


During the course of floristic studies of Japanese lichens, Glyphis cicatricosa Ach. (Graphidaceae, Ascomycota), a pantropical species (Galloway Reference Galloway2007), was found on the bark of Phellodendron amurense Rupr. in a suburban area of Tsukuba-city (36°06'03''N, 140°06'43''E). This is on the Pacific side of eastern Honshu, with a warm-temperate climate and is the northernmost locality of G. cicatricosa in Japan. The sapling tree was planted in the Tsukuba Botanical Garden (c. 14 ha, 20 m elev.) in 1984 when the mean annual temperature was 12°C, but this has gradually increased since then to almost 15°C in 2015 when G. cicatricosa was discovered (Fig. 1). Does an increase in annual temperature at this site plausibly explain the unexpected phytogeographical distribution of this lichen in terms of ‘global warming’, or is it the result of an ‘urban heat island’ effect, or indeed is it related to one or more other factors?
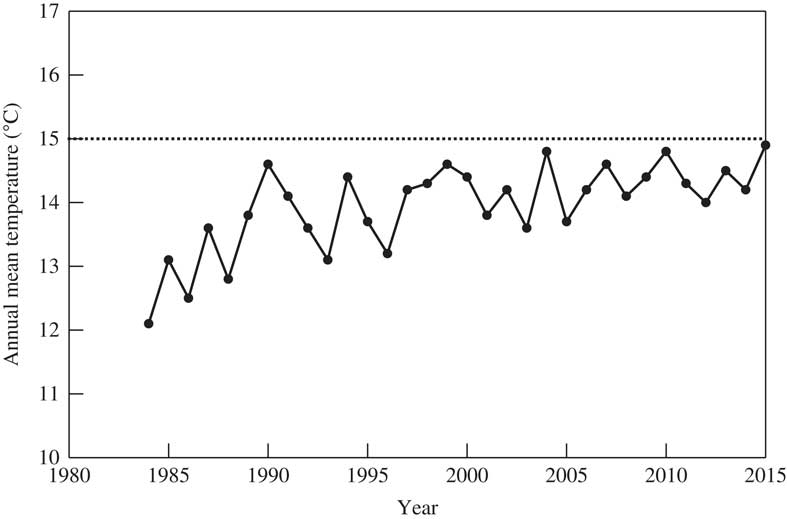
Fig. 1 Mean annual temperature in Tsukuba-city, 1984 to 2015, at the monitoring station located 5·5 km S of the Botanical Garden (Japan Meteorological Agency; http://www.jma.go.jp/). Dashed line represents the threshold 15°C isotherm, the ‘Crinum Line’, which correlates approximately with the distribution of Glyphis cicatricosa.
A voucher specimen of G. cicatricosa collected during a field survey in 2015, now housed in the National Museum of Nature and Science (TNS), was compared with other specimens of the species in TNS listed below. Morphological observations were made using a dissecting microscope (Olympus SZX16) and a differential interference contrast microscope (Olympus BX51). Anatomical examination was undertaken using hand-cut sections mounted in GAW (glycerine: ethanol: water, 1:1:1).
Meteorological and air pollution data were obtained from the Japan Meteorological Agency and the National Institute for Environmental Studies respectively; it should be noted that the distances between the Botanical Garden and the meteorological and air pollution monitoring stations are 5·5 and 8·1 km, respectively.
Glyphis cicatricosa is characterized by its greyish white or brownish thallus with crowded lirellae formed in rounded to elliptical pseudostromata; lirellae with open discs that are covered with brownish pruina; clear hymenium; 8-spored asci with hyaline spores which are 6–12-locular with lenticular cells, 30–55×7–12 μm; and absence of specific chemical substances (Nakanishi Reference Nakanishi1966; Archer Reference Archer2009). The specimen collected from Tsukuba-city agrees with the morphological and chemical features mentioned above, with spore sizes ((min.–) mean±1SD (–max.)) as follows: (31·7–)35·1±3·8(–45·1)×(6·7–) 7·8±0·7(–9·2) μm (n=10).
