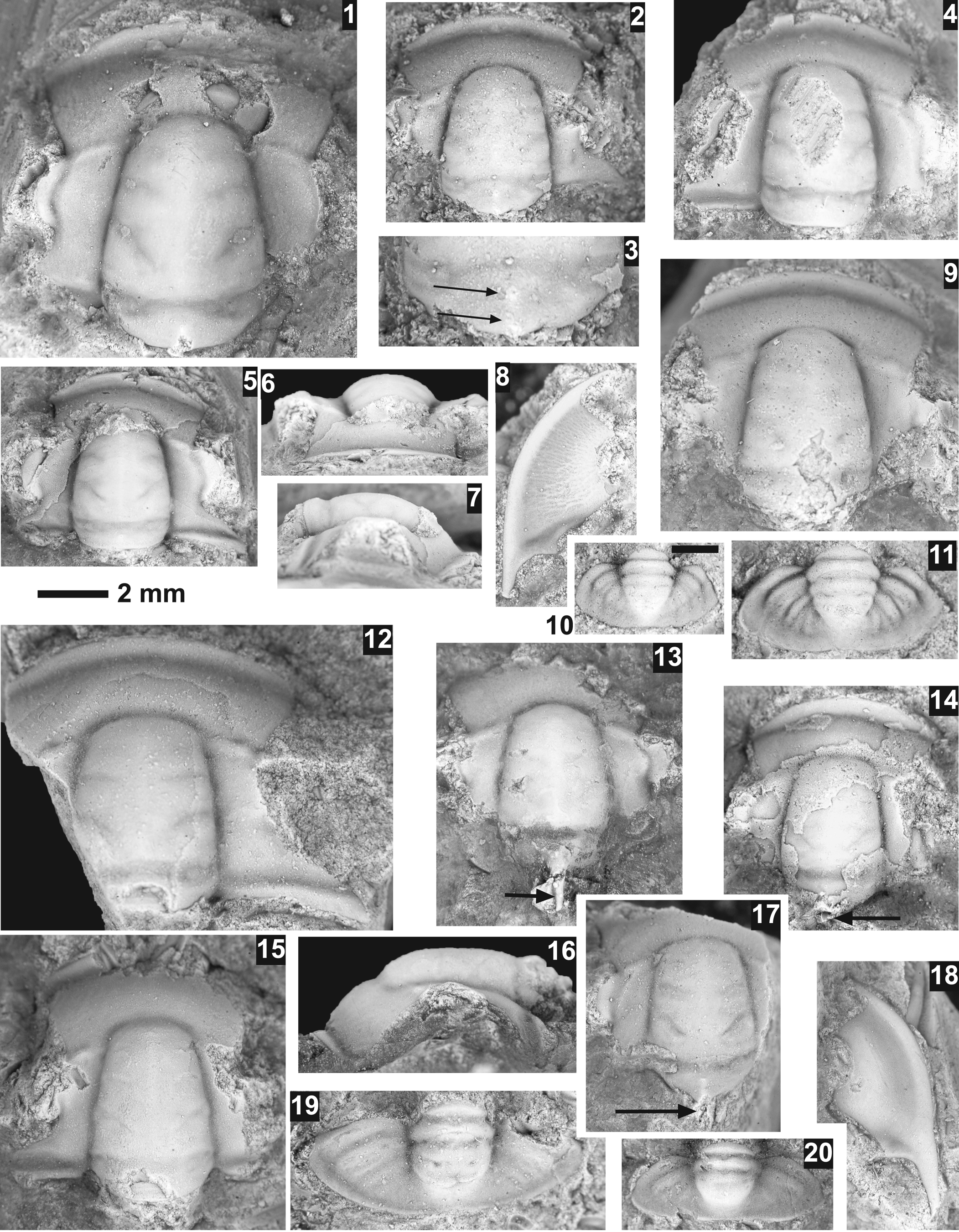
Figure 7. Aphelaspis species from the Steptoean Stage, upper Nounan Formation, Smithfield Canyon, Utah. All specimens use the 2 mm scale bar unless otherwise noted. (1–11) Aphelaspis subditus Palmer, Reference Palmer1962 from USNM loc. 44294 unless otherwise noted: (1) exfoliated cranidium USNM 775644; (2, 3) exfoliated cranidium USNM 775642, close-up of occipital ring showing two nodes (arrows); (4) exfoliated cranidium USNM 775648; (5–7) mostly exfoliated cranidium USNM 775649: (5) dorsal view; (6) anterior view; (7) lateral view; (8) testate librigena USNM 775645; (9) exfoliated cranidium USNM 775643; (10) exfoliated pygidium USNM 775646 from USNM loc. 44295, scale bar = 1 mm; (11) exfoliated pygidium USNM 775647. (12–20) Aphelaspis cf. A. arsoides Rasetti, Reference Rasetti1965 from USNM loc. 44291 unless otherwise mentioned: (12) mostly exfoliated cranidium USNM 775639 (USNM loc. 44293); (13) exfoliated cranidium USNM 775638 (USNM loc. 44293) showing long occipital spine (arrow); (14) partially testate cranidium USNM 775634 showing spine (arrow); (15, 16) exfoliated cranidium USNM 775637: (15) dorsal view; (16) lateral view; (17) exfoliated partial cranidium USNM 775635 showing spine (arrow); (18) exfoliated librigena USNM 775640 (USNM loc. 44293); (19) exfoliated pygidium USNM 775636; (20) exfoliated pygidium USNM 775641 (USNM loc. 44293).
Holotype
USNM143173a, cranidium, from the Dunderberg Formation, Cherry Creek, Nevada, USA.
Occurrence
Nounan Formation (Aphelaspis Biozone), Smithfield Canyon, Utah (see Appendix); upper Bonanza King and lower Dunderberg Shale formations (Aphelaspis Biozone), Nevada (Palmer, Reference Palmer1965); Rabbitkettle Formation (Olenaspella regularis Biozone), Mackenzie Mountains, District of Mackenzie, Canada (Pratt, Reference Pratt1992).
Remarks
The specimens from USNM 44294 are most like A. subditus in the construction of the frontal area of the cranidium and shape of the librigena. However, the pygidium is more of a rectangular outline and is more like A. brachyphaspis Palmer, Reference Palmer1962. This is not unexpected given that Palmer (Reference Palmer1965, fig. 10) suggested that this species gave rise to A. subditus. The specimens from USNM 44295 (Fig. 7.11) have a more elongated pygidium, more typical of the species. Palmer (Reference Palmer1965, p. 60) reported this species from the upper part of the Aphelaspis Biozone.
A more unusual feature of these specimens from USNM 44294 is that the occipital ring has essentially two small occipital nodes, one near the middle of the ring and the other posterior, near the posterior margin (Fig. 7.2, 7.4). This is not a feature of A. subditus or any other member of the genus from the Great Basin. However, Rasetti (Reference Rasetti1965) reported A. arses (Walcott, Reference Walcott1916a) from the Nolichucky Formation from Tennessee that has both an occipital spine and a node. Furthermore, another species with a large occipital spine similar to A. arsoides reported from Tennessee (Rasetti, Reference Rasetti1965) occurs in the Worm Creek section at USNM 44292. Aphelaspis arses, A. arsoides, and A. spinosa are the only three members of the species that have occipital spines. Specimens from USNM loc. 44295 have only one occipital node.
Aphelaspis cf. A. arsoides Rasetti, Reference Rasetti1965
Figure 7.12–7.20
- Reference Rasetti1965
Aphelaspis arsoides Rasetti, p. 94, pl. 11, figs. 15–21, pl. 12, fig. 22.
Holotype
USNM144677, cranidium, from the Nolichucky Formation, Jefferson County, Tennessee, USA.
Occurrence
Nounan Formation (Aphelaspis Biozone), Smithfield Canyon, Utah (see Appendix).
Remarks
The specimens assigned to this species are similar to the type material from the Nolichucky Formation, Tennessee, in having a long, flat-lying, and narrow occipital spine (Fig. 7.13, 7.17), horizontal intraocular region, pitting in the furrows of the cranidium (Fig. 7.14), librigenal features, and pygidial features. These specimens are dissimilar from the Tennessee species in the absence of an occipital spine in smaller specimens and less-pronounced pitting in the cranidial and librigenal anterior borders, which is the reason for the cf. designation. These specimens differ from A. arses also from Tennessee in the latter's possession of a dorsally arched, broad-based, occipital spine; pitting in the furrows of the cranidium; and more-divergent anterior branches of the facial sutures. The specimens are also very similar to the A. subditus from USNM loc. 44294, but the latter lacks the long occipital spine in specimens of the same size, pitting in the cranidial furrows, and the nearly horizontal intraocular regions. This species is also similar to A. spinosa Palmer, Reference Palmer1954 in the possession of an occipital spine but differs in having a narrower anterior border and better-defined anterior border furrow. Saltzman et al. (Reference Saltzman, Cowan, Runkel, Runnegar, Stewart and Palmer2004) reported A. spinosa from ICS-1433, which occurs approximately 1 m below the first occurrence of Aphelaspis cf. A. arsoides reported here.
Genus Bromella Palmer, Reference Palmer1965
Type species
Bromella veritas Palmer, Reference Palmer1965, Dunderberg Shale, Nevada (by original designation).
Remarks
Palmer (Reference Palmer1965) discussed the similarity of Bromella to Aphelaspis, Dytremacephalus, and Prehousia, but the differences between these genera and Bromella are consistent as outlined by Palmer (Reference Palmer1965).
Bromella utahensis new species
Figure 8
- Reference Cothren, Farrell, Sundberg, Dehler and Schmitz2022
Bromella veritas, Cothren et al., fig. 2.6, 2.10.
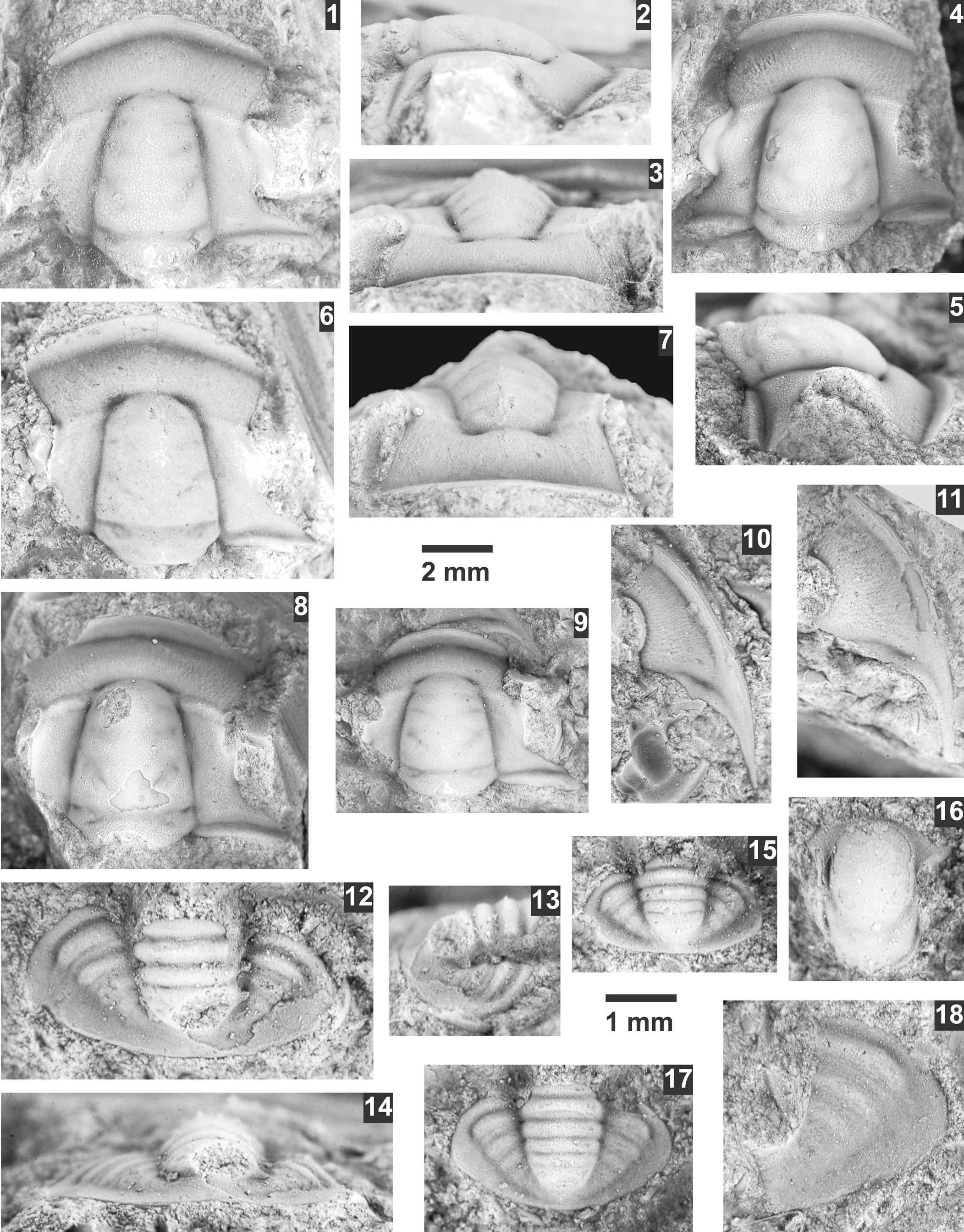
Figure 8. Bromella utahensis n. sp. from the Prehousia Biozone, Steptoean Stage, upper Nounan Formation, Smithfield Canyon, Utah. All specimens are from USNM loc. 44296 and paratypes unless otherwise mentioned. (1–3) Exfoliated cranidium USNM 775676: (1) dorsal view; (2) lateral view; (3) anterior view. (4, 5) Testate cranidium USNM 775673: (4) dorsal view; (5) lateral view. (6, 7) Exfoliated cranidium USNM 775672. (8) Testate cranidium USNM 775675. (9) Testate cranidium USNM 775674. (10) Testate librigena USNM 775677. (11) Testate librigena USNM 775679. (12–14) mostly exfoliated pygidium USNM 775681: (12) dorsal view; (13) lateral view; (14) posterior view. (15) Exfoliated pygidium USNM 775682. (16) Mostly exfoliated hypostome USNM 775680. (17) Exfoliated, holotype pygidium USNM 775678. (18) Exfoliated, partial pygidium USNM 775683. (1–11) Specimens use the 2 mm scale; (12–18) specimens use the 1 mm scale bar.
Holotype
Pygidium USMN 775678 from the Nounan Formation, Smithfield Canyon, Utah (USMN loc. 44296).
Paratypes
Specimens USMN 775669–775677 and 775679–775683 from the Nounan Formation, Smithfield Canyon, Utah (USMN loc. 44296).
Diagnosis
Pygidium with relatively long axis, four axial rings, and well-defined pleural furrows.
Occurrence
Nounan Formation (Prehousia Biozone), Smithfield Canyon, Utah (see Appendix).
Description
Cranidium of moderate size, length 6.7 ± 1.0 mm (N = 6); subpentagonal, moderate convexity (sag. and trans.); anterior margin moderately and unevenly curved, slightly arched dorsally; posterior margin (excluding occipital ring) directed laterally. Anterior branches of facial sutures divergent by 22 ± 5° from anterior border; posterior branches strongly divergent at 41 ± 6°. Glabella elongate, conical shaped, length 71 ± 2% cranidial length; width 51 ± 4% cranidial width; tapered; width at anterior end 71 ± 2% of posterior glabellar width; moderate convexity (sag. and trans.); frontal lobe rounded. Axial furrows moderately deep; preglabellar furrow forming deep pits laterally and shallow medially. Lateral glabellar furrows shallow; S1 bifurcated and deepest; S2 and S3 projected slightly posteriorly; S4 projected laterally and very faint. Occipital ring slightly elevated above rest of glabella, moderately convex; length 19 ± 1% glabellar length; posterior margin moderately convex posteriorly, with small occipital node. SO straight to very slightly curved anteriorly medially; deepest laterally; moderate depth. Frontal area length 29 ± 2% cranidial length, unequally divided. Preglabellar field slightly convex, moderately downsloping, length 66 ± 5% frontal area length. Anterior border very slightly convex, flat to upsloping, length 34 ± 5% frontal area length, slightly tapering laterally, no medial inbend or swelling. Anterior border furrow moderate depth, curvature of 146 ± 4°. Fixigena slightly convex, nearly level, anterior area strongly downsloping anteriorly, width 38 ± 4% of posterior glabellar width. Palpebral lobes moderately curved, slightly elevated, narrow, and moderately long, length 40 ± 3% glabellar length; anterior margin located transversely opposite of S3 or L4. Ocular ridge well defined, directed slightly posterolaterally from glabella at 73 ± 3° to sagittal axis. Posterior area of fixigena triangular, projected slightly posterolaterally, terminated with sharply rounded corner; length 30 ± 3% glabellar length; width 57 ± 9% glabellar length.
Librigenae moderate size, length approximately 7.5–7.8 mm (N = 2); moderately wide at about 56–59% length without spine; lateral margin moderately curved. Genal field slightly convex. Border moderately convex, slightly widening and flattening posteriorly, width about 13–20% librigenal width; border furrow moderate depth, shallowing posteriorly. Librigenal spine moderate length, about 50–64% librigenal length; broad based and flattened, directed posterolaterally.
A single hypostome possibly belonging to the species is subrectangular with narrow lateral and posterior borders and furrows, oval anterior lobe, and crescent-shaped posterior lobe approximately 25% of hypostomal length. Thin anterior border with broadening triangular wing structures laterally.
Rostral plate and thorax unknown.
Pygidium small to moderate length, length 1.5–2.0 mm (N = 3); subrectangular shaped, length 48–52% width; margin smooth with faint undulations/spines? on posterior margin, anterior margin strongly and evenly curved directed slightly posterolaterally; anterolateral corners sharply rounded, transversely opposite posterior portion of axis; no postaxial notch; moderately convex (sag.). Axis moderately tapered, mid-width 83–86% anterior width, anterior width 40–43% pygidial width; length 81–88% pygidial length, extends to nearly posterior border furrow; four axial rings, moderately convex; terminal piece moderate size, rounded posteriorly; axial ring furrows moderate depth. Pleural regions convex, downsloping around margin; pleural furrows narrow and moderately deep, shallowing posteriorly, extending to border furrow; other pleural furrows shallow. Border not slightly convex to flat, broader (tr.) laterally and narrower (sag.) medially. Border furrow well defined, shallows medially.
Fine granules occur on the cranidia, librigenae, and pygidia testate surfaces, smooth on internal surfaces. Terrace lines on the cranidial anterior border and lateral and ventral margins of the librigenal border. Genal caeca occurs on frontal area and genal area of librigena. Exoskeleton relatively thin.
Etymology
Named after the type locality in Utah.
Remarks
The pygidia from the Nounan Formation differ from the two pygidia of Bromella veritas Palmer, Reference Palmer1965 (pl. 18, figs. 5, 9) in possessing longer axes with three to four axial rings and more-pronounced pleural bands and furrows although they are roughly the same length (≈2 mm) as Palmer's illustrated specimens. These differences are the justification for assigning the Nounan specimens to a new species. Cranidial and librigenal features of both B. utahensis and B. veritas are very similar, except that the former has a granulated exoskeleton and smooth internal mold, whereas the latter has a pitted, smooth, or finely granulated surface.
Family Elviniidae Kobayashi, Reference Kobayashi1935
Subfamily Elviniinae Kobayashi, Reference Kobayashi1935
Genus Dytremacephalus Palmer Reference Palmer1954
Type species
Dytremacephalus granulosus Palmer, Reference Palmer1954, Riley Formation, Texas (by original designation).
Dytremacephalus cf. D. asperaxis Palmer, Reference Palmer1965
Figure 9.1–9.3
- Reference Palmer1965
Dytremacephalus asperaxis Palmer, p. 85, pl. 18, figs. 10–13l.
- Reference Cothren, Farrell, Sundberg, Dehler and Schmitz2022
Dytremacephalus cf. D. asperaxis, Cothren et al., fig. 2.12.
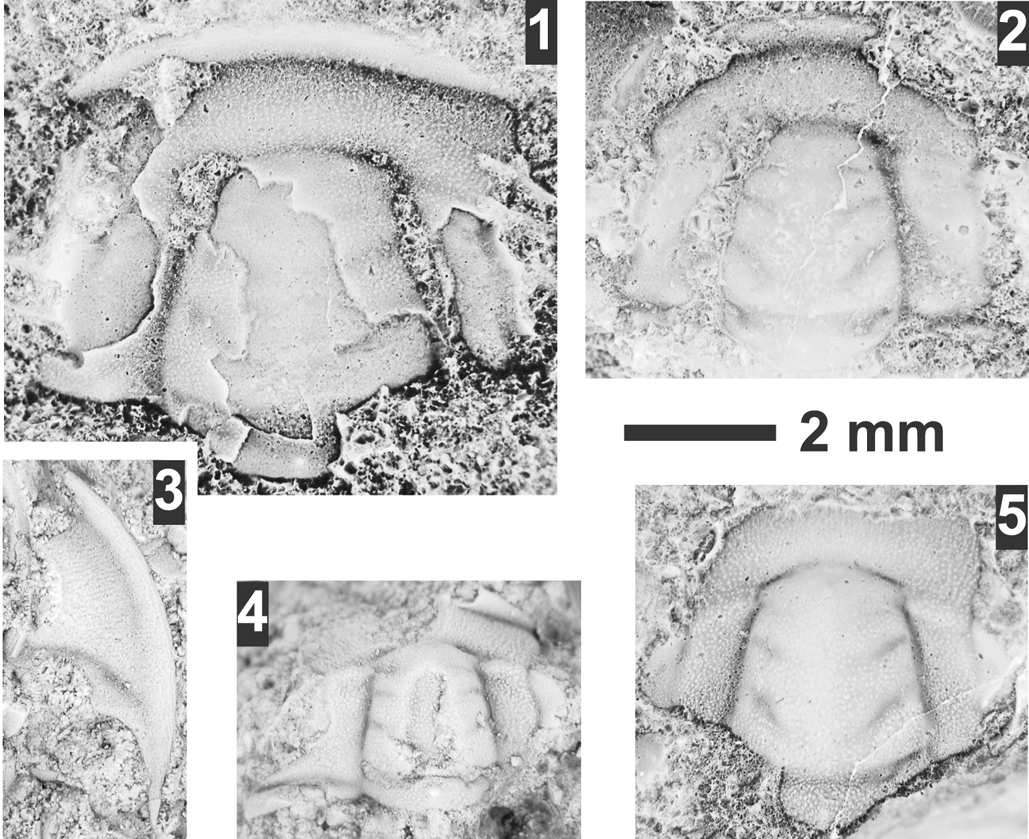
Figure 9. Dytremacephalus spp. from the Prehousia and Dunderbergia biozones, Steptoean Stage, Nounan Formation, Smithfield Canyon, Utah. (1–3) Dytremacephalus cf. D. asperaxis Palmer, Reference Palmer1965 from USNM loc. 44296: (1) partial, mostly testate cranidium USNM 775695, photograph inverted; (2) exfoliated cranidium USNM 775696, photograph inverted; (3) testate librigena USNM 775697. (4) Small cranidium USNM 775693. (5) Cranidium USNM 775694, inverted photograph.
Holotype
USNM 141761, cranidium, from the Dunderberg Formation, Bastian Peak, Nevada, USA.
Occurrence
Nounan Formation (Dunderbergia Biozone), Smithfield Canyon, Utah (see Appendix).
Remarks
These specimens are very similar to the type material of Dytremacephalus asperaxis Palmer, Reference Palmer1965 in overall cranidial and librigenal features. The “cf.” designation is the result of the palpebral lobes appearing to be slightly longer and the surface slightly more granular in these specimens, but it is difficult to ascertain given the fragmentary nature of the Smithfield Canyon specimens. This species is also similar to D. granulosus Palmer, Reference Palmer1954, but the latter differs by its coarser surface granulation.
Dytremacephalus cf. D. granulosus Palmer, Reference Palmer1954
Figure 9.4, 9.5
- Reference Palmer1954
Dytremacephalus granulosus Palmer, p. 750, pl. 85, figs. 5, 6.
- Reference Palmer1965
Dytremacephalus granulosus, Palmer, p. 85, pl. 18, figs. 14, 16–19, 21.
Holotype
USNM 123319, cranidium, from the Lion Mountain Sandstone, Riley Formation, White Creek, Texas, USA.
Occurrence
Nounan Formation (lower Dunderbergia Biozone), Smithfield Canyon, Utah (see Appendix).
Remarks
The materials from USNM loc. 44297 are fragmentary and cannot be placed in confidence. These specimens differ from D. granulosus from Texas in having a finer granulation and a more convex anterior border. The species from USNM loc. 44297 are unusual because of their granulated sclerites and relatively deeper glabellar furrows than most taxa from the Aphelaspis, Dicanthopyge, and Prehousia biozones.
Genus Elburgia Palmer, Reference Palmer1960
Type species
Crepicephalus (Loganellus) granulosus Hall and Whitfield, Reference Hall and Whitfield1877, Dunderberg Formation, Nevada (by original designation).
Elburgia? sp. indet.
Figure 10.5–10.7
Occurrence
Nounan Formation (Dunderbergia Biozone), Smithfield Canyon, Utah (see Appendix).

