



Introduction
The first appearance datum (FAD) of the genus Belemnella Nowak, Reference Nowak1913 – specifically Belemnella lanceolata (von Schlotheim, Reference Von Schlotheim and Leonhard1813) – has traditionally been used to define the base of the Maastrichtian stage in the Boreal Realm in Central and Western Europe. In most areas of Europe, Belemnella almost completely replaced the genus Belemnitella d’Orbigny, Reference Orbigny1840 during the latest Campanian, but in areas like the Middle Vistula River region (Poland) and the Maastricht region (the Netherlands), Belemnella coexisted with Belemnitella during the Early Maastrichtian (Kongiel, Reference Kongiel1962; Keutgen & van der Tuuk, Reference Keutgen and Van der Tuuk1991; Remin, Reference Remin2012, Reference Remin2015). The disappearance of Belemnella during the mid-Maastrichtian is not well understood and – because considerable progress has been made in the correlation of sections especially in Western and Central Europe applying δ13C stable isotope stratigraphy (e.g. Niebuhr et al., Reference Niebuhr, Hampton, Gallagher and Remin2011; Thibault et al., Reference Thibault, Harlou, Schovsbo, Schiøler, Minoletti, Galbrun, Lauridsen, Sheldon, Stemmerik and Surlyk2012; Voigt et al., Reference Voigt, Gale, Jung and Jenkyns2012; Linnert et al., Reference Linnert, Engelke, Wilmsen and Mutterlose2019; Wilmsen et al., Reference Wilmsen, Engelke, Linnert, Mutterlose and Niebuhr2019; Vellekoop et al., Reference Vellekoop, Kaskes, Sinnesael, Huygh, Déhais, Jagt, Speijer and Claeys2022) – there is growing evidence that it did not happen contemporaneously everywhere (see Discussion). It is plausible that a combination of environmental factors, such as temperature changes, sea-level fluctuations and/or modifications of ocean currents, played a significant role (Remin, Reference Remin2018; Remin et al., Reference Remin, Cyglicki and Niechwedowicz2022a, Reference Remin, Krzywiec, Stachowska, Walaszczyk and Todes2022b). During the late Maastrichtian, representatives of the genus Neobelemnella Naidin, Reference Naidin, Menner, Moskvin, Naidin, Solovyev and Shimansky1975 successively expanded their area of distribution from east to west, displacing representatives of the genus Belemnitella. In order to complicate the situation further, there are currently two competing interpretations for the origin of Neobelemnella, namely that it evolved either locally from a Russian or Central Asian species of the genus Belemnella or represents a descendent from Belemnitella/?Neobelemnella subfusiformis a latest Campanian to earliest Maastrichtian species from New Jersey, western USA (Keutgen & Keutgen, Reference Keutgen and Keutgen2020).
As a matter of fact, belemnite research is in a state of flux these days because advances involving AI-technology have fundamentally questioned earlier systematic approaches. Remin (Reference Remin2012, Reference Remin2015) developed an innovative concept of belemnite determination and overcame the problem of the a priori subdivision of belemnite guards into species before statistical treatments (Christensen, Reference Christensen1975, Reference Christensen1995; Schulz, Reference Schulz1979). The artificial neural networks approach, the self-organising Kohonen algorithm creates groups of similar specimens (based on measurable features = similar input data) that may be regarded as so-called morphogroups, which may be interpreted as being composed of representatives of a paleontological/biological species. Remin developed a concept that allows determining guards of Belemitella, Belemnella, Neobelemnella and, possibly, also Fusiteuthis Kongiel, Reference Kongiel1962 applying the same approach. For the genus Belemnella, it turned out that the methods of Remin (Reference Remin2012) and Schulz (Reference Schulz1979) resulted in different species concepts (Schulz, Reference Schulz1979; Niebuhr et al., Reference Niebuhr, Hampton, Gallagher and Remin2011; Remin, Reference Remin2012; Keutgen et al., Reference Keutgen, Remin and Walaszczyk2012). Even more significant, Remin’s (Reference Remin2018) study of the belemnites from the upper lower Maastrichtian of Hrebenne (southeast Poland) revealed the presence of two species, Belemnella praearkhangelskii Naidin, Reference Naidin1964a and Belemnella kajnarensis Naidin, Reference Naidin1964a, which cannot be separated applying the systematic approach of Schulz (Reference Schulz1979). Both results substantially question determinations of Belemnella in Western and Central Europe based on the system of Schulz (Reference Schulz1979). In addition, the determinations of eastern European belemnite species in studies such Naidin (Reference Naidin1952, Reference Naidin1964a, 1974) and Baraboshkin et al., (Reference Baraboshkin, Guzhikov, Aleksandrova, Fomin, Pokrovskiy, Grishchenko, Manikin and Naumov2021, Reference Baraboshkin, Guzhikov, Aleksandrova, Akinin, Ryabov, Ustinova, Rtischev and Vishnevskaya2024) are difficult to assess because the basis of their systematic approach is not clear, usually not representing a population statistical concept at all.
It follows that for a thorough review of Belemnella, only the fauna of the Middle Vistula River valley is currently available, supplemented by sections in the immediate vicinity. The composed Middle Vistula valley section is an ideal reference due to its central location between Eastern and Western Europe because influences from both regions can be expected having an impact on the Middle Vistula belemnite fauna.
