



Introduction
Fishes of the order Gadiformes (cods, hakes, and allies) include more than 600 living species arranged in 84 genera and 13 families, many of which are of high commercial importance, constituting over one quarter of the world's marine fish catch (Cohen et al., Reference Cohen, Inada, Iwamoto and Scialabba1990; Nelson et al., Reference Nelson, Grande and Wilson2016). Although morphological and molecular studies support the monophyly of the Gadiformes (e.g., Endo, Reference Endo2002; Roa-Varón and Ortí, Reference Roa-Varón and Ortí2009), the within-group relationships remain problematic to establish (e.g., Grand et al., Reference Grand, Zaragüeta Bagils, Vélez and Ung2014; Hughes et al., Reference Hughes2018). Codfishes are widespread in all the oceans from polar to tropical waters, ranging from deep-sea, neritic, estuarine, and freshwater habitats (Cohen et al., Reference Cohen, Inada, Iwamoto and Scialabba1990; Nelson et al., Reference Nelson, Grande and Wilson2016). They are characterized by a set of morphological features (which are not unique to gadiforms), including a scapular foramen present between the scapula and coracoid, absence of basihyal, premaxilla forming the entire margin of upper jaw, toothless ectopterygoid, absence of orbitosphenoid and basisphenoid, hyomandibula with a single condyle, and lack of swim bladder or pneumatic duct (Endo, Reference Endo2002; Nelson et al., Reference Nelson, Grande and Wilson2016). Moreover, codfishes are unique in having a pince-nez-shaped saccular otolith often with a collicular crest above the ostium-cauda junction of the crista inferior (Endo, Reference Endo2002; Borden et al., Reference Borden, Grande, Smith, Arratia, Schultze and Wilson2013; Grande et al., Reference Grande, Borden, Smith, Arratia, Schultze and Wilson2013).
Although a Cretaceous origin has been hypothesized from their sister-group relationships with zeiforms, reliable occurrences of fossil gadiforms are known only from Cenozoic deposits and include otoliths, isolated bones, and articulated skeletons (see Cohen, Reference Cohen, Moser, Richards, Cohen, Fahay, Kendall and Richardson1984; Nolf and Steurbaut, Reference Nolf, Steurbaut and Cohen1989; Carnevale, Reference Carnevale2007; Kriwet and Hecht, Reference Kriwet and Hecht2008; Carnevale et al., Reference Carnevale, Harzhauser and Schultz2012; Carnevale and Harzhauser, Reference Carnevale and Harzhauser2013; Přikryl et al., Reference Přikryl, Brzobohatý and Gregorová2016; Schwarzhans et al., Reference Schwarzhans, Carnevale, Bratishko, Japundžić and Bradić2017; Přikryl, Reference Přikryl2018). The Cretaceous origin of this group seems to be also supported by the presence of a precocious bipolar distribution of their occurrences in the early Paleogene, with gadoids in the Paleocene of North Atlantic and North Sea Basin and macrouroids in the Southern Ocean (Kriwet and Hecht, Reference Kriwet and Hecht2008). In the Eocene, gadoids and macrouroids are represented in North America and several sites of Europe, including both Boreal and Tethyan localities, mostly by otoliths and isolated bones (see Kriwet and Hecht, Reference Kriwet and Hecht2008). The aim of this paper is to report a new occurrence of this group of fishes from middle to late Eocene sediments of the Sverdlovsk and Tyumen regions, Russia. Comparisons with other Eocene gadiforms from almost coeval deposits are provided, and paleobiogeographic implications are subsequently discussed.
Geological setting
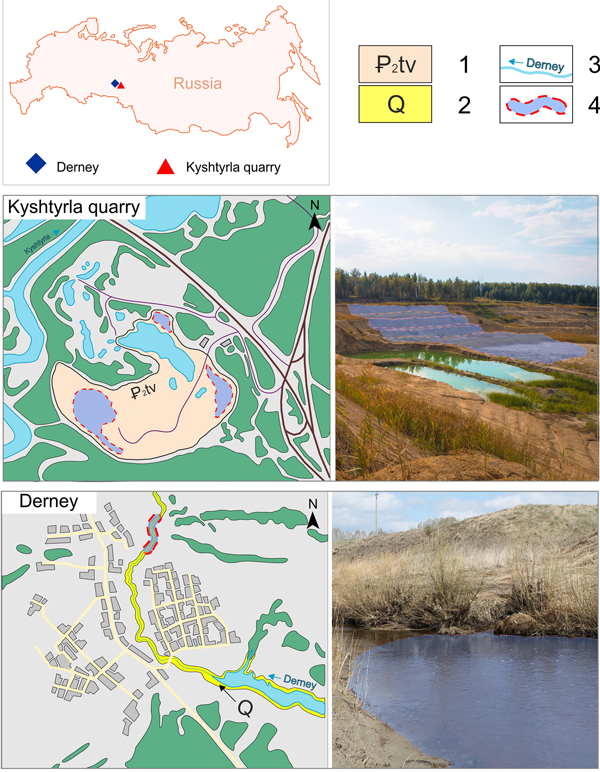
The material examined comes from two localities of the Trans-Uralian region and Western Siberia, Russia (Fig. 1). The Eocene sediments of these localities are part of the Tavda Formation, a widespread unit accumulated in the West Siberian basin during the Bartonian and Priabonian (middle to late Eocene) and established in 1944 by A. Bogdanovich in a pit near the Tavda town (Martynov et al., Reference Martynov, Sigov, Chirva, Grossgeim and Korobkov1975; Shatskii, Reference Shatskii and Shatsky1978). During the middle–late Eocene, the West Siberian Basin was a large epicontinental marine basin in which the Turgay Strait probably connected the Arctic waters to the North and the Peritethys to the South, until the Eocene–Oligocene boundary (Yasamanov, Reference Yasamanov1976; Leonov, Reference Leonov1998). The Tavda Formation either overlies or slightly contacts the Irbit Formation. The deposits of the Kurtamysh and Atlym formations or continental Quaternary sediments directly overlie the Tavda Formation. The sediments consist of greenish-grey, bluish, black, and flake-like lenticular-layered montmorillonite and beidellite-mica clays, sometimes with silty and sandy clay intercalations including clayey diatomite, mica, silica, quartz, quartz-feldspar, quartz glauconite–grained sands, and siltstones. Most often, all of these mineral inclusions are confined to the base of the section and form a layer up to 15 m thick. Nodules and inclusions of siderites, marls, stress marks, phosphorite nodules, and gypsum crystals are also present. The thickness of the formation ranges from 10 to 15 m in the west to up to 220 m in the east.
