


Staphylococcus aureus is one of the major contagious udder pathogens and causes great economic losses in the dairy industry by way of compromising milk quality, decreasing milk production and causing recurrent infections (Wang et al., Reference Wang, Zhang, Zhou, He, Yong, Shen, Szenci and Han2016; Gao et al., Reference Gao, Barkema, Zhang, Liu, Deng, Cai, Shan, Zhang, Zou, Kastelic and Han2017). Antimicrobial agents have been used to treat bovine mastitis of bacterial origin worldwide. However, antimicrobial resistance (AMR) in bacteria challenges efficacy, making treatment costly, time consuming and increasing the risk of therapeutic failure (Sharma et al., Reference Sharma, Rokana, Chandra, Singh, Gulhane, Gill, Ray, Puniya and Panwar2017). Indeed, AMR can disseminate without any geographical boundaries and affect diverse animal populations including humans (Littmann and Viens, Reference Littmann and Viens2015). It has been widely recognized as one of the most serious public health challenges of the twenty-first century (Hernando-Amado et al., Reference Hernando-Amado, Coque, Baquero and Martínez2019). It is, therefore, important to survey and study the occurrence of AMR in pathogenic flora of dairy cows, to understand the possible mechanism of AMR and to develop alternative strategies and detect novel target sites for combating AMR.
Bacteria can develop AMR either vertically (gene mutation) or horizontally (obtaining new genes from other species or strains: Martinez and Baquero, Reference Martinez and Baquero2000; Palmer et al. Reference Palmer, Veronica and Gilmore2010). It has been reported that integrons play a key role in dissemination of AMR through horizontal gene transfer, particularly in Gram-negative pathogens (Xia et al., Reference Xia, Xu, Qin, Li, Liu, Kang, Gu and Ma2016). Nandi et al. (Reference Nandi, Maurer, Hofacre and Summers2004) reported that in poultry litter, the major reservoir for class 1 integrons is Gram-positive bacteria instead of Gram-negative Enterobacteriaceae. For S. aureus, presence of class 1 integrons was reported in samples from humans (Deng et al., Reference Deng, Bao, Ji, Chen, Liu, Miao, Chen, Bian, Li and Yu2015a), however, studies on presence of integrons in S. aureus isolates obtained from dairy cows are still scarce. Therefore, the objectives of this study were to determine the phenotypic and genotypic AMR as well as the occurrence of class 1 integrons in S. aureus strains isolated from bovine clinical mastitis in China, and to investigate the associations between class 1 integrons and AMR genotypes and phenotypes.
Materials and methods
The study was conducted in compliance with guidelines of the Beijing Municipality on the Review of Welfare and Ethics of Laboratory Animals, approved by the Beijing Municipality Administration Office of Laboratory Animals (BAOLA) and approved by the China Agricultural University Animal Ethics Committee (protocol CAU-AEC-2010-0603).
Herds and sampling
A total of 103 S. aureus isolates were collected over a period of four years (2013–2016) from quarter milk samples of Holstein cows with clinical mastitis in 19 large (> 500 cows) dairy herds in 9 provinces which are the major producers of milk in China (online Supplementary Fig. S1). Briefly, 1021 milk samples were collected following NMC guidelines (2017) from lactating cows (one sample per cow) suffering from clinical mastitis (NMC, 2017). Clinical mastitis was diagnosed based on visual abnormality /inflammation of the udder or its milk secretion. Samples were collected in sterile bottles (50 mL) and transported on ice to the mastitis laboratory at the China Agricultural University (Beijing, China).
Isolation and identification of S. aureus
Milk samples (10 μl out of 50 ml) were spread-plated on TSA (trypticase soya agar; Sigma, India) supplemented with 5% defibrinated sheep blood and incubated at 37 ℃ for 16–24 h. S. aureus isolates were identified by colony morphology, hemolysis, Gram staining, catalase test, tube coagulase test, oxidation and fermentation of mannitol salt agar (AOBOX, Beijing, China), as well as 16S rRNA, coa and nuc gene sequence analysis, as described by Wang et al. (Reference Wang, Zhang, Zhou, He, Yong, Shen, Szenci and Han2016).