In order to compare the first and last appearance datum (LAD) of belemnites in different regions, carbon isotope data are used for correlation purpose when available (e.g. Niebuhr et al., Reference Niebuhr, Hampton, Gallagher and Remin2011; Thibault et al., Reference Thibault, Harlou, Schovsbo, Schiøler, Minoletti, Galbrun, Lauridsen, Sheldon, Stemmerik and Surlyk2012; Voigt et al., Reference Voigt, Gale, Jung and Jenkyns2012; Linnert et al., Reference Linnert, Engelke, Wilmsen and Mutterlose2019; Wilmsen et al., Reference Wilmsen, Engelke, Linnert, Mutterlose and Niebuhr2019), otherwise calcareous nannofossils are used. In the Kronsmoor quarry ‘Saturn’, the base of the Maastrichtian corresponds to the marl layer mb609, a horizon close to the base of the Belemnella obtusa Zone, 12.5 m above flint layer F600 (Wilmsen et al., Reference Wilmsen, Engelke, Linnert, Mutterlose and Niebuhr2019). Exactly at the same stratigraphic level the boundary was placed in the Middle Vistula River Valley section, Poland (Remin, Reference Remin2012). The potential lower-upper Maastrichtian substage boundary for years was defined by the FAD of Belemnitella junior Nowak, Reference Nowak1913 – however, the boundary has not been formally defined yet. For the convenience of this review, and for placing the belemnite record in an independent framework, reference is made to the position of the lower-upper Maastrichtian boundary as identified by Boussaha et al. (Reference Boussaha, Thibault and Stemmerik2016) in the Danish Chalk with the FAD of the calcareous nannofossil Lithraphidites quadratus Bramlette & Martini, Reference Bramlette and Martini1964, dated at that locality at c. 68.8 Myr, although the position of the FAD of L. quadratus is globally highly diachronous (Thibault et al., Reference Thibault, Harlou, Schovsbo, Schiøler, Minoletti, Galbrun, Lauridsen, Sheldon, Stemmerik and Surlyk2012, fig. 8; Boussaha et al., Reference Boussaha, Thibault and Stemmerik2016, fig. 10). When δ13C stable isotope data are available, a correlation of the lower-upper Maastrichtian boundary level as defined in the Stevns-1 borehole is here considered (Voigt et al., Reference Voigt, Gale, Jung and Jenkyns2012; Vellekoop et al., Reference Vellekoop, Kaskes, Sinnesael, Huygh, Déhais, Jagt, Speijer and Claeys2022; Dubicka et al., Reference Dubicka, Wierny, Bojanowski, Rakociński, Walaszczyk and Thibault2023), in all other cases the FAD of L. quadratus is taken as an approximation of this boundary. In addition, upper Maastrichtian belemnite records in the Boreal Realm are linked to the FADs of Nephrolithus frequens Górka, Reference Górka1957 and Cribrosphaerella daniae Perch-Nielsen, Reference Perch-Nielsen1973 (Thibault, Reference Thibault2016, fig. 6).
Genus Belemnella Nowak, Reference Nowak1913
The currently accepted official definition for the base of the Maastrichtian stage was published by Odin & Lamaurelle (Reference Odin and Lamaurelle2001) for the Global Stratotype Section and Point (GSSP) of the Campanian-Maastrichtian boundary at Tercis (France) and correlated with the marl layer mb609 in the Kronsmoor quarry ‘Saturn’ in northern Germany, close to the base of the Belemnella obtusa Zone, 12.5 m above flint layer F600 (Wilmsen et al., Reference Wilmsen, Engelke, Linnert, Mutterlose and Niebuhr2019). This position is distinctly above the classical definition of the boundary by the appearance of Belemnella at the flint layer F600 (Fig. 1). From the oldest Belemnella Zone, the uppermost Campanian Belemnella lanceolata Zone, only two species are known, Belemnella lanceolata (von Schlotheim, Reference Von Schlotheim and Leonhard1813) and Belemnella longissima Schulz, Reference Schulz1979, the earliest representatives of the latter species being known only from Kronsmoor and probably Balsvik, Sweden (Remin, Reference Remin2012). The fact that already two species are reported from the stratigraphic oldest Belemnella Zone indicates that adaptive radiation in Belemnella began early in the latest Campanian and that local species also developed in other areas, which are not discussed here (Fig. 2). For the sake of completeness, however, the Russian Belemnella licharewi Jeletzky, Reference Jeletzky1941 should be mentioned because it is often cited as one of the stratigraphically oldest Belemnella species on the Russian Platform (Christensen, Reference Christensen1997a, Reference Christensen1777b).

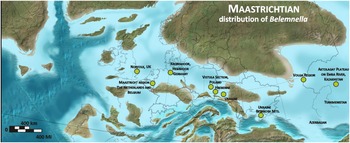
Figure 1. Stratigraphic ranges of Belemnella species and proposed belemnite zonation for the Middle Vistula valley section, central Poland; base of the Maastrichtian Stage according to the GSSP at Tercis, France (1) and as conventionally based on belemnites for the Boreal Realm (2). (L q: Lithraphidites quadratus).