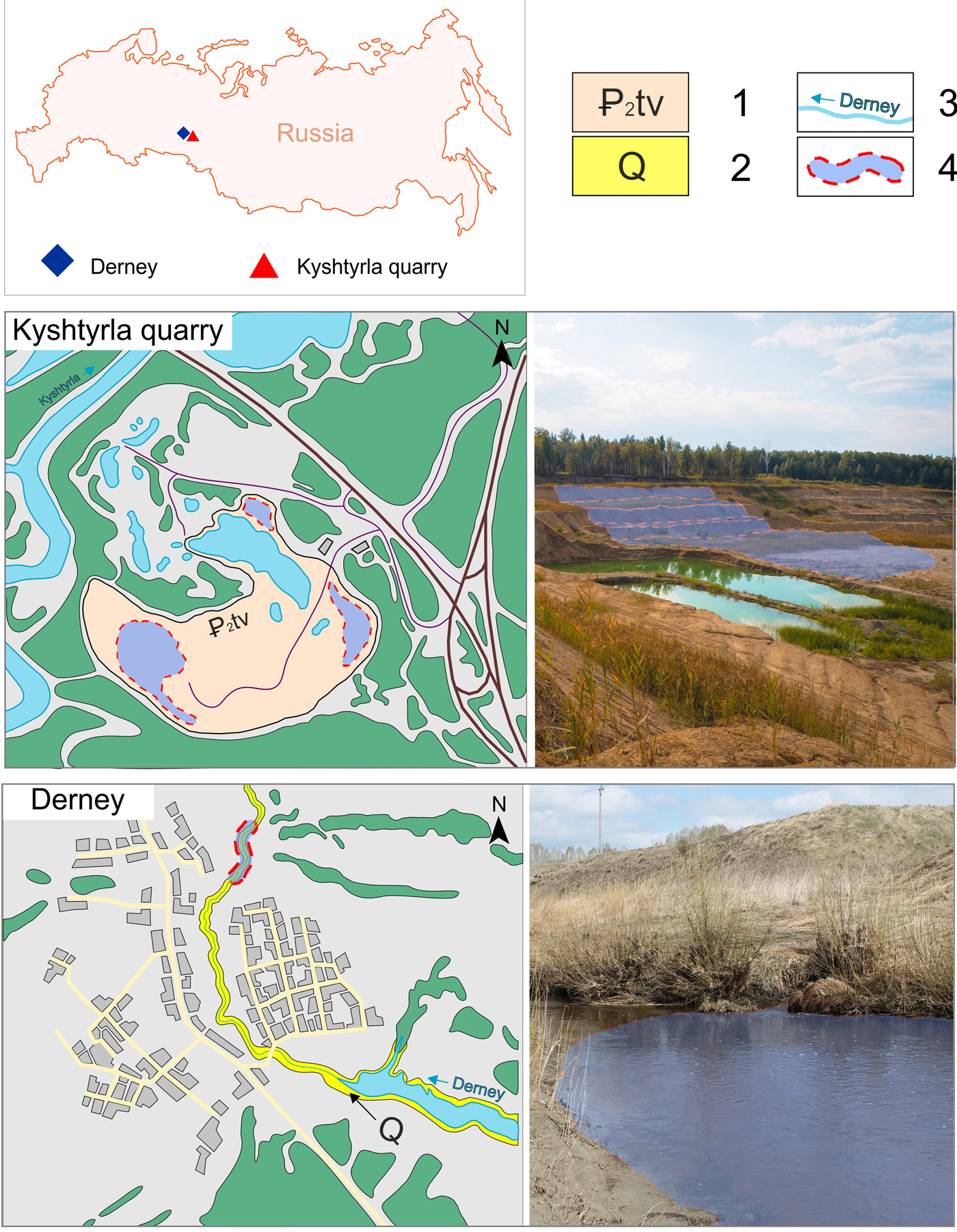
Figure 1. Location map and detailed overview of the provenance sites of the specimens. 1 = Tavda Formation outcrops in Kyshtyrla quarry; 2 = Quaternary sediments in Derney site; 3 = small rivers; 4 = sampling sites.