Glyphis cicatricosa is widely distributed in tropical to warm-temperate regions such as south-eastern USA, Central and South America, South, South-East and East Asia, Australia, New Zealand and the Pacific (e.g. Archer Reference Archer2009; Moon et al. Reference Moon, Nakanishi, Futagami and Kashiwadani2015). In Japan, it is reported from warm-temperate and subtropical regions in the south and the Bonin Islands (Nakanishi Reference Nakanishi1966; Kurokawa Reference Kurokawa1969). The record from Tsukuba-city is the most northern locality within Japan (Fig. 2). Here the tree on which it occurs is isolated in a relatively open setting (Fig. 3). The tree on which G. cicatricosa is found was originally transplanted from a montane area of central Japan (Yamanashi Prefecture) where this lichen is absent and has almost certainly become established after the tree was collected. On the tree trunk c. 10 thalli (each 1–3 cm in diam.) grow, 30 to 150 cm above the ground, mainly in association with Graphis handelii Zahlbr. and scattered thalli of Lecanora pulverulenta Müll. Arg. and Pertusaria pertusa (L.) Tuck.
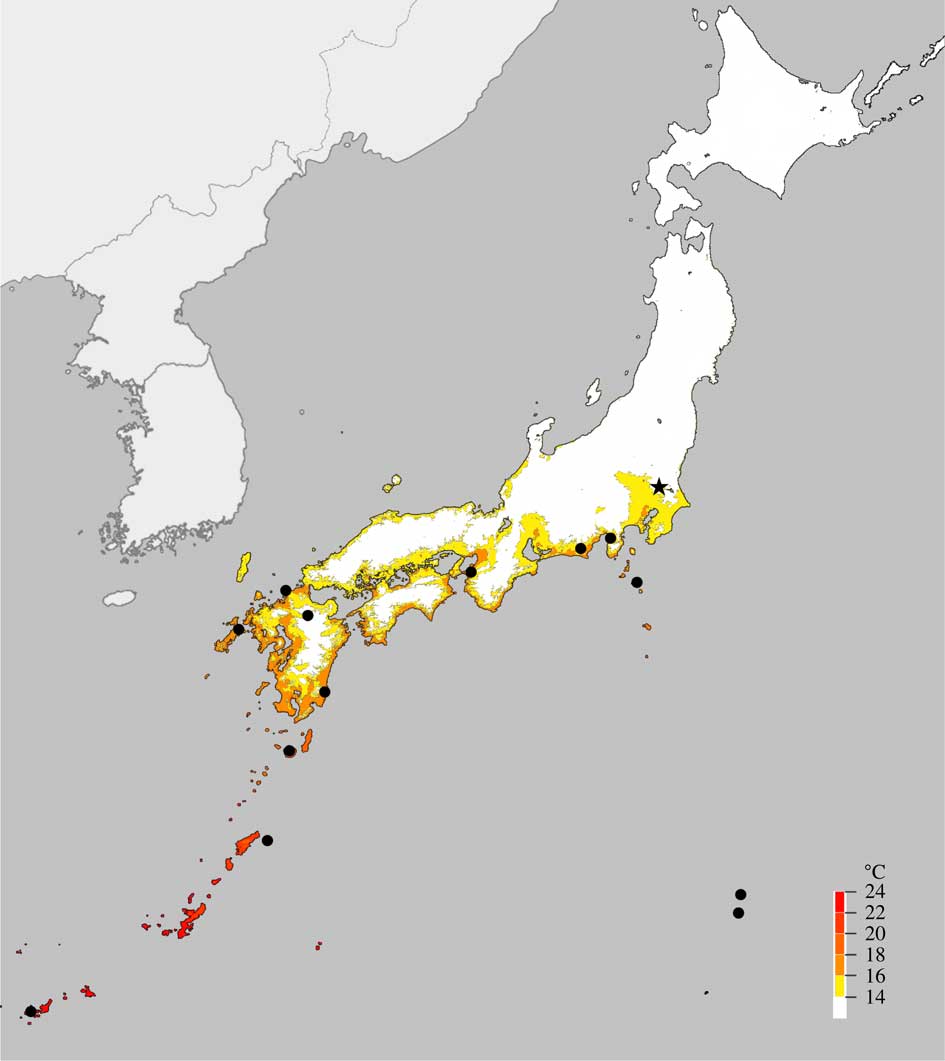
Fig. 2 Distribution map of Glyphis cicatricosa in Japan, with records from Tsukuba-city (★) and other localities (●); mean annual temperature from 1981 to 2010 delineated (Japan Meteorological Agency; http://www.jma.go.jp/).