Figure 10. Trilobites from the Dunderbergia Biozone, Steptoean Stage, upper Nounan Formation, Smithfield Canyon, Utah. All specimens are from USNM loc. 44298. (1–4) Dokimocephaliid indet.: (1–3) partial, exfoliated cranidium USNM 775687: (1) dorsal view; (2) anterior view; (3) lateral view; (4) exfoliated librigena USNM 775688. (5–7) Elburgia? sp. indet.: (5) exfoliated cranidium USNM 775689; (6) small cranidium USNM 775690; (7) small partial cranidium USNM 775691. (1–5) Specimens use the 2 mm scale bar; (6, 7) specimens use the 1 mm scale bar.
Remarks
The material from USNM 44298 is small cranidia, mostly exfoliated and fragmented; thus, their placement within the genus or to a specific species is not possible. The material is tentatively assigned to Elburgia due to the overall glabellar shape, character of the glabellar furrows, and relatively long palpebral lobes. Ontogenetic changes may explain differences in relative glabellar shape and length, palpebral length and positions, and length of the preglabellar area observed in smaller specimens from the larger specimens illustrated by Palmer (Reference Palmer1965; for example, see in these morphologies during ontogeny illustrated by Sundberg, Reference Sundberg2020; Sundberg and Webster, Reference Sundberg and Webster2022).
Genus Elvinia Walcott, Reference Walcott1924
Type species
Dikelocephalus roemeri Shumard, Reference Shumard1861, Wilberns Formation, Texas (by original designation).
Elvinia sp. indet.
Figure 11.4
Occurrence
St. Charles Formation (Elvinia Biozone), Smithfield Canyon, Utah (see Appendix).