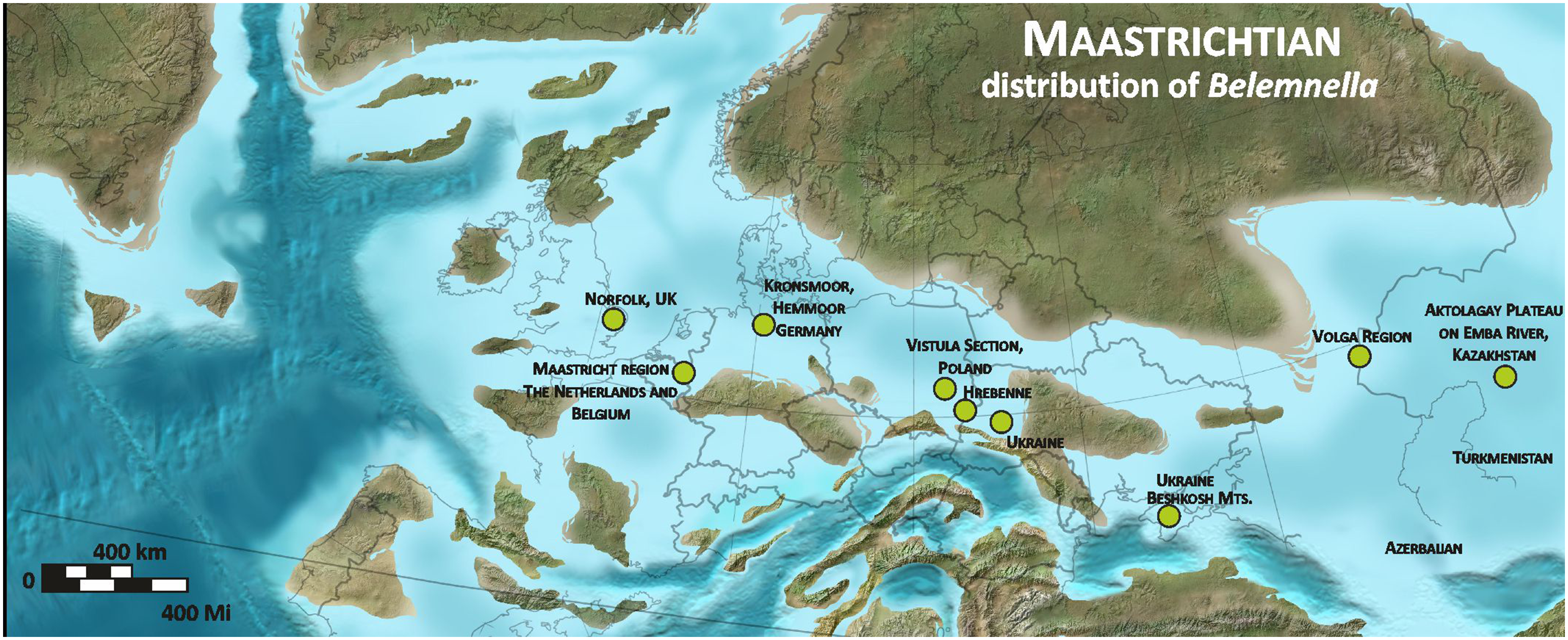
Figure 2. Paleogeographic distribution of the representatives of Belemnella. Used with permission of Colorado Plateau Geosystems Inc.Global Paleogeography and Tectonics in Deep Time © 2016.
From the stratigraphical youngest Campanian Belemnella inflata Zone already four species of Belemnella are known from the Middle Vistula section (five from Kronsmoor), of which only one species (Bl. longissima) was able to pass the Campanian-Maastrichtian boundary. Yet, four new species appeared successively within the lowermost Maastrichtian Belemnella obtusa Zone (Fig. 1). This reflects either another period of rapid adaptive radiation or alternatively migration of species from other areas into the Middle Vistula valley.
The Belemnella fauna as exposed at Hrebenne may be regarded typical of the ‘mid’ lower Maastrichtian (Fig. 1). Three species have been identified: Belemnella sumensis Jeletzky, Reference Jeletzky1949, Bl. praearkhangelskii and Bl. kajnarensis (Remin, Reference Remin2018). The index species of the Belemnella sumensis Zone has also been recorded from Boiska in the Middle Vistula Valley (Kongiel, Reference Kongiel1962 (as Bl. occidentalis), and unpublished specimens are housed in the coll. Remin). At Boiska, stratigraphically younger strata than at the Hrebenne section are exposed. Because the FAD of the calcareous nannofossil L. quadratus is also known from Boiska, a position close to the lower-upper Maastrichtian boundary can be deduced (see also Peryt et al., Reference Peryt, Dubicka and Wierny2022; Dubicka et al., Reference Dubicka, Wierny, Bojanowski, Rakociński, Walaszczyk and Thibault2023). Noteworthy, Bl. sumensis is the only species of Belemnella in the uppermost Lower Maastrichtian of the Middle Vistula River section, potentially indicating a decrease in local species diversity of Belemnella before its extinction close to the top of the Lower Maastrichtian, at least in Western and Central Europe.
In summary, four Belemnella zones can be identified in the uppermost Campanian and lower Maastrichtian of the Middle Vistula valley. The upper Campanian lanceolata and inflata zones and the Lower Maastrichtian obtusa and sumensis zones. The duration of these zones may be roughly estimated from the numerical ages deduced for northern Germany (Voigt & Schönfeld, Reference Voigt and Schönfeld2010; Thibault et al., Reference Thibault, Harlou, Schovsbo, Schiøler, Minoletti, Galbrun, Lauridsen, Sheldon, Stemmerik and Surlyk2012): lanceolata Zone c. 200 kyr; inflata Zone c. 600 kyr; obtusa Zone c. 1.4 Myr; sumensis Zone c. 2.4 Myr.
Genus Belemnitella d’Orbigny, Reference Orbigny1840
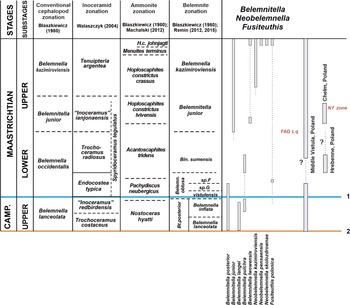
Christensen (Reference Christensen2000) distinguished two groups, the Belemnitella mucronata- and the Belemnitella langei-group. Both groups are present in the topmost Campanian Belemnella lanceolata and Belemnella inflata zones of the Middle Vistula valley (Fig. 3). Of the mucronata-group only Belemnitella posterior Kongiel, Reference Kongiel1962 is present (Keutgen & Remin, unpublished). It resembles Belemnitella carlsbergensis Christensen, Reference Christensen1998 from the lanceolata Zone of Sweden, which may represent a junior synonym. Last representatives of Belemnitella langei Jeletzky, Reference Jeletzky1948 and records of Belemnitella pulchra Schulz, Reference Schulz1982, both members of the langei-group, are known from the Middle Vistula River valley as well (Keutgen & Remin, unpublished; Fig. 3). With three species of Belemnitella, the latest Campanian Belemnella zones of the Middle Vistula section are characterised by a comparatively high species diversity.

Figure 3. Stratigraphic ranges of species of Belemnitella, Neobelemnella, and Fusiteuthis and proposed belemnite zonation for the Middle Vistula valley section, central Poland; base of the Maastrichtian Stage according to the GSSP at Tercis, France (1) and as conventionally based on belemnites for the Boreal Realm (2). (L q: Lithraphidites quadratus; N f: Nephrolithus frequens).