The Tavda Formation is usually divided into lower and upper subformations. The lower subformation (20–100 m) is represented by greenish silty clay with thin interlayers (1–2 mm) of light grey sand and siltstone and is characterized by the Rhombodinium draco assemblage. According to recent studies on organic-walled microphytoplankton, planktonic and benthic foraminifera, the lower Tavda subformation is dated to the Bartonian (Akhmet'ev et al., Reference Akhmet'ev, Aleksandrova, Beniamovskii, Vitukhin and Glezer2004; Vasilyeva et al., Reference Vasilyeva, Malyshkina and Amon2006; Vasilyeva, Reference Vasilyeva2015). The upper Tavda subformation (30–50 m), composed of greenish aleurite thin-layer clay with layers of clayey siderite, belongs to the Charlesdowinea clathrata angulosa assemblage, dating back to the Priabonian (Iakovleva et al., Reference Iakovleva, Rosseau and Cavagnetto2000; Akhmet'ev et al., Reference Akhmet'ev, Aleksandrova, Beniamovskii, Vitukhin and Glezer2004; Akhmet'ev and Beniamovski, Reference Akhmet'ev and Beniamovski2006). These data are included in the current stratigraphic scheme of the region (Unified regional stratigraphic schemes for Neogene and Paleogene deposits of the West Siberian Plain, 2001).
Kyshtyrla
Part of the material comes from Kyshtyrla, a clay pit located 4 km southeast of the village of Vinzili, 2 km northeast of the village of Kyshtyrla, 5 km northwest of the Bogandinskaya railway station, and 33 km south of the city of Tyumen (Tyumen region, Russia) (Fig. 1). From a geomorphologic point of view, the area is a hilly plain with altitude of 55–65 m confined to the first and (partially to) the second terraces of Pyshma river. The clays are exposed for about 14 m, whereas during exploratory drillings the maximum measured thickness was at least 60–80 m. The clays are covered by Quaternary clay and sandy sediments through a nondepositional unconformity. Clays in the lower part of the section form a single horizon with a thickness of up to 5 m of dark-grey, aleuritic, layered sediments. The upper part of the section is represented by a 9 m thick horizon of bluish-grey and greenish-grey aleuritic clays including small scales of mica. The associated fossils include mollusk shells, isolated shark teeth, fragments of bony fishes, and carbonaceous plant remains (Trubin, Reference Trubin2018; Popov et al., Reference Popov, Trubin, Smirnov, Ordovsky, Goncharova and Amitrov2019). Abundant bioturbation traces are also present (Nesterov et al., Reference Nesterov, Trubin, Smirnov and Yan2018).
Derney
The other locality is represented by a series of small cliffs near the Derney river in the Borovlyanskoe village (Sverdlovsk region), which is located 170 km from Yekaterinburg. The fossils from this locality are reworked from the Eocene Tavda Formation into Pleistocene blue silty clays of fluvial origin (Stefanovsky et al., Reference Stefanovsky, Zinoviev, Trofimova and Strukova2002; Malyshkina, Reference Malyshkina2003, Reference Malyshkina2006, Reference Malyshkina2012). The lower part of the section is a vivianite-rich blue clay with the fossil remains of large late Pleistocene mammals, including Mammuthus primigenius (Blumenbach, Reference Blumenbach1799), Coelodonta antiquitatis (Blumenbach, Reference Blumenbach1807), Rangifer sp., and Equus sp. (Malyshkina, Reference Malyshkina2006). The upper part of the section consists of grey quartz sands, with lenses of silty clay and coarser material, in which Eocene shark teeth and isolated bony fish remains are associated with Pleistocene tetrapods (Stefanovsky et al., Reference Stefanovsky, Zinoviev, Trofimova and Strukova2002; Malyshkina, Reference Malyshkina2006). Due to their reworked nature, the fossils are mostly represented by poorly preserved isolated remains.
Materials and methods
The fossil material was obtained during fieldwork carried out by P. Smirnov and Y. Trubin in the Kyshtyrla and Derney sites from 2014 to 2017. The specimens were collected from the surface and during the screen washing of sediments with a sieve of 1 mm mesh size. The preparation of the fossils was carried out in an alkaline solution to remove the clay material following the method of Guzhov (Reference Guzhov2009). Part of the matrix associated with fossils was mechanically removed using needles and small scalpels. The material was examined using a Leica Wild M3C stereomicroscope equipped with a camera lucida drawing arm. The specimen Kr-BF-24 was investigated noninvasively with a micro-CT device SkyScan/Bruker 1173 at the Department of Paleontology of University of Vienna, and the processing of the TIFF-image stacks was performed with Amira v.5.4.1 (Visualization Sciences Group). Measurements were taken to the nearest 0.1 mm using the software TpsDig 2.19 (Rohlf, Reference Rohlf2005). Osteological terminology mostly follows Inada (Reference Inada1981) and Endo (Reference Endo2002).
Repositories and institutional abbreviations
Fossils are kept in the Museum of the History of Science and Technology of Zauralie, Tyumen Industrial University, Russia. Specimens are labelled with Kr-BF (Kr stands for ‘collectible miscellaneous,’ ‘Коллекционное разное’ in Russian, and BF stands for ‘bony fishes’). Extant specimens used for comparisons are housed in the Evolutionary Morphology Research Group lab at the Department of Palaeontology, University of Vienna (EMRG) and Naturhistorisches Museum Wien (NMW).