Figure 11. Trilobites from the Elvinia Biozone, Steptoean Stage, St. Charles Formation, Smithfield Canyon, Utah. All specimens are from USNM loc. 44301. (1–3) Pseudosaratogia sp. indet. partially exfoliated cranidium USNM 775657: (1) dorsal view; (2) lateral view; (3) anterior view. (4) Elvinia sp. indet, mostly exfoliated pygidium USNM 775661. (5–9) Iddingsia sp. indet.: (5–7) exfoliated pygidium USNM 775658: (5) lateral view; (6) dorsal view; (7) posterior vie; (8) mostly exfoliated librigena USNM 775660; (9) exfoliated librigena USNM 775659. (1–3) Specimens use the 2 mm scale bar; (4–9) specimens use the 4 mm scale bar.
Remarks
A single large, well-preserved pygidium from USMN loc. 44301 (Fig. 11.4) is very similar to the pygidium of Elvinia roemeri (Shumard, Reference Shumard1861) illustrated by Westrop (Reference Westrop1986, p. 30, fig. 15), Chatterton and Ludvigsen (Reference Chatterton and Ludvigsen1998, figs. 23, 24), Chatterton and Gibb (Reference Chatterton and Gibb2016, pl. 33–35), and Chatterton (Reference Chatterton2020, fig. 8.3, 8.4), differing in the greater angulation of the lateral anterior border. With no additional specimens or other associated exoskeleton parts, this specimen is left in open nomenclature.
Genus Irvingella Ulrich and Resser in Walcott, Reference Walcott1924
Type species
Irvingella major Ulrich and Resser in Walcott, Reference Walcott1924, Franconia Formation, Wisconsin (by original designation).
Irvingella cf. I. flohri Resser, Reference Resser1942
Figure 12.1–12.5
- Reference Resser1942
Irvingella flohri Resser, p. 24, pl. 4, figs. 12–14.
- Reference Resser1942
Irvingella adamsensis Resser, p. 24, pl. 4, figs. 7–11.
- Reference Deland and Shaw1956
Irvingella aff. I. flohri; Deland and Shaw, p. 556, pl. 66, figs. 10–12.
- Reference Palmer1965
Irvingella flohri; Palmer, p. 47, pl. 6, figs. 16, 19, 20, 24.
- Reference Westrop and Adrain2016
Irvingella flohri; Westrop and Adrain, p. 410, figs. 9, 10A–C, H–J (see for complete synonymy).
- Reference Cothren, Farrell, Sundberg, Dehler and Schmitz2022
Irvingella flohri?; Cothren et al., figs. 2.2–4.