Antimicrobial susceptibiltiy testing
To analyze the antimicrobial susceptibility profiles of all isolates, minimum inhibitory concentrations (MICs) were determined against a panel of 11 different antimicrobials using the broth microdilution method following the Clinical Laboratory and Standard Institutes (CLSI) guidelines (CLSI, 2018). Antimicrobial agents were selected either according to their availability in commercial products or working as representative of an antimicrobial family: penicillin, ampicllin, oxacillin, erythromycin, tetracycline, ciprofloxacin, ceftriaxone, cefotaxime, gentamicin, clindamycin. A concentration of 0.125–64 μg/ml was used for S. aureus. Staphylococcus aureus ATCC 29213 was used as the quality control strain. Breakpoints were defined as described by the CLSI (CLSI, 2018). Isolates were classified as susceptible, intermediate or resistant to each antimicrobial. Multidrug resistance was defined as resistant to at least 3 antimicrobials. As defined by CLSI, MRSAs (methicillin resistant S. aureus) are those strains that possess mecA gene. Frequency distribution of MIC50 and MIC90 were calculated for each antimicrobial.
Presence of antimicrobial resistance genes
Template DNA of the 103 S. aureus isolates were prepared using bacterial genomic DNA extraction kit (CW, Beijing, China) according to the manufacturer instructions (Zhang et al., Reference Zhang, Gao, Barkema, Ali, Liu, Deng, Naushad, Kastelic and Han2018). The 13 acquired resistance genes were analyzed using PCR (primers identified in online Supplementary Table S1).
Detection of three classes of intergrons
Presence of class 1, 2 and 3 integrons was analyzed in all isolates using PCR as per Goldstein et al. (Reference Goldstein, Lee, Sanchez, Hudson, Phillips, Register, Grady, Liebert, Summers, White and Maurer2001). The following program was employed: 5 min of initial denaturation at 94 ℃, followed by 30 cycles (30 s of denaturation at 94 ℃, 30 s of annealing at 55 ℃ and 1 min of extension at 72 ℃) and a final extension step. The PCR products were randomly selected for sequencing to ensure specificity and accuracy.
Gene cassettes inserted in the variable regions of class 1 integrons were amplified as described by Lévesque et al. (Reference Lévesque, Piche, Larose and Roy1995). To determine whether each cassette gene PCR amplicon with the same size had the same content, PCR products were digested with AluI and RsaI (New England Biolabs, Beijing, China). PCR products with the same restriction fragment length polymorphism (RFLP) pattern were considered to contain the same gene cassettes (Lévesque et al., Reference Lévesque, Piche, Larose and Roy1995; Li et al., 2015).
Statistical analysis
SPSS version 22.0 (IBM, Chicago, IL, USA) was used for statistical analysis. Index hierarchical cluster analysis was performed using between groups linkage method with measure of binary squared Euclidean distance to explore the similarity among antimicrobial resistance phenotypes, resistances genes and int1 (Zhang et al., Reference Zhang, Meng, Chen, Tu and Zhang2017). Also, the association between phenotypic and genotypic resistance was explored using χ2 or Fisher's exact test on contingency tables (positive and negative for phenotype or genotype of isolates).
Results
Phenotypic antimicrobial susceptibility
Overall, for antimicrobials tested, MIC50 ranged from 0.25 to 4 μg/ml and MIC90 from 0.5 to 64 μg/ml (Table 1). Susceptibility for single antimicrobial agents ranged from 0% (for erythromycin) to 96% (for gentamicin). All S. aureus isolates demonstrated a low sensitivity to ampicllin (12%) and penicillin (16%). Most isolates (64%) were of intermediate susceptibility for erythromycin, followed by ceftriaxone (22%), clindamycin (4%), cefotaxime (2%), cefotaxime (1%) and ciprofloxacin (1%). In total, 84% of S. aureus isolates were susceptible to oxacillin (Table 2).