Systematic paleontology
Subdivision Teleostei sensu Patterson and Rosen, Reference Patterson and Rosen1977
Order Gadiformes Goodrich, Reference Goodrich and Lankaster1909
Family Merlucciidae Gill, Reference Gill1884
Genus indeterminate, species indeterminate
Figures 2, 3.3, 4
Description
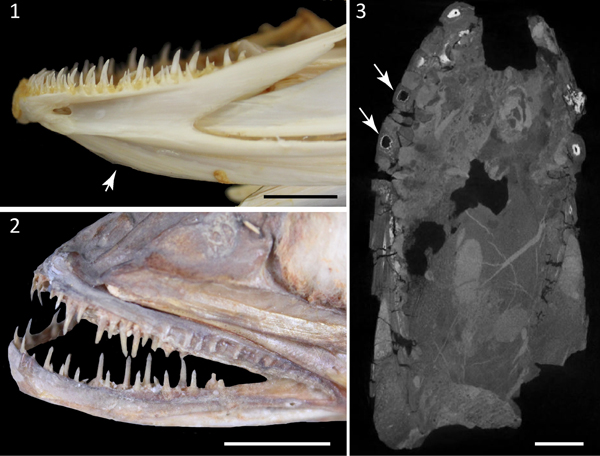
The specimen Kr-BF-24 consists of a single nodule that contains two contralateral partially articulated dentaries pertaining to a single individual (Fig. 2). The specimen is three-dimensionally preserved. The overall morphology of the lower jaws is consistent with that characteristic of the Merlucciidae (see Inada, Reference Inada1981). The dentary is large, trapezoid in shape, measuring about 93 mm in length and 46 mm in height, although its posteriormost portion seems to be absent. The dentary is narrow at the symphysis, becoming gradually higher posteriorly. When observed in occlusal or ventral view the dentaries appear moderately arched, with concave inner and convex outer margins. The ventral median keel, which is typical in several gadiforms such as Merluccius and Gadus (Fig. 3.1; see also Claeson et al., Reference Claeson, Eastman and Macphee2012), is deep, slightly higher than the dorsal part of the dentary and separated from the latter by a deep sensory groove. The pores of the mandibular sensory canal are difficult to recognize. There are two rows of large, fang-like, and regularly spaced teeth on the alveolar surface of each dentary, resembling the condition of some living gadiforms (Fig. 3.1, 3.2). The teeth are sharp, slim, and smooth, having roughly the same size throughout the jaws. Small caps of acrodin can be recognized in some teeth (Fig. 4.1), resembling the condition of some modern (e.g., Merluccius; see Fig. 3.2) and extinct merlucciids (e.g., Rhinocephalus; Rosen and Patterson, Reference Rosen and Patterson1969). Large pulp cavities can also be recognized through the micro-CT scan (Fig. 3.3). The teeth of the outer and inner rows are mesially and posteriorly curved. Being morphologically similar, a single isolated tooth (Kr-BF-11; Fig. 4.2) seems to pertain to the same group, whereas another isolated tooth (Kr-BF-9; Fig. 4.3), which is more robust and shallower, is only tentatively referred to Merlucciidae herein.
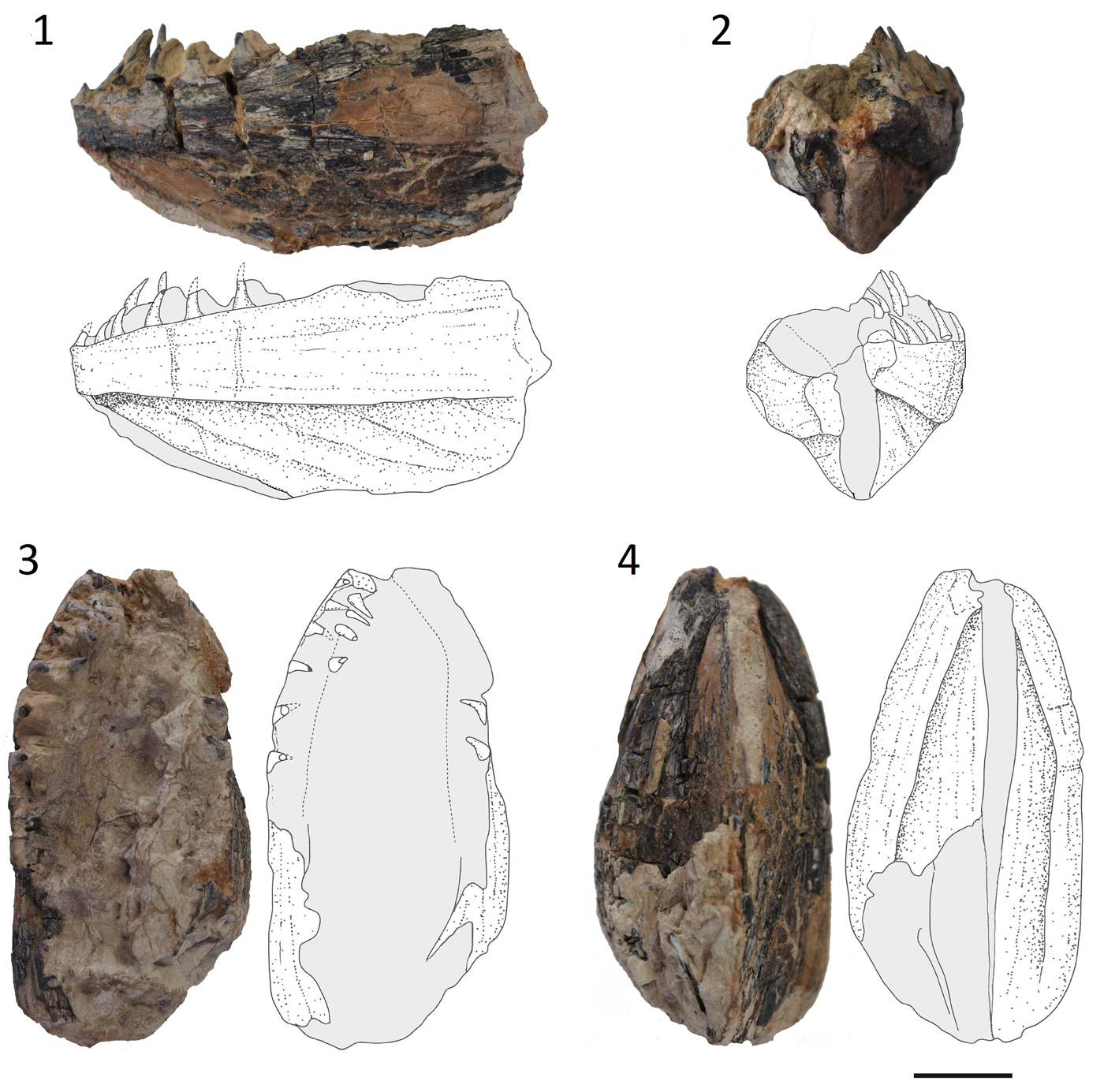
Figure 2. Specimen Kr-BF-24 with interpretative drawings. (1–4) Lower jaws (dentaries) of Merlucciidae incertae sedis in (1) lateral, (2) anterior, (3) occlusal, and (4) ventral views. Scale bar = 20 mm.