Figure 12. Trilobites from the Elvinia Biozone, Steptoean Stage, St. Charles Formation, Smithfield Canyon, Utah. All specimens are from USNM loc. 44301. (1–5) Irvingella cf. I. flohri Resser, Reference Resser1942: (1) exfoliated partial cranidium USNM 775663; (2) pygidial axis USNM 775665; (3) mostly exfoliated cranidium USNM 775664; (4) small testate cranidium USNM 775666; (5) small testate pygidium USNM 775667. (6) Cliffia sp. indet., testate pygidium USNM 775668. (1–3) Specimens use the 2 mm scale bar; (4–6) specimens use the 1 mm scale bar.
Holotype
USNM 108667, cranidium, from the Dunderberg (?) Formation (USGS loc. 60), Eureka, Nevada, USA.
Occurrence
St. Charles Formation (Elvinia Biozone), Smithfield Canyon, Utah (see Appendix); Dunderberg Shale and unnamed limestone (middle Elvinia Biozone), Nevada and Utah (Palmer, Reference Palmer1965).
Remarks
This species is known only from the middle part of the Elvinia Biozone, but the specimens from this horizon are broken. The genal fields of the cranidia of the larger specimens are not well preserved, but they appear to be similar to I. flohri. A smaller cranidium (Fig. 12.4) and pygidium (Fig. 12.5) are also similar to the smaller cranidium and pygidium of I. flohri illustrated by Westrop and Adrain (Reference Westrop and Adrain2016, fig. 9G–K), although the cranidial anterior border and border furrow are very faint in the specimen reported here. Due to the fragmentary nature of the larger specimens, these samples are questionably placed into I. flohri.
Palmer in Saltzman et al. (Reference Saltzman, Cowan, Runkel, Runnegar, Stewart and Palmer2004) reported Irvingella cf. I. flohri from ICS-1440 in Smithfield Canyon.
Subfamily Dokimocephalinae Kobayashi, Reference Kobayashi1935
Dokimocephaliid indet.
Figure 10.1–10.4
- Reference Cothren, Farrell, Sundberg, Dehler and Schmitz2022
Apachia prima, Cothren et al., fig. 2.8.
Occurrence
St. Charles Formation (Dunderbergia Biozone), Smithfield Canyon, Utah (see Appendix).
Remarks
Three cranidia and two librigena from USNM loc. 44298 have the typical features of dokimocephaliids in a prominent, tapered to egged-shaped glabella, relatively narrow fixigena, strongly unevenly curved anterior border, and relatively long palpebral lobes. Included in this group are Apachia Frederickson, Reference Fredrickson1949, Kindbladia Frederickson, Reference Fredrickson1948, and Dunderbergia Walcott, Reference Walcott1924 (Palmer, Reference Palmer1965 assigned this genus to the subfamily Elviniinae). These specimens were originally assigned to A. prima Palmer, Reference Palmer1965 by Cothren et al. (Reference Cothren, Farrell, Sundberg, Dehler and Schmitz2022); however, they do not possess the strongly inflated glabella and strongly downsloping preglabellar area typical of the type species, A. trigonis Frederickson, Reference Fredrickson1949 (pl. 70, figs. 16, 17), or A. butlerensis (Fredrickson, Reference Fredrickson1949) and A. prima (see Palmer, Reference Palmer1965, pl. 3, figs. 5, 10, 15). The specimens are similar to Kindbladia affinis (Walcott, Reference Walcott1884) or K. wichitaensis (Resser, Reference Resser1942; see Westrop, Reference Westrop1986) but differ in their shallower lateral glabellar furrows, more-rounded anterior lobe, and longer palpebral lobes. The species Dunderbergia brevispina Palmer, Reference Palmer1965 and D. calculosa Palmer, Reference Palmer1965 and some specimens of D. variagraulata Palmer, Reference Palmer1954 (see Palmer, Reference Palmer1965, pl. 5, fig. 4) also have similar features to the cranidia and librigena from the St. Charles Formation, but the former generally differ in shallower lateral glabellar furrows, shorter palpebral lobes, and/or more elevated palpebral lobes.
Due to the limited samples of this taxon from the St. Charles Formation and the lack of pygidia, these specimens are left in open nomenclature. A possible specimen was found in collection USNM loc. 44299.
Genus Iddingsia Walcott, Reference Walcott1924
Type species
Ptychoparia similis Walcott, Reference Walcott1884, Dunderberg Formation, Nevada (by original designation).
Iddingsia sp. indet.
Figure 11.5–11.9
Occurrence
St. Charles Formation (Elvinia Biozone), Smithfield Canyon, Utah (see Appendix).
Remarks
Relatively large librigenae similar to I. similis (Walcott, Reference Walcott1884) are from USMN loc. 44301. These librigenae have a broad, convex lateral border that terminates in a long and curved genal spine (see Palmer, Reference Palmer1965, pl. 2, fig. 1). Of particular note, Westrop et al. (Reference Westrop, Poole and Adrain2010) assigned librigenae with genal spines that are narrower based and shorter to Iddingsia and related taxa.
A poorly preserved pygidium with a semicircular outline, narrow border, tapering axis with four axial rings, and weakly defined pleural and intrapleural furrows is similar to pygidia assigned to Kindbladia by Frederickson (Reference Fredrickson1948, pl., fig. 21) and Palmer (Reference Palmer1965, pl. 3, fig. 4). However, Hohensee and Stitt (Reference Hohenssee and Stitt1989) assigned a transversely elongated pygidium with a very blunt axis as belonging to the type species of Kindbladia, K. wichitaensis (Resser, Reference Resser1942). Hohensee and Stitt (Reference Hohenssee and Stitt1989, p. 870) suggested that the pygidium assigned to K. affinis (Walcott, Reference Walcott1884) by Palmer (Reference Palmer1965) should be tentatively assigned to Iddingsia robusta (Walcott, Reference Walcott1884). Due to the poor nature of the preservation, the nomenclature of this pygidium is left open. Palmer in Saltzman et al. (Reference Saltzman, Cowan, Runkel, Runnegar, Stewart and Palmer2004) also identified Kindbladia sp. from ICS1440, which is at the same level as USNM 44301 from which the poorly preserved pygidium was recovered.
Iddingsia cf. I. intermedia Palmer, Reference Palmer1965
Figure 13
- Reference Palmer1965
Iddingsia intermedia Palmer, p. 36, pl. 2, figs. 5–8.

Figure 13. Iddingsia cf. I. intermedia Palmer, Reference Palmer1965 from the Dunderbergia Biozone, Steptoean Stage, Nounan Formation, Smithfield Canyon, Utah. All specimens are from USNM loc. 44298. (1, 2) Testate cranidium USNM 775692: (1) dorsal view; (2) lateral view. (3) Mostly testate partial librigena USNM 7755574. (4) Mostly exfoliated librigena USNM 775581. (1, 2) Specimens use the 2 mm scale bar; (3, 4) specimens use the 4 mm scale bar.
Holotype
USNM 141525 cranidium, from the Dunderberg Shale, McGill, Nevada, USA.
Occurrence
Nounan Formation (Dunderbergia Biozone), Smithfield Canyon, Utah (see Appendix).
Remarks
A single, partial cranidium and two librigenae match the specimens illustrated by Palmer (Reference Palmer1965) in their cranidial convexity, granulation, glabellar shape, and glabellar furrows; however, the frontal area is mostly absent, preventing accurate identification. The two fragmentary librigenae are also similar to the specimen illustrated by Palmer (Reference Palmer1965, pl. 2, fig. 5), including the absence of obvious granules on the genal areas. The specimens are left in open nomenclature due to the fragmentary nature of the specimens. This species was found in the upper half of the Dunderbergia Biozone (Palmer, Reference Palmer1965).
Genus Pseudosaratogia Wilson, 1951
Type species
Pseudosaratogia magna Wilson, Reference Wilson1951, Gatesburg Formation, Pennsylvania (by original designation).
Pseudosaratogia sp. indet.
Figure 11.1–11.3
- Reference Cothren, Farrell, Sundberg, Dehler and Schmitz2022
Pseudosaratogia leptogranulata Cothren et al., fig. 2.1.
Occurrence
St. Charles Formation (Elvinia Biozone), Smithfield Canyon, Utah (see Appendix).
Remarks
A single well-preserved cranidium from USMN loc. 44301 and one poorly preserved cranidium from USNM loc. 44300 were recovered. These cranidia are similar to Pseudosaratogia leptogranulata Palmer, Reference Palmer1960, but the latter differs in possessing a longer and wider preglabellar area, a less-curved anterior border, a more-tapered glabella, and more-pronounced lateral glabellar furrows. The difference in glabellar tapering may be the result of ontogeny with the specimen from USMN loc. 44301 having a length of 6.7 mm versus the type specimen having a length of 11.3 mm.
Family Pterocephaliidae Kobayashi, Reference Kobayashi1935
Subfamily Housiinae Hupé, Reference Hupé1953
Genus Housia Walcott, Reference Walcott1916b
Type species
Dolichometopus (Housia) varro Walcott, Reference Walcott1916b, Orr Formation, House Range, Utah.
Housia ovata Palmer, Reference Palmer1960
Figure 14
- Reference Palmer1960
Housia ovata Palmer, p. 75, pl. 7, figs. 1–7, 9.
- Reference Palmer1965
Housia ovata; Palmer, p. 65, pl. 12, figs. 8–11.
- Reference Lee and Chatterton2005
Housia ovata; Lee and Chatterton, pl. 7, figs. 1, 2, 5, 9.
- Reference Cothren, Farrell, Sundberg, Dehler and Schmitz2022
Housia ovata; Cothren et al., fig. 2.5.

Figure 14. Housia ovata Palmer, Reference Palmer1965 from the Elvinia Biozone, Steptoean Stage, St. Charles Formation, Smithfield Canyon, Utah. All specimens are from USNM loc. 44299. (1–3) Exfoliated pygidium USNM 775684: (1) dorsal view; (2) posterior view; (3) lateral view. (4) Testate pygidium USNM 775686. (5) Exfoliated pygidium USNM 775685.
Holotype
USNM 141678 cranidium, from the Corset Springs Shale, Snake Range, Nevada, USA.
Occurrence
St. Charles Formation (Elvinia Biozone), Smithfield Canyon, Utah (see Appendix); Dunderberg Formation and Corset Springs Shale (Elvinia Biozone), Nevada (Palmer, Reference Palmer1965).
Remarks
Only pygidia were found. This species was found in the lower part of the Elvinia Biozone in several sections (Palmer, Reference Palmer1965). This species was also reported by Saltzman et al. (Reference Saltzman, Cowan, Runkel, Runnegar, Stewart and Palmer2004) from loc. ICS-1438 at Smithfield Canyon.
Genus Tumicephalus Palmer, Reference Palmer1965
Type species
Tumicephalus depressus Palmer, Reference Palmer1965 from the Dunderberg Shale, Utah, USA.
Remarks
Palmer (Reference Palmer1965) had questionably assigned Tumicephalus to the subfamily Housiinae. A phylogenetic study by Hopkins (Reference Hopkins2011) verified the genus's placement within the subfamily.
Tumicephalus depressus Palmer, Reference Palmer1965
Figure 15
- Reference Palmer1965
Tumicephalus depressus Palmer, p. 90, pl. 13, figs. 19–23.