From the lower Maastrichtian Belemnella obtusa Zone only Bt. posterior is known from the Middle Vistula River valley (Remin, Reference Remin2015), whereas from the United Kingdom, Belgium and the Netherlands the last representatives of Belemnitella minor II are recorded (Christensen, Reference Christensen1995, Reference Christensen1999; Keutgen & van der Tuuk, Reference Keutgen and Van der Tuuk1991), in addition to rare Bt. pulchra from Belgium (Christensen, Reference Christensen1999) and northern Germany (Schulz, Reference Schulz1982).
Belemnitella species from the lower Maastrichtian Belemnella sumensis Zone are rarely recorded and belong to Belemnitella junior Nowak, Reference Nowak1913 (Kongiel, Reference Kongiel1962; Keutgen & van der Tuuk, Reference Keutgen and Van der Tuuk1991; Keutgen et al., Reference Keutgen, Jagt, Felder and Yazykova2010) and Bt. pulchra (Schulz, Reference Schulz1982; Remin, Reference Remin2018).
The Upper Maastrichtian Belemnitella junior Zone is defined in practice either by the first appearance of Bt. junior or, in case the species first co-occurs with representatives of Belemnella, by the disappearance of representatives of the latter. From the junior Zone representatives of both the mucronata-group (Bt. junior) and the langei-group (Belemnitella lwowensis Naidin, Reference Naidin1952) are known (Fig. 3). Bt. lwowensis is regarded as a descendent of Bt. pulchra, but the stratigraphic level at which the transition between the two species took place is unclear. Bt. lwowensis and Bt. junior are sporadically recorded also from the latest Maastrichtian Neobelemnella kazimiroviensis Zone (Kongiel, Reference Kongiel1962; Christensen et al., Reference Christensen, Schmid and Schulz2004). They became extinct together with all other belemnite species at the K-Pg boundary.
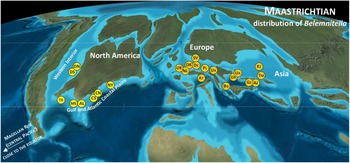
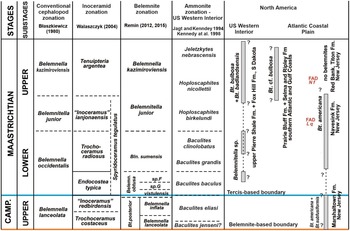
Belemnitella was characterised by a wider geographical distribution than Belemnella (Figs. 2, 4). The former is also known from Maastrichtian deposits in North America, namely from the Atlantic Coastal Plain and the Western Interior. From the Monmouth County region of New Jersey, Remin in Kopun et al. (Reference Kopun, Castro, Garb, Larina and Remin2012) identified Belemnitella americana (Morton, Reference Morton1830) and Belemnitella subfusiformis (Whitfield, Reference Whitfield1892) from the Navesink Formation, the latter species found only locally at the base of the Formation (Fig. 5). According to Sugarman et al. (Reference Sugarman, Kenneth, Bukry and Feigenson1995), the base of the Navesink Formation is situated close to the base of the calcareous nannofossil zone CC25a (UC19). The FAD of L. quadratus (base UC20a) is assumed to be within the Navesink Formation, and the FAD of N. frequens (base UC20b) is already in the lower part of the Red Bank Formation and above the stratigraphic youngest records of Bt. americana in the Navesink Formation of New Jersey, indicating its disappearance in the lowermost upper Maastrichtian at least for the Monmouth County region, where the stratigraphic range of Bt. americana is best documented. For the Western Interior, Larson (Reference Larson2010) reported rare specimens of Belemnitella cf. bulbosa Meek & Hayden, Reference Meek and Hayden1857 (Belemnitella sp. of Kennedy et al., Reference Kennedy, Landman, Christensen, Cobban and Hancock1998) from the Baculites baculus and Baculites clinolobatus zones of the Pierre Shale of South Dakota (Fig. 5). The typical Belemnitella bulbosa Meek & Hayden (Reference Meek and Hayden1857) is a rare species and regionally confined to the Fox Hills Formation and upper Pierre Shale of the Western Interior (South and North Dakota), occurring throughout the Timber Lake facies (Jeletzkytes nebrascensis Zone) and the Trail City facies (Hoploscaphites nicolletii Zone). Remin in Landman et al. (Reference Landman, Remin, Garb and Chamberlain2013) distinguished two belemnite species in the Enning facies at Badlands National Park, South Dakota: Bt. bulbosa and Belemnitella badlandsensis Landman et al., Reference Landman, Remin, Garb and Chamberlain2013. Both species from the nebrascensis Zone differ mainly in the observed range of their fissure angles.

Figure 4. Paleogeographic distribution of the representatives of Belemnitella. Letter symbols are standard symbols for countries and states (US). Used with permission of Colorado Plateau Geosystems Inc.Global Paleogeography and Tectonics in Deep Time © 2016.

Figure 5. Ammonite, belemnite, and inoceramid bivalve stratigraphy for the topmost Campanian and Maastrichtian in selected sections in the US Western Interior and Gulf and Atlantic Coastal Plains. (L q: Lithraphidites quadratus; N f: Nephrolithus frequens).
For the sake of completeness, it may be added that Zakharov et al. (Reference Zakharov, Pletnev, Melnikov, Smyshlyaeva, Khudik, Evseev, Punina and Popov2007, Reference Zakharov, Melnikov, Popov, Pletnev, Khudik and Punina2012) recorded Belemnitella? sp. from Late Campanian-Maastrichtian, likely mid-Maastrichtian deposits of the Magellan Rise in the Pacific Ocean.