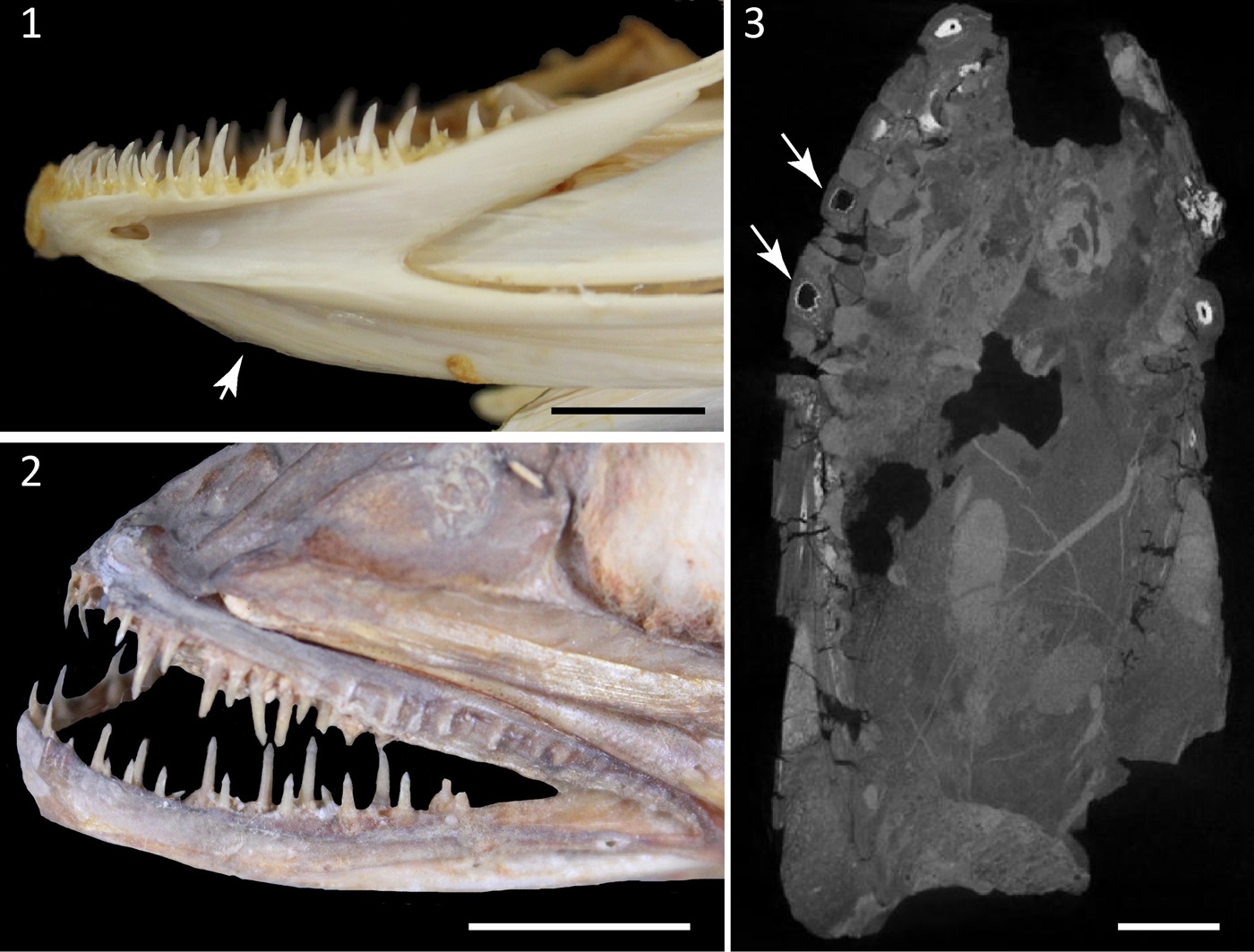
Figure 3. Comparative extant material and detail of the micro-CT scan. (1) Detail of the dentary of Gadus morhua (EMRG-Act-H-1) showing a deep ventral keel (arrow); (2) close-up of the jaws of Merluccius vulgaris (NMW 79449); (3) Kr-BF-24, image taken from micro-CT scan showing the large pulp cavities of some teeth (arrows). Scale bars = 10 mm.