Figure 15. Tumicephalus depressus Palmer, Reference Palmer1965 from the Prehousia Biozone, Steptoean Stage, upper Nounan Formation, Smithfield Canyon, Utah. All specimens are from 44293 unless otherwise mentioned. (1, 2) Exfoliated cranidium USNM 775653: (1) dorsal view; (2) anterior view. (3, 4) Partially testate cranidium USNM 775651: (3) dorsal view; (4) lateral view. (5) Testate librigena USNM 775656 (loc. USNM 44291). (6) Exfoliated librigena USNM 775650. (7) Exfoliated pygidium USNM 775654. (8) Mostly exfoliated pygidium USNM 775655 (loc. USNM 44291). (9) Testate pygidium and thoracic segment USNM 775652. (1–4, 6–9) Specimens use the 1 mm scale bar; (5) specimen uses the 2 mm scale bar.
Holotype
USNM 141705 cranidium from the Hicks Formation, Deep Creek Range, Utah, USA.
Occurrence
Nounan Formation (Aphelaspis Biozone), Smithfield Canyon, Utah (see Appendix); Dunderberg Formation, Lincoln Peak Formation, Hicks Formation, and Johns Wash Limestone (Dicanthopyge Biozone), Utah and Nevada (Palmer, Reference Palmer1965).
Remarks
These specimens are typically small in the collections but do not represent a juvenile of the co-occurring Aphelaspis. The specimens have an upturned anterior border with a stronger curvature, swollen preglabellar area, and dorsally sloped intraocular area from the glabella to the palpebral lobe unlike smaller specimens of Aphelaspis. Palmer (Reference Palmer1965, p. 90) stated that this species is commonly found in the Dicanthopyge Biozone; however, its occurrence with Aphelaspis cf. A. arsoides and below A. subditus either extends the range of T. depressus into the Aphelaspis Biozone or extends the range of A. suditus into the Dicanthopyge Biozone. Palmer (Reference Palmer1965) included Aphelaspis tumifrons Resser, Reference Resser1938 into Tumicephalus. Rasetti (Reference Rasetti1965) reported A. tumifrons commonly from the same horizons as A. arsoides in the southern Appalachians; at Smithfield Canyon, it occurs with Aphelaspis cf. A. arsoides.
Superfamily Uncertain
Family Catillicephalidae Raymond, Reference Raymond1938
Genus Welleraspis Kobayashi, Reference Kobayashi1935
Type species
Solenopleura jerseyensis Weller, Reference Weller1899, upper Cambrian, New Jersey (by original designation).
Welleraspis? sp. indet.
Figure 16.13
Occurrence
Nounan Formation (Crepicephalus Biozone), Smithfield Canyon, Utah (see Appendix).

Figure 16. Trilobites from the Crepicephalus Biozone, Marjuman Stage, Nounan Formation, Smithfield Canyon, Utah. All specimens are from USNM loc. 44290. (1–6) Coosina cf. C. amage (Walcott, Reference Walcott1916b): (1) partially testate cranidium USNM 775624; (2) partially testate pygidium USNM 775623; (3) partially testate cranidium USNM 775622; (4) librigena USNM 775575; (5, 6) librigena with ocular region preserved USNM 775588, inverted photograph: (5) dorsal view; (6) lateral view. (7–9) “Coosella” berryi (Lochman, Reference Lochman1940): (7, 8) exfoliated cranidium USNM 775621: (7) dorsal view; (8) anterior view; (9) exfoliated pygidium USNM 775620. (10–12) Blountia sp. indet. exfoliated pygidium USNM 775619: (10) dorsal view; (11) posterior view; (12) lateral view. (13) Welleraspis? sp. indet. cranidium USNM 775629. (14–20) Genevievalla sp. indet.: (14) testate pygidium USNM 775626; (15) partially testate pygidium USNM 775625; (16, 17) exfoliated cranidium USNM 775627: (16) dorsal view; (17) oblique view; (18, 19) partially testate cranidium USNM 775628: (18) dorsal view; (19) lateral view. (1–6) Specimens use the 4 mm scale bar; (7–19) specimens use the 2 mm scale bar.
Remarks
Only a few small cranidia were found, and without associated pygidia the genus cannot be firmly assigned.
Family Cheilocephalidae Shaw, Reference Shaw1956
Genus Cheilocephalus Berkey, Reference Berkey1898
Type species
Cheilocephalus saintcroixensis Berkey, Reference Berkey1898, St. Lawrence Formation, Minnesota (by original designation).
Cheilocephalus sp. indet.
Figure 17
- Reference Cothren, Farrell, Sundberg, Dehler and Schmitz2022
Cheilocephalus sp., Cothren et al., fig. 2.13.

Figure 17. Cheilocephalus sp. from the Prehousia Biozone, Steptoean Stage, Nounan Formation, Smithfield Canyon, Utah. All specimens are from USNM loc. 44296. (1–4) Exfoliated pygidium USNM 775698: (1) dorsal counterpart inverted photograph; (2) dorsal view; (3) posterior view; (4) lateral view.
Occurrence
Nounan Formation (Prehousia Biozone), Smithfield Canyon, Utah (see Appendix).
Remarks
The pygidium found from USNM loc. 44296 has a wide border that is slightly upturned at the edge, a high profile with a steep descent from the axis to the anterior border, and no evidence of the pleural furrows crossing the border. In these features, the pygidium is similar to Cheilocephalus brachyops Palmer, Reference Palmer1965, C. brevilobus (Walcott, Reference Walcott1916b), and C. granulosus Palmer, Reference Palmer1965. These taxa occur in the Aphelaspis to Elvinia biozones according to Palmer; thus, its occurrence here in the Prehousia Biozone is possible. Palmer in Saltzman et al. (Reference Saltzman, Cowan, Runkel, Runnegar, Stewart and Palmer2004) reported C. granulosus from loc. ICS-1437 at Smithfield Canyon. This species is known from the Dicanthopyge to Dunderbergia biozones in Nevada (Palmer, Reference Palmer1965).
Family Crepicephalidae Kobayashi, Reference Kobayashi1935
Genus Coosella Lochman, Reference Lochman1936
Type species
Coosella prolifica Lochman, Reference Lochman1936, Bonneterre Dolomite, Missouri (by original designation).
“Coosella” berryi (Lochman, Reference Lochman1940)
Figure 16.7–16.9
- Reference Lochman1940
Prochuangia? berryi Lochman (part), p. 39, pl. 4, figs. 18–20 (only).
- Reference Tasch1951
Genevievalla campbellina Tasch (part), p. 292, pl. 46, figs. 17, 18 (only).
- Reference Robison1960
Genevievalla campbellina; Robison, p. 25, pl. 2, fig. 8.
- Reference Rasetti1965
Undetermined Pygidium No. 2 Rasetti, p. 113, pl. 6, figs. 20, 21.
- Reference Cothren, Farrell, Sundberg, Dehler and Schmitz2022
Coosia sp., Cothren et al., fig. 2.14.
Holotype
USNM 98750 cranidium from the Bonneterre Dolomite, Missouri, USA.
Occurrence
Nounan Formation (Crepicephalus Biozone), Smithfield Canyon, Utah (see Appendix); Bonneterre Dolomite (Cedaria Biozone), near Jaydee, Missouri (Lochman, Reference Lochman1940); Nolinchuky Formation (Crepicephalus Biozone), Rogersville, Virginia (Rasetti, Reference Rasetti1965); Warrior Formation (Cedaria to Crepicephalus biozones?), Pennsylvania (Tasch, Reference Tasch1951); Orr Formation (upper Cedaria to lower Crepicephalus biozones), House Range, Utah (Robison, Reference Robison1960; Eby, Reference Eby1981).
Remarks
A cranidium and a few pygidia were found at USNM loc. 44290. These samples are very similar to specimens reported by Eby (Reference Eby1981) from the upper Cedaria to lower Crepicephalus biozones, House Range, Utah. Eby, in his dissertation, reported 25 cranidia and 56 pygidia of Prochuangia? berryi under a new genus, which has yet to be formally established. These sclerite associations illustrate that this species does not belong to Prochuangia as suggested by Lochman (Reference Lochman1940) or Genevievalla as reported by Tasch (Reference Tasch1951) and Robison (Reference Robison1960) on the basis of cranidial and/or pygidial differences. Rasetti (Reference Rasetti1965, p. 114) noted that the pygidia from Virginia are similar to Coosia or a related taxon, of which the authors agree. However, the cranidia are unlike any of the Coosia or related taxa in its nearly parallel-sided glabella that extends to the anterior border furrow. At present, this species is placed into “Coosella” on the basis of the pygidial characteristics, but this placement is problematic, and it probably belongs to a new genus. Specimens reported here are not complete or abundant enough to justify proposing a new generic name.
Genus Coosia Walcott, Reference Walcott1911
Type species
Coosia superba Walcott, Reference Walcott1911, Conasauga Formation, Alabama (by original designation).
Coosia? sp.
Figure 18.3, 18.4
Occurrence
Nounan Formation (Cedaria Biozone), Smithfield Canyon, Utah (see Appendix).

Figure 18. Trilobites from the Cedaria Biozone, Marjuman Stage, Nounan Formation, Smithfield Canyon, Utah. All specimens are from USNM loc. 44289. (1) Glaphyraspis sp. indet. testate cranidium USNM 775630. (2) Menomonia cf. M. tuberculata (Resser, Reference Resser1942) testate librigena USNM 775633. (3, 4) Coosia? sp. indet.: (3) exfoliated partial cranidium USNM 775632; (4) testate partial cranidium USNM 775631.
Remarks
These small cranidia could belong to meraspides of any related taxon to Coosia. No pygidia can be associated with the cranidia.
Genus Coosina Rasetti, Reference Rasetti1956
Type species
Maryvillia ariston Walcott, Reference Walcott1916b, Maryville Formation, Tennessee (by original designation).
Coosina cf. C. amage (Walcott, Reference Walcott1916b)
Figure 16.1–16.6
- Reference Walcott1916b
Blountina amage Walcott, p. 398, pl. 64, fig. 3, 3a.
- Reference Walcott1916b
Blountina alethas Walcott (part), p. 397, pl. 64, fig. 1b, c (only).
- Reference Resser1938
Coosella amage; Resser, p. 70, pl. 13, figs. 12, 13.
- Reference Rasetti1965
Coosina amage; Rasetti, Reference Rasetti1965, p. 50, pl. 7, figs. 14–22.
Holotype
USNM 62824, cranidium, from the Nolichucky Formation, near Knoxville (USNM loc. 107), Tennessee, USA.
Occurrence
Nounan Formation (Crepicephalus Biozone), Smithfield Canyon, Utah (see Appendix)
Remarks
These specimens are incompletely preserved and appear to have a shorter preglabellar area when compared to Rasetti (Reference Rasetti1965). As a result, these specimens are assigned to Coosina cf. C. amage (Walcott, Reference Walcott1916b).
Family Idahoiidae Lochman, Reference Lochman1956
Genus Saratogia Walcott, Reference Walcott1916a
Type species
Conocephalites calciferous Walcott, Reference Walcott1879 from the Hoyt Limestone, New York, (by original designation).
Subgenus Saratogia (Idahoia) Walcott, Reference Walcott1924
Type species
Idahoia serapio Walcott, Reference Walcott1924 from the Ovid Formation, Idaho, (by original designation).
Saratogia (Idahoia) aff. S. (I.) fria Lochman and Hu, Reference Lochman and Hu1959
Figure 19.1–19.10
- ?Reference Bell and Ellinwood1962
Saratogia fria Lochman and Hu, Reference Lochman and Hu1959, Bell and Ellinwood, p. 394, pl. 53, figs. 19, 20 [non figs. 13–18, 21].