Table 1. Minimal inhibitory concentration (MIC) distribution for the 103 Staphylococcus aureus isolates
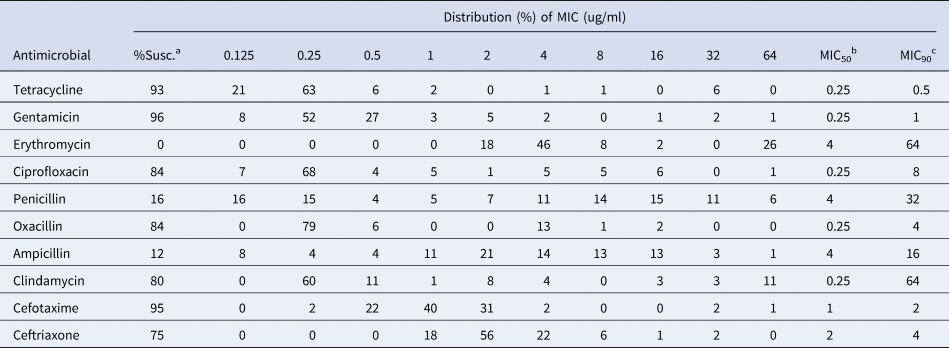
a Percent of susceptible isolates according to CLSI (2018).
b The MIC value that inhibits growth of 50% of the isolates.
c The MIC value that inhibits growth of 90% of the isolates.
indicates the CLSI epidemiological cut off values.
Table 2. Antimicrobial activity and potency of 11 antimicrobials against MRSA and MSSA isolated from clinical mastitis in Chinese dairy herds
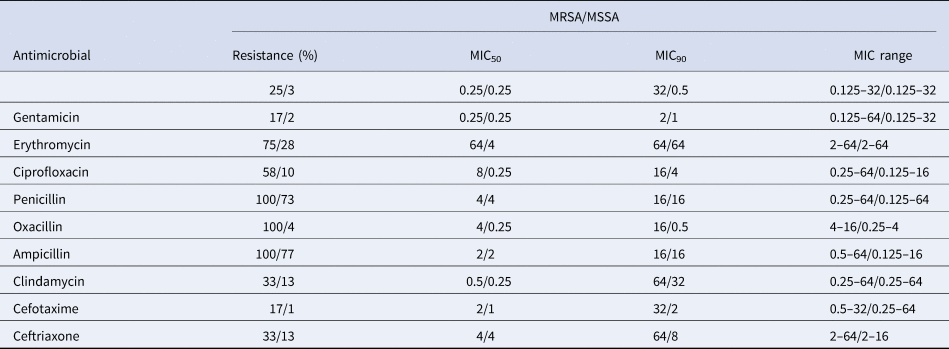
A total of 12 isolates (12%) were identified as MRSA (methicillin-resistant Staphylococcus aureus, mecA gene positive), which were distributed in 8 regions except Inner Mongolia. Compared to MSSA (methicillin-sensitive Staphylococcus aureus, mecA gene negative) isolates, MRSA isolates demonstrated higher resistance rate to all the 10 antimicrobial agents. In addition, the MIC ranges of gentamicin, ciprofloxacin, penicillin, ampicllin, oxacillin, cefotaxime and ceftriaxone were larger in MRSA isolates. Similarly, the MIC50 results of MRSA isolates were higher than MSSA for all antimicrobial agents except for tetracycline, gentamicin, penicillin, ampicllin and ceftriaxone; MIC90 values of MRSA isolates were also higher than MSSA isolates except for penicillin and ampicllin (Table 3). In addition, the multidrug resistance phenotype was much more frequent in MRSA isolates (12, 100%) than in MSSA isolates (51, 56%: Fig. 1). MSSA isolates demonstrated different resistance patterns: 24 MSSA isolates (47%) showed resistance to 3 antimicrobials, 15 isolates (10%) to 5 antimicrobials and 10 isolates to 4 antimicrobials. While for MRSA isolates, resistance to 4, 5, 6, 9 and 10 antimicrobials were identified with 4, 4, 2,1 and 1 isolates, respectively.