Figure 4. Fossil remains referred to Merlucciidae incertae sedis. (1) Close-up of a tooth of specimen Kr-BF-24 showing the small cap of acrodin; (2) Kr-BF-11, isolated tooth; (3) Kr-BF-9, isolated tooth tentatively referred to Merlucciidae. Scale bars = 1 mm.
Materials
Kr-BF-24, a single nodule containing three-dimensionally preserved lower jaws, Kyshtyrla site; Kr-BF-9, isolated tooth, Derney site; Kr-BF-11, isolated tooth, Derney site.
Remarks
The almost complete lower jaws described herein can be aligned to the gadiform family Merlucciidae due to the presence of some characters, including a deep keel on the dentary and sharp and pointed fang-like teeth arranged in two rows and with small caps of acrodin (see Rosen and Patterson, Reference Rosen and Patterson1969; Inada, Reference Inada1981; Bieńkowska-Wasiluk et al., Reference Bieńkowska-Wasiluk, Bonde, Møller and Gaździcki2013, fig. 13). Eocene lower jaws of gadiform fishes were reported from the La Meseta Formation in Antarctica by Jerzmańska and Świdnicki (Reference Jerzmańska and Świdnicki1992) and identified as Merlucciidae by Claeson et al. (Reference Claeson, Eastman and Macphee2012), although Bieńkowska-Wasiluk et al. (Reference Bieńkowska-Wasiluk, Bonde, Møller and Gaździcki2013) assigned this material to the cod icefishes of the percomorph suborder Notothenioidei. The overall appearance of the dentaries from the middle–late Eocene of Tyumen region is quite different from the jaws from the Eocene of Antarctica and from those of other Eocene fossil gadiforms such as Rhinocephalus (Rosen and Patterson, Reference Rosen and Patterson1969), being much shallower and more elongated.
Order Gadiformes Goodrich, Reference Goodrich and Lankaster1909
Family indeterminate
Figure 5
Description
The material reported herein is tentatively referred to the Gadiformes due to the considerable similarities with the extant taxa (e.g., Brosme, Gadus, Macrouronus, Merluccius, Molva, Pollachius) as well as with fossil material from other Eocene localities (e.g., Inada, Reference Inada1981; Cohen, Reference Cohen1989; Howes, Reference Howes1991a; Endo, Reference Endo2002).

Figure 5. Fossils referred to Gadiformes incertae sedis. (1) Kr-BF-6, isolated vertebral centrum in (from left to right) anterior, dorsal, lateral, and ventral views; (2) Kr-BF-4, isolated vertebral centrum in (from left to right) anterior, dorsal, lateral, and ventral views; (3) Kr-BF-14, basioccipital in dorsal (left) and ventral views (right); (4) Kr-BF-3, vomer in dorsal (left) and ventral views (right). Scale bars = 2 mm.
The overall morphology of the vertebrae is very similar to that of the gadiform centra from the Eocene of La Meseta Formation, Antarctica, described and figured by Jerzmańska (Reference Jerzmańska1988) and Jerzmańska and Świdnicki (Reference Jerzmańska and Świdnicki1992) (compare also Tercerie et al., Reference Tercerie, Bearez, Pruvost, Bailly and Vignes-Lebbe2016). The vertebrae are massive and well ossified, strongly amphicoelous, subrectangular in outline, and higher than long (Fig. 5.1, 5.2). Distinct growth rings can be recognized on both anterior and posterior faces. Only the base of neural arches is preserved, suggesting that they were fused to centra. The vertebrae Kr-BF-5, Kr-BF-6, and Kr-BF-7 pertain to the abdominal portion of the body since they are trapezoid in cross section with their minimum and maximum width at their dorsal and ventral margins, respectively. Part of a large parapophysis is preserved on the ventral surface of the vertebra Kr-BF-6 (Fig. 5.1), suggesting that it might have been part of the posterior portion of the abdominal region. Unfortunately, due to inadequate preservation, it is not possible to detect the length of the base of the parapophysis. Along the dorsal surface, two narrow but deep grooves are separated by a thin bony lamella (Fig. 5.1, 5.2), whereas two or three grooves can be recognized on the lateral sides. A single narrow groove is present on the ventral surface of all the vertebrae, and broad and deep depressions can be observed medially to the base of the right and left parapophyses (Fig. 5.1). Prezygapophyses are mostly not preserved, but small and fan-shaped postzygapophyses are visible in Kr-BF-4 (Fig. 5.2; arrow). The large size of the vertebrae (about 10 mm in diameter) might indicate that they were from large individuals.
The material also includes an isolated basioccipital (Kr-BF-14; Fig. 5.3), which is elongate and measures about 9 mm in length and 4 mm in width. In dorsal view, it is possible to recognize an irregular face for the articulation with the exoccipitals. In posterior view, the facet for the articulation with the first centrum is ovoid in shape. In ventral view, there is no trace of the sutural surface for the parasphenoid, suggesting that the bone is incomplete and some millimeters are missing from its anterior edge.
The specimen Kr-BF-3 seems to represent a part of a vomer (Fig. 5.4). It measures about 13 mm in length, but it is likely that most of the posterior part of the bone is missing. The head of the vomer is robust and considerably widened, whereas its ventral margin is apparently toothless, resembling the condition of Muraenolepis and some morids (e.g., Danilchenko, Reference Danilchenko1967; Endo, Reference Endo2002), although we cannot exclude that this might be a taphonomic artefact. The lateral wings of the vomer are represented by tiny sheets of bones separating the dorsal from the ventral sides.