Figure 19. Trilobites from the Sunwaptan Stage, St. Charles Formation, Smithfield Canyon, Utah. All specimens are from USNM loc. 44304, exfoliated or mostly exfoliated. (1–10) Saratogia (Idahoia) aff. S. (I.) fria Lochman and Hu, Reference Lochman and Hu1959: (1) small partial cranidium USNM 781838; (2) cranidium USNM 781837; (3) exfoliated cranidium USNM 781826; (4) cranidia USNM 781824 and 781825 (upper right); (5) partial cranidium USNM 781836; (6) silicified librigena USNM 781827; (7) cranidium USNM 781835; (8) cranidium USNM 781833; (9) exfoliated partial cranidium USNM 781839; (10) silicified librigena USNM 781828. (11–16) Wilbernia cf. W. explanata (Whitfield, Reference Whitfield1880): (11) partial cranidium USNM 781832; (12, 13) cranidium USNM 781829: (12) dorsal view; (13) anterior view; (14) mostly exfoliated librigena USNM 781834; (15) testate pygidium USNM 781831; (16) partial pygidium USNM 781830. (1) White scale bar = 2 mm; (2–16) specimens use the 4 mm scale bar.
Occurrence
St. Charles Formation (Taenicephalus Biozone), Smithfield Canyon, Utah (see Appendix).
Remarks
The specimens from Smithfield Canyon are similar to those of Saratogia (I.) fria from the St. Charles Formation of Idaho (Lochman and Hu, Reference Lochman and Hu1959) in glabellar shape, relative length of the preglabellar area, position of palpebral lobes, and angle of anterior suture. However, the type material (Lochman and Hu, Reference Lochman and Hu1959, pl. 59, figs. 1–11; Ludvigsen and Westrop, Reference Ludvigsen and Westrop1983, pl. 9, fig. 4) possesses a long occipital spine, which is lacking in the specimens from Smithfield Canyon, which possess only a small occipital node. Bell and Ellinwood (Reference Bell and Ellinwood1962) figured two specimens (pl. 53, figs. 19, 20) from Texas without occipital spine or node that they assign to S. fria. They mention all of the cranidial features are the same other than the spine, but one of the key features they discussed for S. fria from Texas is the presence of coarse granules on the “occipital ring, occipital spine, posterior limbs, and preoccipital glabellar lobes…” (Bell and Ellinwood, Reference Bell and Ellinwood1962, p. 394). These granules are not apparent on the type, the Texas (Ludvigsen and Westrop, Reference Ludvigsen and Westrop1983, pl. 9, fig. 5), or the Smithfield material, and the forms without an occipital spine may represent the same species as the Smithfield material.
The Smithfield material is left in open nomenclature due to the lack of well-preserved material.
Genus Wilbernia Walcott, Reference Walcott1924
Type species
Ptychoparia pero Walcott, Reference Walcott1890 from the Wilberns Formation, Texas (by original designation).
Wilbernia aff. W. expansa Frederickson, Reference Fredrickson1949
Figure 20.4–20.11
Occurrence
St. Charles Formation (Ellipsocephaloides Biozone), Smithfield Canyon, Utah (see Appendix).

Figure 20. Wilbernia species from the Sunwaptan Stage, St. Charles Formation, Smithfield Canyon, Utah. All specimens are from USNM loc. 44303. (1–3) Wilbernia aff. W. pero (Whitfield, Reference Whitfield1880), partial, exfoliated cranidium USNM 781843: (1) dorsal view; (2) lateral view; (3) anterior view. (4–11) Wilbernia aff. W. expansa Frederickson, 1949: (4–6) mostly testate cranidium USNM 781841: (4) dorsal view; (5) anterior view; (6) lateral view; (7) exfoliated librigena USNM 781840; (8–11) mostly testate pygidium USNM 781842: (8) inverted counterpart dorsal showing ornamentation; (9) dorsal view; (10) lateral view; (11) posterior view.
Remarks
The single cranidium from the St. Charles Formation is similar to Wilbernia expansa in its short frontal area, concave anterior border, wide intraocular area of the fixigena, and slightly tapered glabella. A difference between specimens illustrated by Frederickson (Reference Fredrickson1949, pl. 72, figs. 13–16) and those illustrated by Bell and Ellinwood (Reference Bell and Ellinwood1962, pl. 54, figs. 11, 12) is the shorter (sag.) anterior border in the latter. A co-occurring librigena (Fig. 20.5) also has a convex border and is also assigned to Wilbernia aff. W. expansa. Co-occurring pygidia (Figure 20.6–20.9) are similar to pygidia assigned to either Wilbernia (Bell et al., Reference Bell, Feniak and Kurtz1952, pl. 34, fig. 4, 4e; Bell and Ellinwood, Reference Bell and Ellinwood1962, pl. 54, fig. 20; Westrop, Reference Westrop1986, pl. 12, figs. 5, 10, 13) or some Levisella Ulrich in Rasetti, Reference Rasetti1944 (Ludvigsen et al., Reference Ludvigsen, Westrop and Kindle1989, pl. 6., fig. 13, pl. 7, figs. 7, 10, pl. 8, fig. 12). These specimens are left in open nomenclature due to the incomplete cranidia and limited specimens.
Wilbernia cf. W. explanata (Whitfield, Reference Whitfield1880)
Figure 19.11–19.16
- Reference Whitfield1880
Conocephalites (Ptychaspis?) explanatus Whitfield, p. 48.
- Reference Westrop1986
Wilbernia explanata, Westrop, p. 43, pl. 12, figs. 1–10, pl. 13, figs. 10–12 (see for further synonymy).
Holotype
Incomplete cranidium from the Loan Rock Formation, Hudson, Wisconsin.
Occurrence
St. Charles Formation (Ellipsocephaloides Biozone), Smithfield Canyon, Utah (see Appendix).
Remarks
The specimens from the St. Charles Formation are similar to Wilbernia explanata in their long frontal area, relatively flat anterior border, wide intraocular area of the fixigena, slightly tapered glabella, and wide pygidium. Most of the specimens reported here are fragmented, but they vary in the length of the preglabellar area, primarily on the basis of specimen size as suggested by Westrop (Reference Westrop1986, p. 43). An additional difference between specimens illustrated by Westrop (Reference Westrop1986, pl. 12, fig. 5) and herein is the rounded terminal piece of the pygidial axis versus the pointed termination of the specimens illustrated herein. The specimens are left in open nomenclature due to the incomplete cranidia and the rounded terminal piece of the pygidial axis.
Wilbernia aff. W. pero (Walcott, Reference Walcott1890)
Figure 20.1–20.3
Occurrence
St. Charles Formation (Ellipsocephaloides Biozone), Smithfield Canyon, Utah (see Appendix).
Remarks
The three cranidia from the St. Charles Formation are similar to Wilbernia pero in their short preglabellar area, narrow intraocular area of the fixigena, and nearly parallel-sided glabella. Most of the sclerites reported here are fragmented but differ from previously illustrated specimens (Bell et al., Reference Bell, Feniak and Kurtz1952, pl. 34, fig. 5a–c; Bell and Ellinwood, Reference Bell and Ellinwood1962, pl. 54, figs. 19, 21; Grant, Reference Grant1962, pl.139, fig. 8a–c; Westrop, Reference Westrop1986, pl. 13, figs. 1–3) in possessing a less convex anterior border and a less constricted (hourglass shape) glabella with a strongly rounded frontal lobe similar to those illustrated by Frederickson (Reference Fredrickson1949, pl. 72, figs. 7–9). These specimens are similar to other species of Wilbernia and specifically W. pero but are different enough that they may represent a new species. The specimens are left in open nomenclature until additional material can be found.
Family Kingstoniidae Kobayashi, Reference Kobayashi1933
Subfamily Kingstoniinae Kobayashi, Reference Kobayashi1933
Genus Kingstonia Walcott, Reference Walcott1924
Type species
Kingstonia apion Walcott, Reference Walcott1924, Maryville Formation, Tennessee (by original designation).
Kingstonia smithfieldensis new species
Figure 21
- Reference Cothren, Farrell, Sundberg, Dehler and Schmitz2022
Kingstonia sp., Cothren et al., fig. 2.16, 2.17.