Table 3. The MIC50/90 of Staphylococcus aureus isolates with or without a particular antimicrobial resistance gene and association between resistance phenotype and genotype
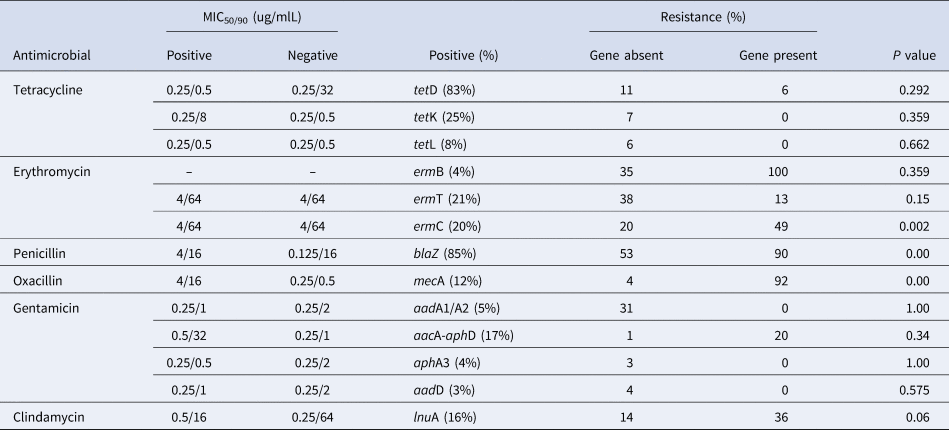
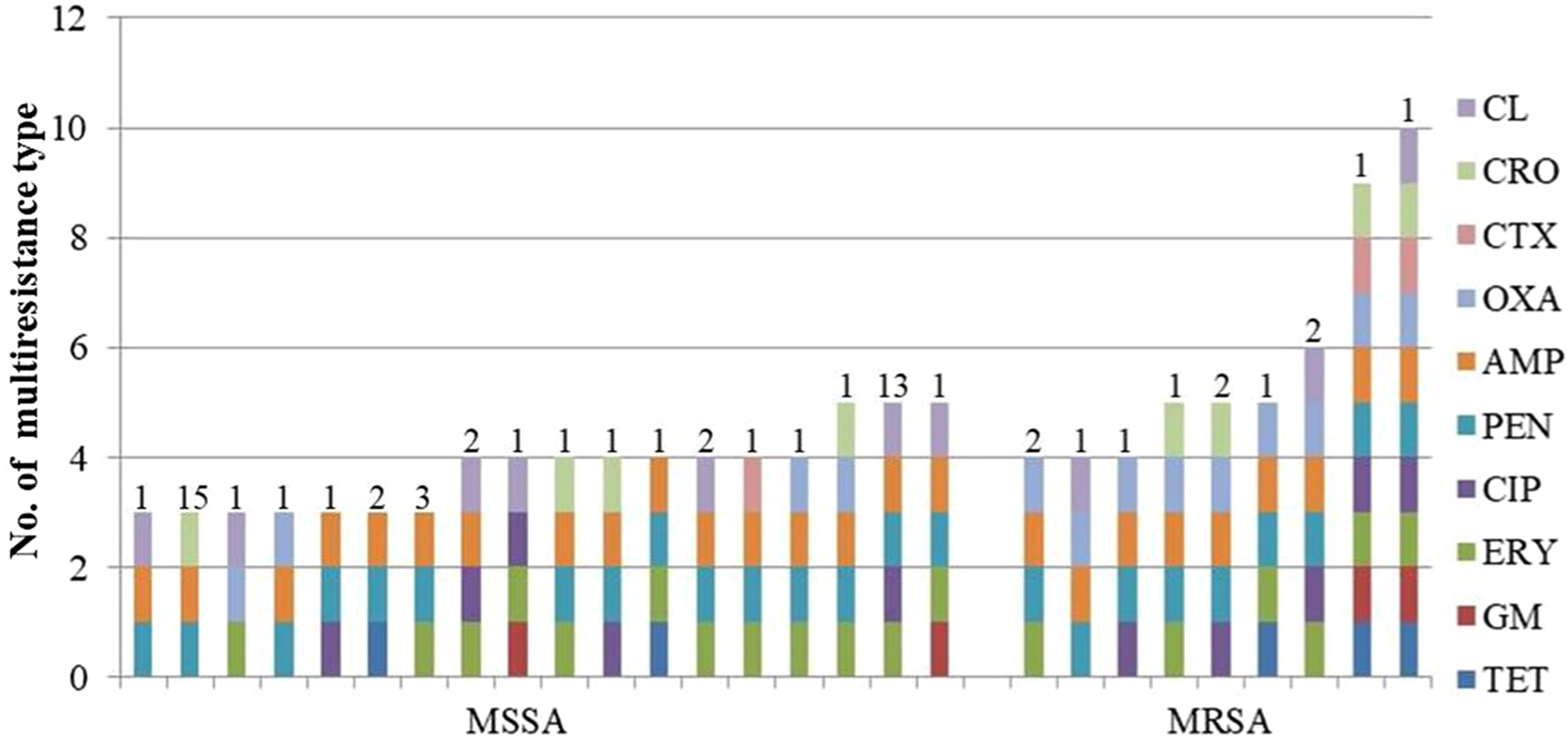
Figure 1. Multi-resistance patterns of MRSA and MSSA isolates from clinical mastitis in China. Different numbers above bars indicate the number of isolates corresponding to the resistance pattern.