Genus Neobelemnella Naidin, Reference Naidin, Menner, Moskvin, Naidin, Solovyev and Shimansky1975
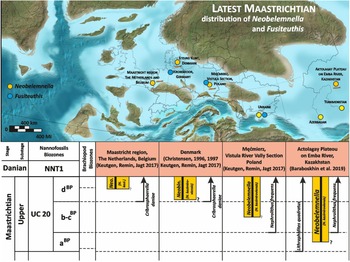
The genus Neobelemnella has recently been revised by Keutgen et al. (Reference Keutgen, Remin and Jagt2017) and Keutgen & Keutgen (Reference Keutgen and Keutgen2020) applying different methodological approaches. Neobelemnella kazimiroviensis (Skołozdrówna, Reference Skołozdrówna1932) and Neobelemnella skolozdrownae (Kongiel, Reference Kongiel1962) were recognised in both approaches. While the first species is widely recorded from Central Asia, Russia, Poland, Denmark, the Netherlands and Belgium, the second is limited to the uppermost Maastrichtian of Poland, Denmark, the Netherlands and Belgium (Fig. 6). Neobelemnella aff. kazimiroviensis (Skołozdrówna, Reference Skołozdrówna1932) was recorded only from Russia and Kazakhstan, while Neobelemnella pensaensis (Naidin, Reference Naidin1952) is difficult to separate from N. kazimiroviensis as transitory forms occur (Keutgen & Keutgen, Reference Keutgen and Keutgen2020).

Figure 6. Paleogeographic distribution of the representatives of Neobelemnella and Fusiteuthis and the diachronic appearance of Neobelemnella across Europe against the FAD of selected nannoplankton species. Used with permission of Colorado Plateau Geosystems Inc.Global Paleogeography and Tectonics in Deep Time © 2016.
Genus Fusiteuthis Kongiel, Reference Kongiel1962
Single representatives of this dubious genus with the single species Fusiteuthis polonica Kongiel, Reference Kongiel1962 have been reported from the uppermost Campanian to upper upper Maastrichtian in northwest Europe and the Crimea: from the lanceolata Zone of Kronsmoor (northern Germany), the obtusa Zone of Dziurków (Poland) and from the uppermost Maastrichtian of Crimea and Poland (Christensen, Reference Christensen2002; Remin, Reference Remin2010). Remin (Reference Remin2010) suggested that F. polonica could represent an intergeneric hybrid between the genera Belemnitella and Belemnella or Belemnitella and Neobelemnella since it possesses features characteristic for both study genera.
Typical belemnite species of the here mentioned genera are shown in Fig. 7, indicating the variation in size and shape of the genera.

Figure 7. A1-A2. Belemnella obtusa Schulz, Reference Schulz1979, KN 810 – holotype, Kronsmoor, Germany (Schulz, Reference Schulz1979; Remin, Reference Remin2012); B1-B2. Belemnella sumensis Jeletzky, Reference Jeletzky1949, HR014, Hrebenne, Poland (Remin, Reference Remin2018); C1-C2. Neobelemnella kazimiroviensis (Skołozdrówna, Reference Skołozdrówna1932), NHMM MK 2516, Maastricht, The Netherlands (Keutgen et al., Reference Keutgen, Remin and Jagt2017); D1-D2. Neobelemnella skolozdrownae (Kongiel, Reference Kongiel1962), MWGUW ZI/69/70, Nasiłów, Poland (Keutgen et al., Reference Keutgen, Remin and Jagt2017); E1. Fusiteuthis polonica Kongiel, Reference Kongiel1962, Mcd 162 - holotype, Nasiłów, Poland (Kongiel, Reference Kongiel1962); F1-F2. Belemnitella badlandsensis Landman, et al., Reference Landman, Remin, Garb and Chamberlain2013, AMNH 79950, holotype, Fox Hills Formation, AMNH loc. 3283, Badlands National Park, Pennington County, South Dakota (Landman et al., Reference Landman, Remin, Garb and Chamberlain2013); G1-G2. Belemnitella bulbosa Meek & Hayden, Reference Meek and Hayden1857, AMNH 79945, Fox Hills Formation, AMNH loc. 3283, Badlands National Park, Pennington County, South Dakota (Landman et al., Reference Landman, Remin, Garb and Chamberlain2013); H1-H2. Belemnitella/?Neobelemnella subfusiformis (Whitfield, Reference Whitfield1892), ANSP 19488, holotype, New Jersey (Jeletzky, Reference Jeletzky, Richards and Ramsdell1962); I1-I2. Belemnitella americana (Morton, Reference Morton1830), (ex Remin’s coll.), New Jersey; J1-J2. Belemniella posterior Kongiel, Reference Kongiel1962, Sad/szk/053, Vistula section, Poland (Remin, Reference Remin2015); K1-K2. Belemnitella langei Jeletzky, Reference Jeletzky1948, Sad/szk/010, Vistula section, Poland (Remin, Reference Remin2015). 1 – ventral view; 2 – view of the split guard showing internal features (in lateral position) or lateral view if not split.