Figure 21. Kingstonia smithfieldensis n. sp. from the Marjuman Stage, Nounan Formation, Smithfield Canyon, Utah. All specimens are from USNM loc. 44290, paratypes unless otherwise mentioned, testate or mostly testate, and in dorsal view unless specified. (1) Small cranidium USNM 775608. (2) Cranidium USNM 775596. (3) Cranidium USNM 775599. (4) Cranidium USNM 775603. (5) Cranidium USNM 775592. (6) Cranidium USNM 775591. (7) Cranidium USNM 775590. (8) Cranidium USNM 775584. (9) Cranidium USNM 775617. (10) Cranidium USNM 775600. (11) Cranidium USNM 775582. (12) Cranidium USNM 775614. (13) Cranidium USNM 775587. (14) Pygidium USNM 775616, right side exfoliated. (15–17) Holotype cranidium USNM 775572: (15) dorsal view; (16) anterior view; (17) lateral view. (18–20) Pygidium USNM 775577: (18) lateral view; (19) dorsal view; (20) posterior view. (21) Pygidium USNM 775618. (22) Pygidium USNM 775601. (23) Pygidium USNM 775602. (24) Pygidium USNM 775610. (25) Pygidium USNM 775576. (26) Pygidium USNM 775580.
Holotype
Cranidium USMN 775572 from the Nounan Formation, Smithfield Canyon, Utah (USMN loc.44289).
Paratypes
Specimens USMN 775573–775618 from the Nounan Formation, Smithfield Canyon, Utah (USMN loc.44289).
Diagnosis
Cranidia with tapering glabella with an expanding frontal lobe, well-defined axial furrows, and occipital ring furrow and pygidium half-moon shaped and well-defined anterior pleural furrow. Lateral glabellar furrows, axial ring furrows, and pleural furrows not defined in ventral surface.
Occurrence
Nounan Formation (Crepicephalus Biozone), Smithfield Canyon, Utah (see Appendix).
Description
Cranidium of small to moderate size, length 3.6 ± 1.2 mm (N = 32); subtriangular, moderate convexity (sag. and trans.); anterior margin moderately and evenly curved, not arched dorsally; posterior margin (excluding occipital ring) directed laterally. Anterior branches of facial sutures convergent by 38 ± 7° to anterior border; posterior branches moderately divergent at 56 ± 8°. Glabella elongate, hourglass shaped, length 100 ± 1% cranidial length; width 50 ± 2% cranidial width; slightly tapered, width at anterior end 84 ± 4% of posterior glabellar width; low convexity (sag. and trans.); frontal lobe bluntly rounded. Axial furrows moderately deep, with constriction at the L4 anterior end at 81 ± 4% of glabellar width; preglabellar furrow very shallow. Lateral glabellar furrows effaced. Occipital ring not elevated above rest of glabella, moderately convex; length 22 ± 3% glabellar length; posterior margin moderately convex posteriorly, no occipital node. SO straight to very slightly curved posteriorly medially; deepest centrally, moderate depth, very shallow laterally. Preglabellar field absent. Anterior border very slightly convex, downsloping, length 0 ± 1% cranidial length, tapering laterally, no medial inbend or swelling. Anterior border furrow very shallow. Fixigena slightly convex, nearly level, anterior area downsloping anteriorly, width 27 ± 3% of posterior glabellar width. Palpebral lobes nearly straight, level, narrow, and moderately long, length 22 ± 3% glabellar length; anterior margin located transversely opposite of S3 or L4. Ocular ridge generally not defined, directed slightly posterolaterally from glabella at 56 ± 3° to sagittal axis. Posterior area of fixigena triangular, with strong anterolateral flexure laterally, terminated with rounded corner; length 38 ± 3% glabellar length; width 38 ± 3% glabella length.
Rostral plate, hypostome, librigenal, and thorax unknown.
Pygidium small to moderate length, length 2.6 ± 0.8 mm (N = 11); half-moon shaped, length 56 ± 3% width; margin smooth, anterior margin nearly straight and directed slightly posterolaterally; anterolateral corners sharply rounded, transversely opposite anterior portion of axis; no postaxial notch; moderately convex (sag.). Axis moderately tapered, mid-width 85 ± 3% anterior width, anterior width 35 ± 2% pygidial width; length 89 ± 1% pygidial length, extends to nearly posterior margin; possibly five or six axial rings, moderately convex; terminal piece small, rounded posteriorly; axial ring furrows shallow. Pleural regions convex, downsloping around margin; anteriormost pleural furrow narrow and moderately deep laterally, extending to near margin; other pleural furrows very shallow to absent. Border not defined.
Terrace lines on the cranidial anterior border and lateral and ventral margins of the librigenal border. Other surfaces may have a small punctuate pattern, but this may be the result of preservation. Exoskeleton relatively thick.
Etymology
Named after the type locality in Smithfield Canyon.
Remarks
Many aspects of this new species are similar to Kingstonia inflata Resser, Reference Resser1938, including the cranidial outline, anterior and posterior cranidial border and furrows, exoskeleton thickness, and pygidial shape and furrows. However, unlike most representatives of Kingstonia, this species has well-defined cranidial and pygidial furrows and a relatively long (sag.) occipital ring. Also, by contrast is the apparent absence of lateral glabellar furrows and pygidial axial ring furrows and pleural/interpleural furrows on the internal molds of the sclerites (see Fig. 21.11, 21.25). Westrop (Reference Westrop1992, p. 244) discussed a potential synapomorphy linking taxa in Kingstoniidae that consists of a very short (5–10% glabellar length—calculated from specimens illustrated by Eby, Reference Eby1981) occipital ring that forms a transverse band that is depressed below the adjacent portions of the glabella. This unique narrow occipital ring occurs in Kingstonia, Bynumia Walcott, Reference Walcott1924, Ankoura Resser, Reference Resser1938, and Bynumina Resser, Reference Resser1942. Westrop (Reference Westrop1992) tentatively included Blountia (Walcott, Reference Walcott1916b) and Maryvillia Walcott, Reference Walcott1916b into Kingstoniidae, which was further substantiated by Armstrong et al. (Reference Armstrong, Westrop and Eoff2020). The new species reported here clearly does not possess this synapomorphy, with an occipital ring nearly 20% glabellar length and occurring slightly at the same elevation as the adjacent glabellar lobes and the hourglass shape of the glabella.
Subfamily Blountiinae Lochman in Lochman and Duncan, Reference Lochman and Duncan1944
Genus Blountia Walcott, Reference Walcott1916b
Type species
Blountia mimula Walcott, Reference Walcott1916b, Maryville Formation, Tennessee (by original designation).
Blountia sp. indet.
Figure 16.10–16.12
- Reference Cothren, Farrell, Sundberg, Dehler and Schmitz2022
Blountia sp. Cothren et al., fig. 2.25.
Occurrence
Nounan Formation (Crepicephalus Biozone), Smithfield Canyon, Utah (see Appendix).
Remarks
A single pygidium was found that fits this genus well, but its granular surface ornamentation is unique. Eby (Reference Eby1981) reported similar pygidia from the upper Cedaria Biozone in the House Range, Utah.
Family Llanoaspididae Lochman in Lochman and Duncan, Reference Lochman and Duncan1944
Genus Genevievella Lochman, Reference Lochman1936
Type species
Genevievella neunia Lochman, Reference Lochman1936, Bonneterre Dolomite, Missouri (by original designation).
Genevievella sp. indet.
Figure 16.14–16.19
Occurrence
Nounan Formation (Crepicephalus Biozone), Smithfield Canyon, Utah (see Appendix).
Remarks
A few cranidia and pygidia were found at USNM loc. 44290. These samples are unlike the previously named species but are very similar to specimens reported by Eby (Reference Eby1981) from the upper Cedaria Biozone, House Range, Utah.
Family Loganellidae Rasetti in Moore, Reference Moore1959
Genus Noelaspis Ludvigsen, Westrop, and Kindle, Reference Ludvigsen, Westrop and Kindle1989
Type species
Noelaspis bilobata Ludvigsen, Westrop, and Kindle, Reference Ludvigsen, Westrop and Kindle1989 from the Shallow Bay Formation, western Newfoundland (by original designation).
Noelaspis? sp. indet.
Figure 22
Occurrence
St. Charles Formation (Taenicephalus Biozone), Smithfield Canyon, Utah (see Appendix)

Figure 22. Noelaspis? sp. from the Sunwaptan Stage, St. Charles Formation, Smithfield Canyon, Utah. All specimens are from USNM loc. 44302. (1) Exfoliated cranidium USNM 775709. (2–4) Exfoliated cranidium USNM 775708: (2) dorsal view; (3) anterior view; (4) oblique view. (5) Exfoliated partial pygidium USNM 775712. (6) Exfoliated hypostome USNM 775710. (7) Inverted photograph of partial pygidium USNM 775711. (1–4, 6) Specimens use the 4 mm scale bar; (5, 7) specimens use the 2 mm scale bar.
Remarks
These specimens are most similar to Noelaspis in the placement and lengths of the palpebral lobes, glabellar furrows, lateral glabellar furrows, fixigenal width, frontal area, and pygidial outline. However, the specimens are questionably assigned to Noelaspis on the basis of the stronger rounded frontal lobe, narrower glabella, and strongly upsloping intraocular region of the fixigena. The associated hypostome is similar to the closely related Orygmaspis Resser, Reference Resser1937 as illustrated by Ludvigsen et al. (Reference Ludvigsen, Westrop and Kindle1989, pl. 10, fig. 5).
Noelaspis has previously been reported from the Beothuckia duomenta fauna of Newfoundland, which is compared to the Stigmacephalus oweni fauna and Ellipsocephaloides Biozone of Alberta (Westrop, Reference Westrop1986, Ludvigsen et al., Reference Ludvigsen, Westrop and Kindle1989).
Type species
Liostracus parvus Walcott, Reference Walcott1899, upper Cambrian, Wyoming (by original designation).
Glaphyraspis sp. indet.
Figure 18.1
Occurrence
Nounan Formation (Cedaria Biozone), Smithfield Canyon, Utah (see Appendix).
Remarks
Two small cranidia have a parallel-sided glabella, wide fixigena, and a very thin anterior border similar to a form found by Eby (Reference Eby1981) from the lower Crepicephalus Biozone of the House Range, Utah. These specimens may represent a new species, but the lack of material requires open nomenclature.
Family Menomoniidae Walcott, Reference Walcott1916a
Genus Menomonia Walcott, Reference Walcott1916a
Type species
Conocephalites calymenoides Whitfield, Reference Whitfield1878, Eau Claire Formation, Wisconsin (by original designation).
Menomonia cf. M. tuberculata Rasetti, Reference Rasetti1965
Figure 18.2
- Reference Rasetti1965
Menomonia tuberculata Rasetti, p. 62, p. 2, figs. 19–24.
Holotype
Cranidium USMN 144696 from the Maryville Limestone, Hawkins County, near Rogersville, Tennessee, USA.
Occurrence
Nounan Formation (Cedaria Biozone), Smithfield Canyon, Utah (see Appendix).
Remarks
A single librigena with the characteristic surface sculpture and shape of M. tuberculata Rasetti, Reference Rasetti1965 was found. The Nounan specimen differs in have a larger range in tubercle sizes than the single librigena illustrated by Rasetti (Reference Rasetti1965, p. 2, fig. 22). This taxon is from the Cedaria Biozone of the Maryville Limestone of Tennessee (Rasetti, Reference Rasetti1965).
Family Phylacteridae Ludvigsen and Westrop in Ludvigsen, Westrop, and Kindle, Reference Ludvigsen, Westrop and Kindle1989
Genus Cliffia Wilson, Reference Wilson1951
Type species
Acrocephalites latagenae Wilson, Reference Wilson1949, Morgan Creek Member, Wilberns Formation, Texas (by original designation).
Cliffia sp. indet.
Figure 12.6
Occurrence
St Charles Formation (Elvinia Biozone), Smithfield Canyon, Utah (see Appendix).
Remarks
A single triangular-shaped pygidium with prominent intrapleural furrows, broad axial furrows with faint nodes, and long axis resembles either Cliffia (see Wilson, Reference Wilson1951, pl. 90, fig. 22; Westrop, Reference Westrop1986, pl. 27, figs. 4–6) or Aphelotoxon Palmer, Reference Palmer1965 (pl. 19, figs. 6, 11). Given the prominent pleural bands and elongated axis, the specimen is assigned to Cliffia; however, the broadness of the axis is more typical of Aphelotoxon.
Family Parabolinoididae Lochman, Reference Lochman1956
Genus Taenicephalus Ulrich and Resser in Walcott, Reference Walcott1924
Type species
Conocephalites shumardi Hall, Reference Hall1863 from the Lone Rock Formation, Wisconsin (by original designation).
Taenicephalus westropi Chatterton and Gibb, Reference Chatterton and Gibb2016
Figure 23.1–23.10, 23.12–23.14
- Reference Westrop1986
Taenicephalus sp. A, Westrop, p. 51, pl. 22, figs. 14–16.
- Reference Chatterton and Gibb2016
Taenicephalus westropi Chatterton and Gibb, p. 81, pl. 57, figs. 1–5, 7.