Materials.—Kr-BF-4, Kr-BF-5, Kr-BF-6, and Kr-BF-7: isolated vertebral centra, Kyshtyrla site; Kr-BF-15, and Kr-BF-17: isolated vertebral centra, Derney site; Kr-BF-14: a single basioccipital, Derney site; Kr-BF-3: partial vomer, Kyshtyrla site.
Remarks
Isolated vertebral centra belonging to indeterminate gadiforms were also reported from the Eocene of the La Meseta Formation in Antarctica by Jerzmańska (Reference Jerzmańska1988) and Jerzmańska and Świdnicki (Reference Jerzmańska and Świdnicki1992). Nevertheless, the attribution of the Antarctic vertebrae to this group of bony fishes has been questioned by Bieńkowska-Wasiluk et al. (Reference Bieńkowska-Wasiluk, Bonde, Møller and Gaździcki2013), who pointed out that strong parapophyses and the presence of several lateral bony lamellae are shared by gadiforms and some nothenioids. However, our comparison of the Eocene vertebrae from Russia and those figured by Tercerie et al. (Reference Tercerie, Bearez, Pruvost, Bailly and Vignes-Lebbe2016) shows that several gadiforms (including Gadus, Macruronus, and Merluccius) do exhibit abdominal and caudal vertebral centra higher than long, therefore supporting our attribution of the vertebrae from the Eocene of Russia to the Gadiformes.
Origin and early paleogeographic distribution
The oldest reliable gadiform remains are from Paleocene deposits and include about 20 occurrences from 10 localities (Fig. 6). More precisely, the oldest known gadiform appears to be a partially complete articulated gadoid skeleton from the Danian of West Greenland (Rosen and Patterson, Reference Rosen and Patterson1969; Cohen, Reference Cohen, Moser, Richards, Cohen, Fahay, Kendall and Richardson1984). Early gadoids also are known from isolated otoliths recorded from the Paleocene–early Eocene of Austria, Belgium, Denmark, Germany, Greenland, United Kingdom, Ukraine, and United States (Schwarzhans, Reference Schwarzhans2003, Reference Schwarzhans2004, Reference Schwarzhans2012; Schwarzhans and Bratishko, Reference Schwarzhans and Bratishko2011). Macrouroid otoliths were reported from the Paleocene of southern Australia and New Zealand (e.g., Schwarzhans, Reference Schwarzhans1985), possibly indicating a Southern Hemisphere origin for this group and a subsequent migration to the Northern Hemisphere through the northern margin of Antarctica and an Atlantic corridor (Kriwet and Hecht, Reference Kriwet and Hecht2008) (Fig. 6). This might suggest that two origination and radiation events of modern gadiform lineages occurred in the early history of gadiforms, with gadoids originating and evolving in the Northern Hemisphere, while macrouroids diversified in the Southern Hemisphere (Kriwet and Hecht, Reference Kriwet and Hecht2008). This already wide and distinct distribution in the Paleocene might indicate that the origin of gadiforms possibly occurred in the Late Cretaceous, although no reliable records from the Mesozoic have been reported so far (see Svetovidov, Reference Svetovidov1948; Fedotov and Bannikov, Reference Fedotov, Bannikov and Cohen1989; Kriwet and Hecht, Reference Kriwet and Hecht2008; Carnevale and Johnson, Reference Carnevale and Johnson2015). The gadoid migration into the Southern Hemisphere possibly occurred very early in their evolutionary history, likely in relation to oceanic adaptations (Kriwet and Hecht, Reference Kriwet and Hecht2008) and following the same corridor suggested for other Eocene marine fish lineages (Long, Reference Long1994; Marramà et al., Reference Marramà, Engelbrecht, Mörs, Reguero and Kriwet2018a, Reference Marramà, Schultz and Kriwetb). Howes (Reference Howes1991b) suggested that gadoids migrated from oceanic to shelf habitat during their early evolution. However, the fossil record seems to contradict this hypothesis since most of the early gadoids are from shelf areas of the North Sea Basin with a maximum depth of about 250 m during the Paleocene (Gemmer et al., Reference Gemmer, Huuse, Clauden and Nielsen2002).