Occurrence of resistance genes
Of the 13 resistance genes, blaZ was detected most frequently (85%), followed by tetD (83%). tetK, ermT and ermC were also detected in considerable proportion (Table 3). Furthermore, we analyzed the frequency of drug-specific resistance on presence or absence of a particular antimicrobial resistance gene, results indicated that presence of some resistance genes (ermC, blaZ and mecA) was associated with drug-specific resistance phenotype (Table 3).
Gene patterns were commonly observed for resistant isolates included the presence of single tetL (17% of tetracycline-resistant isolates), tetD-tetK (33% of tetracycline-resistant isolates), tetD-tetL (17% of tetracycline-resistant isolates), ermT (24% of erythromycin-resistant isolates), ermC (27% of erythromycin-resistant isolates), ermB (5% of erythromycin-resistant isolates) and ermT-ermC (3% of erythromycin-resistant isolates) (Fig. 2).
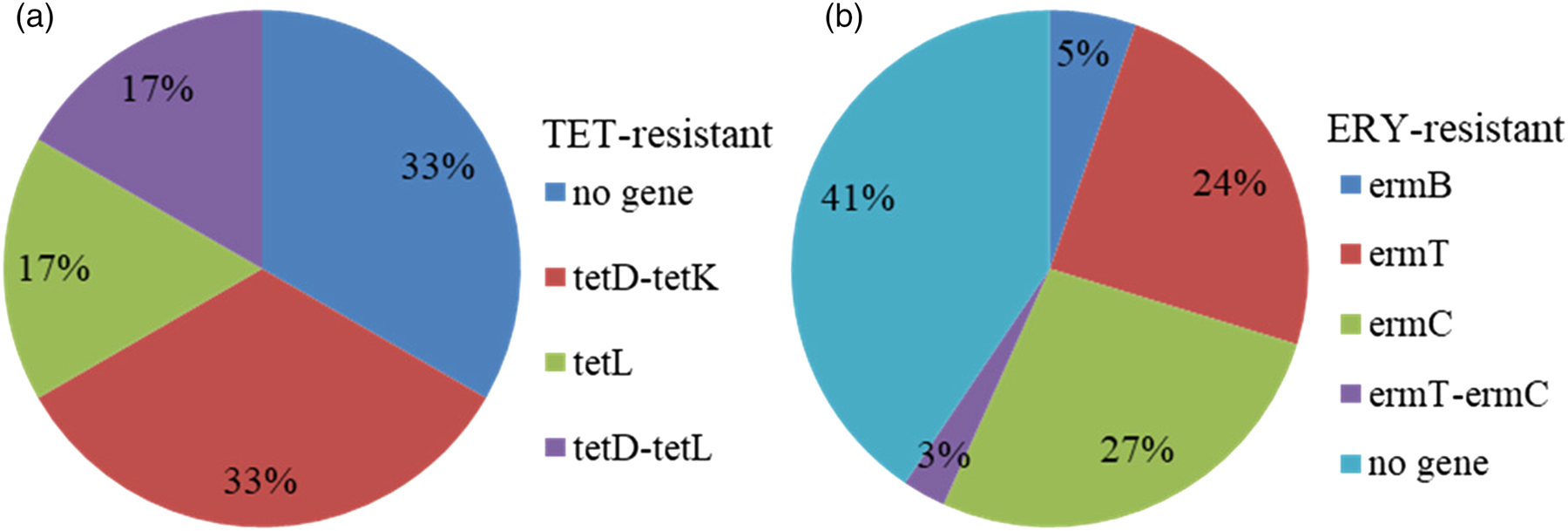
Figure 2. Gene patterns in Staphylococcus aureus from clinical mastitis resistant to antimicrobials of TET (a) and ERY (b).