Discussion
In the uppermost Campanian, the genus Belemnella appeared almost contemporaneously from Kazakhstan in the east to the United Kingdom in the west (Fig. 1) and rapidly began to evolve into local (sub)species, thereby replacing the genus Belemnitella. With respect to the genus Belemnitella, species diversity was unusually high with three species in the uppermost Campanian lanceolata Zone of the Middle Vistula River valley (Poland), which is in contrast to only one species of Belemnella (Remin, Reference Remin2012, Reference Remin2015). In the latest Campanian inflata Zone, the number of Belemnitella species remained similar and that of Belemnella rose to four, resulting in a very unusual belemnite species diversity of seven. However, the number of collected Belemnitella specimens (N = 22) is distinctly smaller (35%) than that of Belemnella (N = 40; Remin, Reference Remin2012, Reference Remin2015). The spread of Belemnella is associated with the negative excursion in δ13C at the Campanian-Maastrichtian boundary (Wilmsen et al., Reference Wilmsen, Engelke, Linnert, Mutterlose and Niebuhr2019), which is usually interpreted as indicative of a sea-level fall. An argument in favour of this interpretation represents the earliest Maastrichtian isotope minimum (eMim) dated by Wilmsen et al. (Reference Wilmsen, Engelke, Linnert, Mutterlose and Niebuhr2019) at 71.94 Myr, which corresponds well with the sea-level lowstand KMa1 of Haq (Reference Haq2014) at 72.0 Myr. Following Wiese’s et al. (Reference Wiese, Košták and Wood2009) argumentation, the sea-level fall progressively created conditions for the migration of Belemnella by the expansion of shelf settings with favourable depths for belemnite immigration. However, the origin of the genus Belemnella is still a mystery. Schulz (Reference Schulz1979) suggested a relationship with the genus Belemnellocamax Naidin, Reference Naidin1964b, the latest known representative of which being almost exclusively recorded from the Baltoscandian Subprovince. By contrast, Naidin (Reference Naidin and Krumgol’ts1974) supported an origin from eastern European species of Belemnitella. The appearance of Belemnella also coincided with a conspicuous latest Campanian cooling event, which has received considerable attention (e.g. Linnert et al., Reference Linnert, Robinson, Lees, Bown, Pérez-Rodríguez, Petrizzo, Falzoni, Littler, Arz and Russell2014, Reference Linnert, Engelke, Wilmsen and Mutterlose2016; Thibault et al., Reference Thibault, Anderskouv, Bjerager, Boldreel, Jelby, Stemmerik and Surlyk2015; Wilmsen & Niebuhr, Reference Wilmsen and Niebuhr2017) and suggests an immigration of Belemnella from a cooler northern or north-eastern region.
The increase in the number of Belemnella species within c. 800 kyr in the lanceolata and inflata zones may be interpreted as a typical example for adaptive radiation, a process where organisms diversify rapidly from an ancestral species into a multitude of new forms due to a change in the environment or the availability of new ecological niches making new resources available. This suggests first of all parapatric and peripatric speciation, both of which go hand in hand with adaptation to ecological niches. Allopatric speciation is also taken into consideration, for example, when populations become separated by a geographic barrier or simply by a large distance between them. If, however, a reproductive barrier is removed, reuniting two previously isolated populations, fertile hybrids may occur. As a consequence, the two populations may unite again, yet exhibiting a larger range of variation. Alternatively, hybrids may be of low fitness, reinforcing the separation of the two populations and resulting in speciation. In rare cases, new species can also be created through hybridization, followed by reproductive isolation, if the hybrids are favoured by natural selection. All of these different speciation processes could have led to the evolution of different ‘morphotypes’ such as distinguished in the Kronsmoor quarry ‘Saturn’ and the Middle Vistula River valley (Remin in Niebuhr et al., Reference Niebuhr, Hampton, Gallagher and Remin2011; Remin, Reference Remin2012).
In the Middle Vistula valley, the Campanian-Maastrichtian boundary was crossed by only two of seven species, one each belonging to Belemnitella and Belemnella, indicating the severe impact of a hypothetical ‘boundary event’ on the belemnite populations (Figs. 1, 3). From The Middle Vistula area, only a single species of the mucronata-group (Bt. posterior) is recorded. From Western Europe (United Kingdom, Netherlands, Belgium), a second species of this group is recorded (B. minor II), and a representative of the langei-group (Bt. pulchra) is known from the obtusa Zone of Kronsmoor in northern Germany and the Mons Basin, Belgium (Schulz, Reference Schulz1982; Christensen, Reference Christensen1999).
In the Middle Vistula section, this ‘boundary event’ corresponds to a characteristic lithologic unit, the ‘boundary marl’. Within the ‘boundary marl’ as well as directly below and above this level, belemnites are rare or absent (Remin, Reference Remin2012), altogether indicating that the ‘boundary marl’ seems thus to represent an interval in which an important event in the early evolutionary history of Belemnella and Belemnitella took place. At the same stratigraphic level in the ‘Saturn’ quarry (Kronsmoor) representatives of Belemnella are lacking, immediately below the Campanian-Maastrichtian boundary at mB609 (Niebuhr et al., Reference Niebuhr, Hampton, Gallagher and Remin2011). Both Bl. obtusa and Bl. vistulensis have their FAD somewhat above the Campanian-Maastrichtian boundary, while Bl. sp. A and Bl. sp. F successively appeared in Poland and northern Germany during the following c. 600 kyr, while Belemnitella became rare, still resulting in the co-existence of up to six belemnite species in the obtusa Zone compared to seven in the topmost Campanian of the Middle Vistula valley (Figs. 1, 3). The percentage of Belemnitella in the fauna comprises c. 7%.
The available data indicate that the ‘boundary marl’ event did not favour Belemnella over Belemnitella, but hit both, which manifests itself in the species composition and frequency. It remains to be tested to what extent the ‘boundary marl’ event was a supra-regional event, but it obviously affected belemnite evolution in northern Germany and Poland.
From the sumensis Zone of Poland in total three species of Belemnella and two of Belemnitella are recorded (Figs. 1, 3), whereby the first record of the second species Bt. junior is from a level high in the sumensis Zone at Boiska (Kongiel, Reference Kongiel1962). From the sumensis Zone at Hrebenne (southeast Poland), 27 specimens were studied, only a single (4%) belonging to Bt. pulchra.