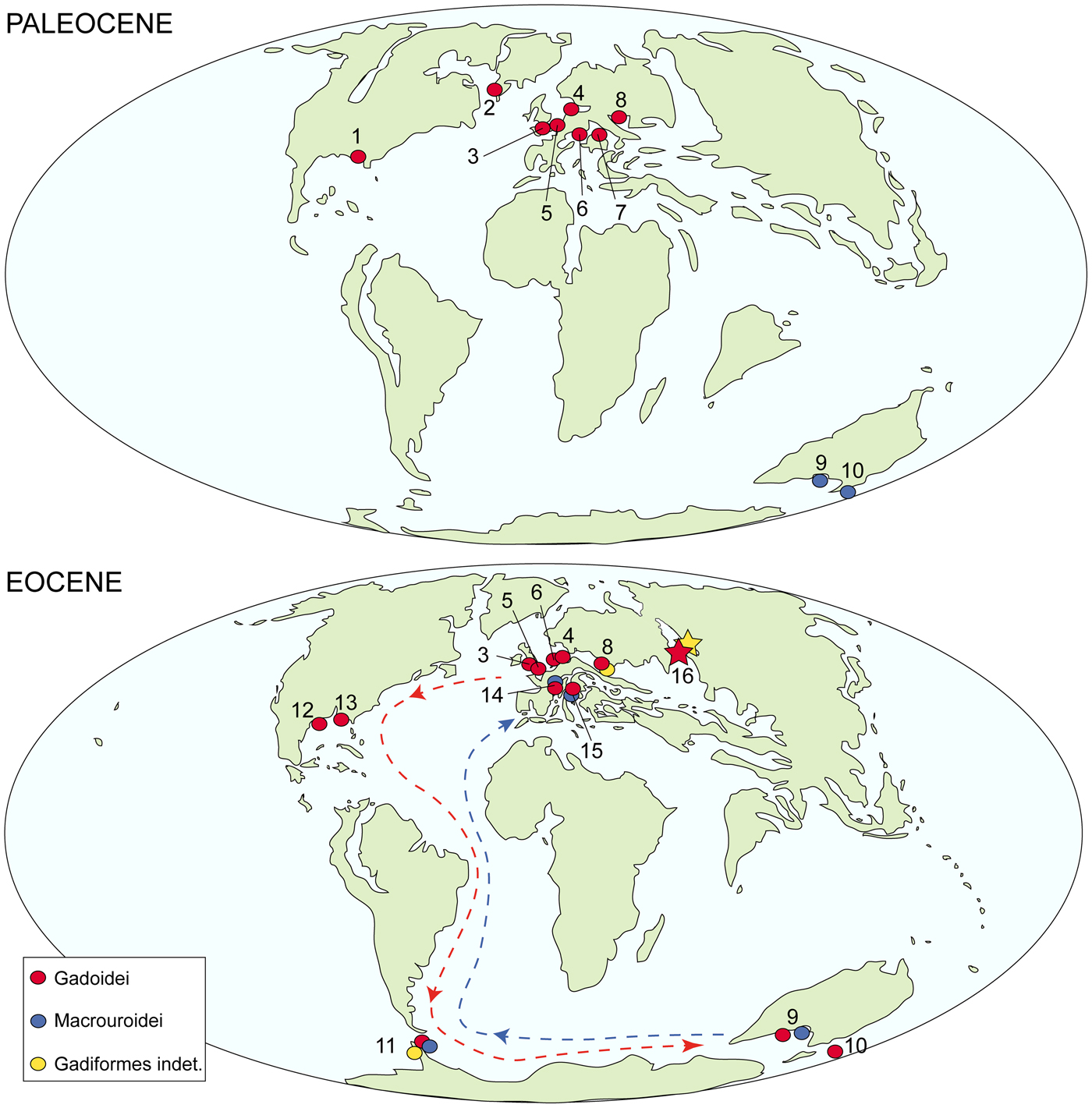
Figure 6. Paleobiogeographic distribution of gadiform occurrences in the Paleocene and Eocene according to Schwarzhans (Reference Schwarzhans1985, Reference Schwarzhans2012), Nolf and Dockery (Reference Nolf and Dockery1993), Kriwet and Hecht (Reference Kriwet and Hecht2008), and Schwarzhans and Bratishko (Reference Schwarzhans and Bratishko2011). The arrows indicate the possible migration routes of gadoids (red) and macrouroids (blue) that occurred in the Eocene. The stars represent the occurrences from Tyumen and Sverdlovsk regions reported in this study. Maps are modified from Scotese (Reference Scotese2002). 1 = Alabama; 2 = Greenland; 3 = England; 4 = Denmark; 5 = Belgium; 6 = Germany; 7 = Austria; 8 = Ukraine; 9 = Australia; 10 = New Zealand; 11 = Antarctica; 12 = Texas; 13 = Louisiana; 14 = France; 15 = Italy; 16 = Russia.
Conclusions
About 50 gadiform occurrences from more than a dozen localities have been reported in the Eocene (Kriwet and Hecht, Reference Kriwet and Hecht2008). In this period, gadiforms were already widespread and well diversified in both the Northern and Southern Hemispheres, and their skeletal remains and otoliths have been reported from Europe, North America, Oceania, and Antarctica up to now (e.g., Schwarzhans, Reference Schwarzhans1980, Reference Schwarzhans1981; Bonde et al., Reference Bonde, Andersen, Hald and Jakobsen2008, Reference Bonde, Madsen, Schultz, Sylvestersen and Jakobsen2011; Kriwet and Hecht, Reference Kriwet and Hecht2008). The present report of merlucciid and indeterminable gadiform remains from the middle–late Eocene of the Trans-Urals Russia extends the paleogeographic distribution of this group of fishes in the Northern Hemisphere eastward during their early radiation, providing evidence of the existence of a gadoid contingent in the Turgay Strait just before its definitive closure.
Acknowledgments
The authors thank V. Alifanov (Paleontological Institute, Russian Academy of Sciences) for the useful suggestions and assistance to preparation and B. Riedel (Naturhistorisches Museum Wien) for the access to comparative material under her care. Thanks are also due to C. Pfaff (University of Vienna) for performing micro-CT scans of the specimen Kr-BF-24 and to J. Cawley (University of Vienna) for improving the English of our manuscript. The manuscript has been greatly improved by the valuable revisions of T. Přikryl (Czech Academy of Sciences), by an anonymous reviewer, and by editorial comments of A. Murray and H.-D. Sues. Financial support was provided by the Austrian Science Fund (FWF) [M2368-B25 to G.M.] and Università degli Studi di Torino [ex-60% 2017 and 2018 grants to G.C.]. Open access funding provided by Austrian Science Fund (FWF).





 Open access
Open access