Class 1 integrons
Of the 103 isolates, 84 (83%) were positive for intI1. From 76 of 84 isolates (89%), the class 1 integrons variable regions were amplified by PCR, yielding amplicons of 3 distinct sizes: 1600 bp (74 isolates), 1000 bp (1 isolate) and 750 bp (1 isolate). Digested with AluI and RsaI, a total of 5 different patterns were detected (Fig. 3).
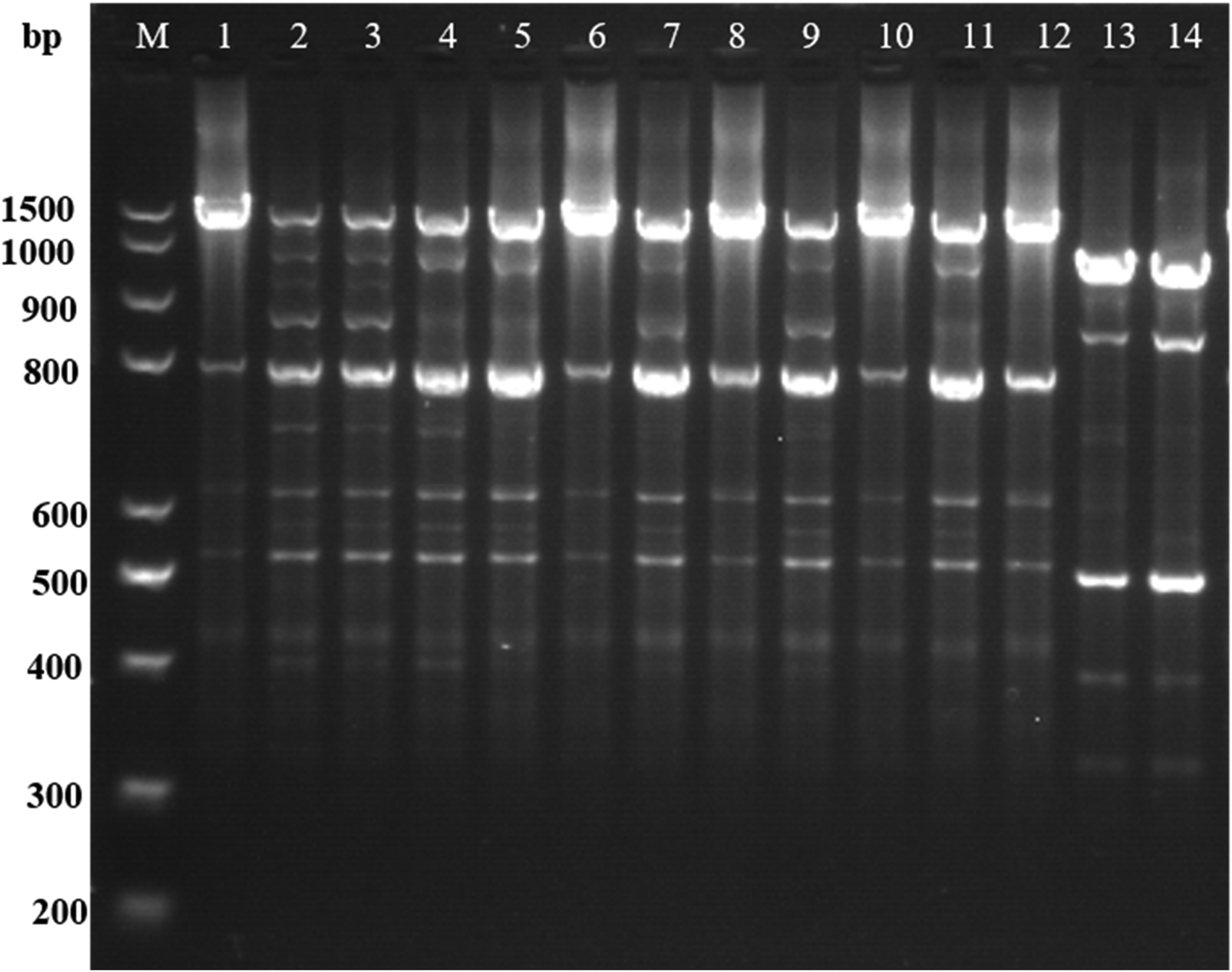
Figure 3. Representative profiles identified among Staphylococcus aureus from clinical mastitis by AluI and RsaI digestion in class 1 integrons gene cassette.