In the type Maastricht region (the Netherlands, Belgium), the stratigraphically oldest Bt. junior is known from Altembroeck (northeast Belgium) from a level close to the base of unit 4 of the Vijlen Member (sumensis Zone), which was estimated c. 300 kyr older than the base of the Vijlen Member as exposed at Hallembaye quarry (Belgium, 50°44’54” N, 5°38’54” E), formerly known as Ciment Portland Liégeois (CPL), currently Kreco (Keutgen, Reference Keutgen2018). Vellekoop et al. (Reference Vellekoop, Kaskes, Sinnesael, Huygh, Déhais, Jagt, Speijer and Claeys2022, table 1) dated the base of the Vijlen Member at Hallembaye at c. 70.4 Myr, which implies for the base of the unit 4 of the Vijlen Member at Altembroeck a numerical age of c. 70.7 Myr. An alternative interpretation would take the dating of the ‘Zonneberg Horizon’ by Vellekoop et al. (Reference Vellekoop, Kaskes, Sinnesael, Huygh, Déhais, Jagt, Speijer and Claeys2022, table 1) as a reference, which would suggest a numerical age for the base of unit 4 of the Vijlen member of 70.3 Myr (= 69.9 + 0.4 Myr), assuming a sedimentation period for the units 4 and 5 of the Vijlen Member of 400 kyr (Keutgen, Reference Keutgen1998). Although these calculations allow only a rough estimate for the FAD of Bt. junior at Altembroeck between c. 70.3 and 70.7 Myr, correlation with the sumensis/tridens isotope minimum, dated at 71.1 Myr (Wilmsen et al., Reference Wilmsen, Engelke, Linnert, Mutterlose and Niebuhr2019), might be plausible when taking into account that deposition of unit 4 of the Vijlen Member occurred during a minor sea-level lowstand (Felder & Bless, Reference Felder and Bless1994). Thibault et al. (Reference Thibault, Harlou, Schovsbo, Schiøler, Minoletti, Galbrun, Lauridsen, Sheldon, Stemmerik and Surlyk2012, fig. 8) correlated the level of the sumensis/tridens isotope minimum with a position within the carbon-isotope events M1- (c. 70.2–71.0 Myr), which would imply a slightly younger age than that suggested by Wilmsen et al. (Reference Wilmsen, Engelke, Linnert, Mutterlose and Niebuhr2019). A rough correlation of the base of unit 4 of the Vijlen Member at Altembroeck with the sumensis/tridens isotope minimum at Kronsmoor is also supported by records of Bl. praearkhangelskii above this level at both localities (Schulz, Reference Schulz1979; Keutgen, Reference Keutgen1997). If the correlation of the appearance of Bl. praearkhangelskii in Altembroeck and Kronsmoor and that of the sumensis/tridens isotope minimum and the base of unit 4 of the Vijlen Member can be confirmed in future studies, this would imply that the appearance of both Bt. junior and Bl. praearkhangelskii would have been supported by a sea-level fall that created conditions favourable for the immigration of belemnites (Wiese et al., Reference Wiese, Košták and Wood2009). Noteworthy, the appearance of Bt. junior in the Maastricht area did not coincide with a temperature increase (Vonhof et al., Reference Vonhof, Jagt, Immenhauser, Smit, Van den Berg, Saher, Keutgen, Reijmer, Jagt, Jagt-Yazykova and Schins2011; Keutgen, Reference Keutgen2018).
The stratigraphically oldest record of Bt. junior in the Maastricht area is conservatively dated at c. 70.3 Myr. It suggests a concurrent range of Bl. sumensis and Bt. junior in the Maastricht region of c. 250 kyr (Keutgen, Reference Keutgen2018). The disappearance of the genus Belemnella in the Maastricht region thus corresponds to a position in the ‘upper’ sumensis Zone in northern Germany, indicating that the genus Belemnella disappeared as much as c. 1.6 Myr earlier in the type Maastricht area than in northern Germany (Thibault et al., Reference Thibault, Harlou, Schovsbo, Schiøler, Minoletti, Galbrun, Lauridsen, Sheldon, Stemmerik and Surlyk2012).
At Boiska, Poland, Bt. junior and Bl. sumensis co-occur as well, however, at a stratigraphically higher level as indicated by the presence of L. quadratus (Dubicka & Peryt, Reference Dubicka and Peryt2012). Seemingly, the situation at the Aktolagai plateau section (middle course of the Emba River, Western Kazakhstan) around the FAD of L. quadratus resembles that at Boiska. Baraboshkin et al. (Reference Baraboshkin, Benyamovskiy, Guzhikov, Aleksandrova, Pervushov, Celler, Ovechkina, Kalyakin, Kopaevich, Guzhikova, Pokrovskiy, Baraboshkin, Yakovishina, Zholtaev, Ali-Zade, Degtyarev, Ergaliev, Kilipko, Sakiev, Seitmuratova, Zholtaev, Ali-Zade, Degtyarev, Ergaliev, Kilipko, Sakiev, Seitmuratova, Zhaymina, Fazylov, Bekenova and Musina2019, fig. 6) reported ?Bt. junior co-occurring with Belemnella, probably with Bl. sumensis. Already Naidin (Reference Naidin1973) reported on the co-occurrence of Bt. junior and Bl. sumensis from the northern part of the Donbass region. By contrast, Bt. junior is not known from the northeastern parts of the Russian Platform, for example, from the Volga region (Naidin, Reference Naidin1973).