Figure 23. Trilobites from the Taenicephalus Biozone, Sunwaptan Stage, St. Charles Formation, Smithfield Canyon, Utah. All specimens are from USNM loc. 44302, in dorsal view. (1–10, 12–14) Taenicephalus westropi Chatterton and Gibb, Reference Chatterton and Gibb2016: (1) mostly exfoliated cranidium USNM 781844; (2, 3) part, counterpart of exfoliated cranidium USNM 781848, inverted photograph of internal mold; (4) exfoliated cranidium USNM 781847; (5) exfoliated cranidium USNM 781845; (6, 7) part, counterpart of exfoliated cranidium USNM 781852; (8) testate librigena USNM 781850; (9) exfoliated librigena USNM 781849; (10) mostly exfoliated cranidium USNM 781846; (12) exfoliated pygidium USNM 781854; (13) exfoliated pygidium USNM 781853; (14) exfoliated pygidium USNM 781855. (11) Kendallina sp. indet. exfoliated cranidium USNM 781851. (1–10) Specimens use the 2 mm scale bar; (11–14) specimens use the 1 mm scale bar.
Holotype
Articulated carapace (UA14129) from the McKay Group, high in Green Creek Section, British Columbia, Canada.
Occurrence
St. Charles Formation (Taenicephalus Biozone), Smithfield Canyon, Utah (see Appendix); Bison Creek Formation (Taenicephalus Biozone), Alberta (Westrop, Reference Westrop1986); McKay Group, British Columbia (Chatterton and Gibb, Reference Chatterton and Gibb2016).
Remarks
Specimens from the St. Charles Formation are similar to Taenicephalus sp. A of Westrop (Reference Westrop1986) in their smooth surfaces and subequally divided frontal area. Chatterton and Gibb (Reference Chatterton and Gibb2016) assigned Westrop's specimens to T. westropi on the basis of complete specimens preserved in shale. The St. Charles specimens are also similar to T. shumardi (Hall, Reference Hall1863) illustrated by Westrop (Reference Westrop1986, pl. 21, figs. 5–17) in their convex preglabellar area, anterior border length, position of the palpebral lobes, and nature of the pygidium, but the latter has a more textured surface and an unequal division of the frontal area.
Genus Kendallina Berg in Moore, Reference Moore1959
Type species
Conocephalites eryon Hall, Reference Hall1863, from the Loan Rock Formation, Wisconsin (by original designation).
Kendallina sp. indet.
Figure 23.11
Occurrence
St. Charles Formation (Taenicephalus Biozone), Smithfield Canyon, Utah (see Appendix).
Remarks
A single small cranidium is here reported from the St. Charles Formation. The specimen is similar to Kendallina eryon illustrated by Westrop (Reference Westrop1986, pl. 16, figs. 11–13) in cranidial shape but has deeper lateral glabellar furrows. The specimen is also similar to Kendallina crassitesta Westrop, Reference Westrop1986 (pl. 20, figs.1–6, pl. 21, figs. 3, 4; Chatterton and Gibb, Reference Chatterton and Gibb2016, pl. 55, figs. 1–10) but differs in the presence of a nonpitted anterior border furrow, more tapered glabella, and deeper glabellar furrows. These features could be the result of ontogeny given the small size of the specimen reported herein; thus, the specimen is left in open nomenclature.
Acknowledgments
We are thankful to all who assisted during fieldwork. We thank the staff at the Utah State University Stable Isotope Geochemistry Lab. We appreciate assistance from J. Wilkins at the United States Forest Service in obtaining permits to collect rocks in the Naomi Peak Wilderness Area. Helpful and constructive reviews and additional communications were provided by S. Westrop and B. Pratt.
This work was generously funded by the National Science Foundation EAR# 1337887 and the Denver Museum of Natural History and Science. Partial support for this work came from NSF grant EAR-1955155 from the Sedimentary Geology and Paleobiology Program.
Declaration of competing interests
The authors declare none.
Appendix
Localities
Samples are from three measured sections approximately 1.5 miles from the trailhead of Smithfield Canyon, 5 miles east of Smithfield, Utah. Correlations of sections one and two use the base of the ~9 m thick thrombolite horizon as a datum. Correlations of sections two and three use the top of the Worm Creek Member of the St. Charles Formation as a datum. Sample horizons are reported as total meterage from the base of the composite section with meterage from the base of individual sections in parentheses.
Smithfield Canyon—Section 1
Measured by C.M.D. and described by H.R.C. on the southwest-facing slope starting at approximately 7,800 ft elevation and 1,110 ft above the trail. This section is on the ridge between the two north–south drainages north of the “Grotto Ridge.” A total of 115 m was measured starting at the first exposure above a vegetated drainage; 0–21 m was measured on 28 June 2021 and 21–115 m was measured on 9 September 2021. Samples collected by C.M.D. and H.R.C. on 28 June 2021.
Bottom of section at 41.90311°, –111.70677°; top at 41.90313°, –111.70665°.
USNM-44289: Thinly bedded to wavy bedded trilobite packstone to grainstone interbedded with siltstone to shale at 11.0 m (11.0 m). Cedaria Biozone. Glaphyraspis sp. indet., Coosia? sp. indet., Menomonia cf. M. tuberculata, agnostid sp.
USNM-44290: Thinly bedded to wavy bedded trilobite packstone to grainstone interbedded with siltstone to shale at 18.5 m (18.5 m). Crepicephalus Biozone. Kingstonia smithfieldensis n. sp., Blountia sp. indet., Genevievalla sp. indet., “Coosella” berryi, Coosina cf. C. amage, Welleraspis? sp. indet.
Smithfield Canyon—Section 2
Measured by C.M.D. and described by H.R.C. on the west-facing slope starting at approximately 7,200 ft elevation and 800 ft above the trail. This section is on the ridge east of the second (west to east) drainage north of the “Grotto Ridge.” A total of 120 m was measured starting at the top of a dolomite cliff (correlative to ~50–75 m in section 1); 0–33 m was measured 15 November 2021, 33–90 m was measured 2 July 2021, and 90–120 m was measured 7 August 2021.
Bottom of section at 41.90042°, –111.70185°; top at 41.90042°, –111.70185°.
USNM-44291: Sandy, trough cross-bedded, trilobite grainstone at 129.5 m (56 m). Aphelaspis Biozone. Collected by C.M.D. and H.R.C. 2 July 2021. Tumicephalus depressus, Aphelaspis cf. A. arsoides.
USNM-44292: Sandy, trough cross-bedded, trilobite grainstone at 130.6 m (57 m). Aphelaspis Biozone. Collected by C.M.D. and H.R.C. 2 July 2021. Tumicephalus depressus, Aphelaspis cf. A. arsoides.
USNM-44293: Sandy, trough cross-bedded, trilobite grainstone at 131.2 m. Aphelaspis Biozone. Collected by C.M.D. and H.R.C. 2 July 2021. Tumicephalus depressus, Aphelaspis cf. A. arsoides.
USNM-44294: Sandy, trough cross-bedded, trilobite grainstone at 131.7 m (58 m). Aphelaspis Biozone. Collected by C.M.D. and H.R.C. 28 June 2021. Aphelaspis subditus.
USNM-44295: Sandy, trough cross-bedded, trilobite grainstone at 132.4 m (59 m). Aphelaspis Biozone. Collected by C.M.D. and H.R.C. 28 June 2021. Aphelaspis subditus.
USNM-44296: Rippled trilobite grainstone with interbedded (5 cm thick) sand stringers at 141 m (67.5 m). Prehousia Biozone. Collected by C.M.D. and H.R.C. 8 August 2021. Bromella utahensis, Dytremacephalus cf. D. asperaxis, Cheilocephalus sp. indet.
USNM-44297: Normally graded oolite packstone interbedded with trilobite grainstone at 142 m (68.5 m). Dunderbergia Biozone. Collected by C.M.D. and H.R.C. 28 June 2021. Dytremacephalus cf. D. granulosus.
USNM-44298: Sandy trilobite grainstone at 151.8 m (78 m). Dunderbergia Biozone. Collected by C.M.D. and H.R.C. 7 August 2021. Elburgia? sp. indet., Iddingsia cf. I. intermedia, Dokimocephaliid sp. indet.
USNM-44299: Trilobite grainstone interbedded with shale at 183.2 m (110 m). Elvinia Biozone. Collected by C.M.D. and H.R.C. 7 August 2021. Housia ovata, Irvingella sp., Dokimocephaliid sp. indet.
USNM-44300: Trilobite grainstone at 189.5 m (116 m). Elvinia Biozone. Collected by C.M.D. and H.R.C. 7 August 2021. Pseudosaratogia sp. indet.
USNM-44301: Trilobite grainstone with intraclastic conglomerate at 190.5 m (117 m). Elvinia Biozone. Collected by C.M.D. and H.R.C. 7 August 2021. Cliffia sp. indet., Elvinia sp. indet., Irvingella cf. I. flohri, Iddingsia sp. indet., Pseudosaratogia sp. indet., Pseudagnostus josepha?.
Smithfield Canyon—Section 3
Measured by C.M.D. and described by H.R.C. on the “Grotto Ridge” starting at 7,000 ft elevation and 400 ft above the trail. A total of 180 m was measured on 17 June 2022 beginning at the top the Worm Creek Member of the St. Charles Formation. Samples collected by C.M.D. and H.R.C. 17 June 2022.
Bottom of section at 41.89525°, –111.70109°; top at 41.89247°, –111.69478°.
USNM-44302: Medium-bedded trilobite grainstone with sandy stringers at 183.2 m (25.7 m). Taenicephalus Biozone. Noelaspis? sp. indet., Taenicephalus westropi, Kendallina sp. indet., Pseudagnostus josepha.
USNM-44303: Thinly bedded to wavy-bedded trilobite grainstone at 189.5 m (34.5 m). Ellipsocephaloides Biozone. Wilbernia aff. W. pero, Wilbernia aff. W. expansa, Pseudagnostus josepha.
USNM-44304: Trilobite packstone with trilobite lags at 190.5 m (55 m). Ellipsocephaloides Biozone. Saratogia (Idahoia) aff. S. (I.) fria, Wilbernia cf. W. explanata, Pseudagnostus josepha.
























 Open access
Open access