M, DNA ladder; 1, SA1 (Pattern 1); 2, SA2 (Pattern 2); 3, SA3 (Pattern 2); 4, SA4 (Pattern 3); 5, SA5 (Pattern 4); 6, SA6 (Pattern 1); 7, SA7 (Pattern 4); 8, SA8 (Pattern 1); 9, SA9 (Pattern 4); 10, SA10 (Pattern 1); 11, SA11 (Pattern 3); 12, SA12 (Pattern 5); 13, SA13 (Pattern 5); 14, SA14 (Pattern 5).
Associations among resistance genes, AMR and intI.
Index cluster analysis indicated that GM-resistance, CTX-resistance, TET-resistance, CIP-resistance, OX-resistance, CL-resistance, CRO-resistance and ERY-resistance in S. aureus isolates were highly associated (Fig. 4a). Resistance to AMP and PEN was associated with intI1. Correspondingly, aminoglycosides resistance genes (aphA3, aadA1/A2), macrolides resistance genes (ermB and ermT), tetracycline resistance genes (tetL and tetK), β-lactams resistance gene mecA and lincosamides resistance gene lunA were also highly associated. A similar association was present for intI1 and blaZ (Fig. 4b).

Figure 4. Index cluster analysis of Staphylococcus aureus from clinical mastitis in antimicrobial resistance phenotypes (a), resistances genes and mobile genetic element intI1 (b). When branch point was less than 5, these branches were determined as related to each other significantly.
Discussion
Despite considerable progress in alternative treatment strategies such as vaccines, AMR remains a major problem in the treatment of bovine mastitis, and poses a serious threat to public health (Wang et al., Reference Wang, Zhang, Zhou, He, Yong, Shen, Szenci and Han2016; Cheng et al., Reference Cheng, Qu, Barkema, Nobrega, Gao, Liu, De Buck, Kastelic, Sun and Han2019). In this study, AMR of S. aureus was exhibited against antimicrobials frequently used in dairy herds, such as erythromycin, penicillin, and ampicillin. Some AMR genes were involved in drug-specific AMR and vice versa. Class 1 integrons were associated with resistance to several drugs (penicillin and ampicillin) and antimicrobial resistance genes blaZ and tetD. Of note, the MIC90 of S. aureus against erythromycin was very high. Despite the lack of availability of an erythromycin clinical breakpoint for S. aureus isolated from bovine mastitis, concern should be raised for this, since it suggests that macrolides such as erythromycin might not be effective for treatment of mastitis caused by S. aureus in China. Furthermore, the MIC50/MIC90 (4/64) of erythromycin was higher than in both MSSA (< 0.25/4) and MRSA (> 4/> 4) isolates from human hospitals in previous report (Pfaller et al., Reference Pfaller, Sader, Flamm, Castanheira and Mendes2018). Similar results were present for CL (0.25/64 vs. < 0.25/2), which indicates that erythromycin and clindamycin resistance in S. aureus is developing regardless of bacterial origin. We observed a lower resistance rate to tetracycline (6 vs. 19%), ciprofloxacin (16 vs. 53%), ampicillin (88 vs. 91%) and penicillin (83 vs. 91%) compared to another study in the mid-east of China, in which 200 CM milk samples were included (Zhang et al., Reference Zhang, Li, Bao, Wei, Zhou, Zhang and Wang2016). Regarding the comparison between different studies, differences in resistance prevalence were observed among countries (Wang et al., Reference Wang, Zhang, Zhou, He, Yong, Shen, Szenci and Han2016; Beyene et al., Reference Beyene, Hayishe, Gizaw, Beyi, Abunna, Mammo, Ayana, Waktole and Abdi2017; Freitas et al., Reference Freitas, Mendes, Villarreal, Santos, Gonçalves, Gonzales and Nascente2018). We inferred these differences may be related to the source of samples, geographical distribution and bacterial genotype. The high and intermediate resistance against erythromycin (65%) and ceftriaxone (20%) indicated that veterinarians need to be more prudent when considering these antimicrobials.