Fig. 3 Glyphis cicatricosa, Tsukuba Botanical Garden. A, habitat; B, location on Phellodendron amurense indicated by arrow, associated mainly with Graphis handelii; C, thallus. Scale: C=2 mm.
The distinctive phytogeographical distribution pattern of pantropical species in Japan is delimited south of the 15°C annual isothermal line (with a mean annual minimum of −3·5°C), known as the ‘Crinum Line’; this is based on the distribution of Crinum asiaticum L. var. japonicum Baker, a vascular plant belonging to the Amaryllidaceae (Koshimizu Reference Koshimizu1938). Many organisms have similar distribution patterns in Japan including pantropical lichens which, according to Sato (Reference Sato1959), are generally distributed in the warmer areas to the south of the 15°C annual isothermal line. Glyphis cicatricosa in Japan (Fig. 2) is clearly pantropical, with a mainly tropical and subtropical distribution and only sporadic records from adjacent warm-temperate regions (see Kurokawa Reference Kurokawa2006). Other pantropical lichen species in Japan are represented by, for example, Dictyonema sericeum (Sw.) Berk., D. moorei (Nyl.) Henssen, Lecanora leprosa Feé, Sticta gracilis (Müll. Arg.) Zahlbr. and S. duplolimbata (Hue) Vain. (Sato Reference Sato1959; Kurokawa Reference Kurokawa2006). Of these pantropical species, only L. leprosa and G. cicatricosa have been found in Tsukuba-city (Ohmura Reference Ohmura2016).
The higher air temperature created in urban areas, the ‘urban heat island effect’, is well known (e.g. McCarthy et al. Reference McCarthy, Best and Betts2010; Kusaka et al. Reference Kusaka, Hara and Takane2012) but it has not been widely recognized as contributing directly to lichen distribution (Coppins Reference Coppins1973), mainly due to the overriding impact of air pollution. In arid or sub-arid climates, both air pollution and drought can affect lichen distribution, as demonstrated by Nimis (Reference Nimis1985). There have been many studies which have suggested that an increase in mean temperature, for example through ‘global warming’, will influence lichen distribution (e.g. van Herk et al. Reference van Herk, Aptroot and van Dobben2002; Seaward Reference Seaward2004; Ellis et al. Reference Ellis, Coppins, Dawson and Seaward2007, Reference Ellis, Eaton, Theodoropoulos, Coppins, Seaward and Simkin2015; Aptroot Reference Aptroot2009; Hauck Reference Hauck2009; Bengtsson & Paltto Reference Bengtsson and Paltto2013; Davydov et al. Reference Davydov, Insarov and Sundetpaev2013; Semenova et al. Reference Semenova, Morgado, Welker, Walker, Smets and Geml2015). Northward shifts in distribution in Japan, as seen in G. cicatricosa, have already been reported in various plants and animals (Kitahara et al. Reference Kitahara, Iriki and Shimizu2001; Murakami et al. Reference Murakami, Matsui and Morimoto2007; Japan Weather Association 2013). Therefore, the relatively recent occurrence of G. cicatricosa, as well as L. leprosa, in Tsukuba-city could be explained in terms of ‘global warming’, exacerbated by an ‘urban heat island’ effect. However, the documented distribution shifts for lichens that can be linked to global warming effects are still rather scanty, and corroborative reports for a few species, as above, have some severe limitations in the degree to which they can be generalized. Due to dramatic reductions in air pollution in many cities worldwide, as exemplified by Tsukuba-city (Fig. 4) and other cities in Japan (Ohmura et al. Reference Ohmura, Kawachi, Ohtara and Sugiyama2008, Reference Ohmura, Kashiwadani and Moon2012, Reference Ohmura, Thor, Frisch, Kashiwadani and Moon2014), lichen distribution patterns no longer demonstrate a clear relationship with sulphur dioxide. This leads to a multivariate process involving urban heat islands which potentially represent higher latitude refugia for the colonization of species that are shifting their distributions as a consequence of global warming. Investigating this phenomenon is facilitated by declining levels of pollution in urban areas where lichen recolonization is of particular interest to biogeographers.