In the lower upper Maastrichtian at least in Western and Central Europe, the number of belemnite species is further reduced to two, both belonging to Belemnitella (Christensen et al., Reference Christensen, Schmid and Schulz2004; Remin in Dubicka & Peryt, Reference Dubicka and Peryt2011). The cause of the extinction of Belemnella in these areas is unclear, but might be linked to the onset of the early Maastrichtian warming. Vonhof et al. (Reference Vonhof, Jagt, Immenhauser, Smit, Van den Berg, Saher, Keutgen, Reijmer, Jagt, Jagt-Yazykova and Schins2011, table 1) recorded oxygen isotope data (δ18O) from the type Maastrichtian area (Netherlands) measured almost exclusively at belemnite guards with the metre scale of the collected belemnites set at 0 at the Zonneberg Horizon, which was interpreted by the late P.J. Felder as the base of unit 6 of the Vijlen Member (Keutgen, Reference Keutgen2018). It follows from their data that δ18O-values remained relatively stable until close to the top of unit 5 of the Vijlen Member and then decreased to a lower level that was reached close to the boundary between the Lixhe 1 and Lixhe 2 Members. Thus, temperature started to rise in the Maastricht region approximately at c. 70.0–70.4 Myr, and Belemnella disappeared c. 50–100 kyr earlier, somewhat above the base of unit 5 of the Vijlen Member – stratigraphically younger records of Belemnella are considered reworked (Keutgen, Reference Keutgen2018). This would favour a close relationship between temperature rise and demise of Belemnella. By contrast, the disappearance of Belemnella in northern Germany (Hemmoor quarry) is dated c. 68.4 Myr (Schulz, Reference Schulz1979; Thibault et al., Reference Thibault, Harlou, Schovsbo, Schiøler, Minoletti, Galbrun, Lauridsen, Sheldon, Stemmerik and Surlyk2012), but for the Stevns-1 borehole (Denmark), which may be best comparable to Hemmoor, the temperature increase appeared at c. 69.5 Myr (Thibault et al., Reference Thibault, Harlou, Schovsbo, Stemmerik and Surlyk2016), c. 1.1 Myr earlier than the demise of Belemnella. Different to the belemnite-derived δ18O-values from the Maastricht region, those from the Stevns-1 borehole represent bulk samples. While the measurements of δ18O-values from belemnites reflect the temperature, where the belemnites lived (Zakharov et al., Reference Zakharov, Tanabe, Safronov and Smyshlyaeva2014), bulk samples relate to the sea-surface temperature (Thibault et al., Reference Thibault, Harlou, Schovsbo, Stemmerik and Surlyk2016) and may not reflect the temperature conditions of the preferred biotope of Belemnella. Wilmsen & Niebuhr (Reference Wilmsen and Niebuhr2017) suggested a nektobentic mode of life for belemnites and Keutgen et al. (Reference Keutgen, Remin and Jagt2017) assumed that small (juvenile) belemnites might have preferred a shallower-marine habitat, while adults ventured out into open and deeper water. Hoffmann and Stevens (Reference Hoffmann and Stevens2020) pointed out that long lateral or vertical migration of belemnites of the Campanian genera Gonioteuthis Bayle, Reference Bayle1878 and Belemnitella d’Orbigny, Reference Orbigny1840 are less probable, as indicated by the presence of nearly all ontogenetic growth stages in populations collected from the marly limestones and calcareous marls exposed in the Höver quarry (northern Germany). These sediments were deposited at depths less than that of the typical Chalk facies but distinctly below storm-wave base (Christensen, Reference Christensen2000), with a depth of c. 70–110 m calculated from the data of Wilmsen & Niebuhr (Reference Wilmsen and Niebuhr2017). It is suggested that older belemnites could have escaped the warmer sea-surface temperatures recorded by Thibault et al. (Reference Thibault, Harlou, Schovsbo, Stemmerik and Surlyk2016) at the bottom of the mid Maastrichtian northern German shelf sea with a water depth of 100–150 m (Wilmsen & Niebuhr, Reference Wilmsen and Niebuhr2017), being deeper than the Vijlen Chalk Member, for example, at Hallembaye with a water depth of c. 80 m (Jagt & Jagt-Yazykova, Reference Jagt, Jagt-Yazykova, Jagt, Donovan and Jagt-Yazykova2012). Assuming a temperature gradient of 12.5–18.75 m/1°C, Wilmsen & Niebuhr (Reference Wilmsen and Niebuhr2017) would explain a by c. 2.0–3.5°C lower temperature at the bottom of the Hemmoor Chalk Sea. However, it may be hypothesised that juvenile representatives of Belemnella may have been exposed to higher temperatures as a consequence of their preferred shallow marine habitat. It is assumed that either they successfully adapted to warmer temperature or that their spawning grounds were situated in colder regions, presumably towards the north.
With the appearance of the genus Neobelemnella in the upper upper Maastrichtian, species diversity increased and the number of species rose to up to five in the Middle Vistula valley – three belonging to Neobelemnella (Keutgen & Keutgen, Reference Keutgen and Keutgen2020) and two of Belemnitella (Kongiel, Reference Kongiel1962; Christensen et al., Reference Christensen, Schmid and Schulz2004), the latter known only from sporadic records. Rare finds of Fusiteuthis polonica are not considered.
The stratigraphically oldest records of N. kazimiroviensis are from Kazakhstan (Aktolagay Plateau) from the calcareous nannofossil zone UC20a, above the FAD of calcareous nannofossil L. quadratus, but below the FAD of N. frequens (Baraboshkin et al., Reference Baraboshkin, Benyamovskiy, Guzhikov, Aleksandrova, Pervushov, Celler, Ovechkina, Kalyakin, Kopaevich, Guzhikova, Pokrovskiy, Baraboshkin, Yakovishina, Zholtaev, Ali-Zade, Degtyarev, Ergaliev, Kilipko, Sakiev, Seitmuratova, Zholtaev, Ali-Zade, Degtyarev, Ergaliev, Kilipko, Sakiev, Seitmuratova, Zhaymina, Fazylov, Bekenova and Musina2019). In Poland, the stratigraphically oldest representatives are from Mięćmierz, which is situated above the FAD of N. frequens (Dubicka & Peryt, Reference Dubicka and Peryt2012) and, hence, they are considered stratigraphically younger than those from Kazakhstan (Fig. 6). First records of N. kazimiroviensis from Denmark are even younger (Cribrosphaerella daniae calcareous nannofossil zone) than those from Poland (Machalski, Reference Machalski1996), and specimens from the Maastricht region (the Netherlands, Belgium) appear even later (c. 50–100 kyr before the K-Pg boundary), altogether implying a westwards migration of this species during the upper Maastrichtian.
Acknowledgements
The authors gratefully acknowledge the suggestions of I. Walaszczyk and two reviewers that were incorporated into the study. This research was funded by the Austrian Science Fund (FWF), grant number P36201-B, Grant-DOI 10.55776/P36201 (N.K) and by the Polish National Science Centre (Narodowe Centrum Nauki), grant number – UMO-2018/29/B/ST10/02947 (Z.R). For the purpose of Open Access, the authors have applied a CC BY public copyright licence to any Author Accepted Manuscript (AAM) version arising from the submission. The authors do not have any conflicts of competing interest. The review was been written following the Cambridge’s Ethical Guidelines.



 Open access
Open access