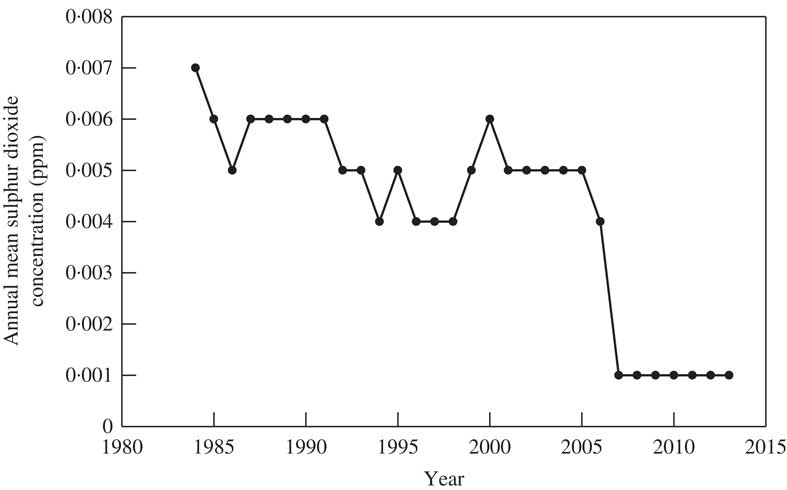
Fig. 4 Annual mean concentration of sulphur dioxide, 1984 to 2013 measured at an air pollution monitoring station located 8·1 km SE of the Botanical Garden (National Institute for Environmental Studies; http://tenbou.nies.go.jp/gis/).
Representative specimens examined. Japan: Honshu, Prov. Hitachi (Pref. Ibaraki): Tsukuba Botanical Garden, Amakubo, Tsukuba-city (36°06'03''N, 140°06'43''E), on bark of Phellodendron amurense, 20 m elev., 2015, Y. Ohmura 11001 (TNS). Prov. Izu (Pref. Shizuoka): Mishima, 1928, Y. Asahina 196 (TNS). Prov. Tohtomi (Pref. Shizuoka): Saruwatari, Aigusa-mura, Ogasa-gun, 7 vii 1932, G. Hashimoto s. n. (TNS). Prov. Izumi (Pref. Osaka): Kami-Kyoshi, Kyoshi-mura, Semnan-gun, 1954, M. Togashi 549056 (TNS). Kyushu, Prov. Buzen (Pref. Fukuoka): Nakatsumiya, Ohshima, Munakata-gun, on tree bark, 20 m elev., 8 viii 1960, W. Yoshitake s. n. (TNS). Prov. Bungo (Pref. Ôita): Nishi-Arita-mura, Hita-gun, 21 ii 1922, Nakayama s. n. (TNS). Prov. Hyuga (Pref. Miyazaki): Nakaura, Nishi-Benbun, Nichinan, 1955, S. Kurokawa 550087 (TNS); Uto Shrine, Minami-Naka-gun, 1955, S. Kurokawa 550117 (TNS). Izu Islands (Tokyo Metropolis): Tairo-pond, Miyake Island, 1976, H. Kashiwadani 13710 (TNS). Bonin Islands (Tokyo Metropolis): Minami-Fukurozawa, Chichi-jima Island, 1977, H. Kashiwadani 14138, 14144 (TNS); Sekimon, Haha-jima Island, on twig of Morus australis, 250 m elev., 2013, Y. Ohmura 9794 (TNS). Ryukyu Islands (Pref. Okinawa): along the river, Nakama-gawa, Iriomote Island, Yaeyama Islands, 0–50 m elev., 1973, I. Yoshimura 248 (TNS); Sakamine, Kikai-jima, on bark of Casuarina equisetifolia, 1988, H. Kashiwadani 35650 (TNS).
Specimens noted in references but not examined. Japan: Prov. Hizen (Pref. Nagasaki): Arikawa, Arikawa-cho, 120 m elev., [undated], M. Nakanishi 9644 (HIRO, Nakanishi Reference Nakanishi1966). Prov. Ohsumi (Pref. Kagoshima): M. Nagata, Yakushima-seihokubu, [undated], M. Oshio 9154 (HIRO, Nakanishi Reference Nakanishi1966).
The authors are most grateful to Dr N. Tanaka and Ms T. Tanimura of the Tsukuba Botanical Garden for habitat information, to Dr M. Nakanishi for providing an additional specimen and valuable comments, and to anonymous referees for their helpful criticism of an earlier draft of this paper. This study is partly supported by JSPS KAKENHI Grant no. 24300314.





 Open access
Open access