MRSA has been reported in livestock in recent years all over the world with different prevalence. In our study, 12% of isolates harbored the mecA gene, much higher than the 0.05% reported by Saini et al. (Reference Saini, McClure, Léger, Keefe, Scholl, Morck and Barkema2012) in Canada, and the 1% by Ahangari et al. (Reference Ahangari, Ghorbanpoor, Shapouri, Gharibi and Ghazvini2017) in Iran. In total, the prevalence of MRSA strains reported from 7% to 19% in China was much higher than that in other countries, perhaps resulting from a misusage of various antibiotics (Zhang et al., Reference Zhang, Li, Bao, Wei, Zhou, Zhang and Wang2016; Li et al., Reference Li, Lu, Wang, Gao, Dai, Shang and Li2017; Qu et al., Reference Qu, Zhao, Nobrega, Cobo, Han, Zhao, Li, Li, Barkema and Gao2019). Compared with MSSA, MRSA in this study showed larger MIC range, higher MIC50/90 values for most antimicrobials and more multi-drug resistance, which was consistent with the previous study (Wang et al., Reference Wang, Zhang, Zhou, He, Yong, Shen, Szenci and Han2016).
In this study, blaZ gene was widespread in the herds, which was consistent with the prominent resistance to penicillin. However, the presence of other AMR genes did not always correspond with the resistance phenotype. For instance, tetD was the most prevalent gene whereas clinical resistance to tetracycline was uncommon. In addition, an association was observed between detection of gene ermC, blaZ and mecA encoding resistance to erythromycin, penicillin and oxacillin and their resistance phenotype. The discordance between phenotypic and genotypic resistance could be due to lack of expression of resistance genes or MIC increases below the adopted thresholds.
Integrons are considered an important source of AMR genes, which facilitate the horizontal spread of resistance genes within microbial populations (Deng et al., Reference Deng, Liu, Peters, Chen, Miao, Li, Li, Chen, Yu, Xu and Shirtliff2015b). Class 1 integrons were detected in 83% of isolates, lower than another study conducted in northwestern China, in which all 121 S. aureus isolates from clinical mastitis samples were positive for the intI1 gene (Li et al., 2018). The proportion of class 1 integrons in S. aureus isolates can be different among farm animals, hospitals and environment. For example, they were present in 73% of wounds, blood, urines, nasal and throat S. aureus isolates from hospitals in Tehran (Mostafa et al., Reference Mostafa, Siadat, Shahcheraghi, Vaziri, Japoni-Nejad, Vand Yousefi, Rajaei, Harifi Mood, Ebrahim Zadeh, Moshiri, Seyed Siamdoust and Rahbar2015), absent from all the MRSA isolates from a Chinese municipal wastewater treatment plant and a swine slaughterhouse wastewater treatment plant but present in all of the other wastewater samples tested (Wan and Chou, Reference Wan and Chou2015). High occurrence of intI1 gene suggests that S. aureus isolated from bovine mastitis can acquire and easily disseminate resistance genes.
Amplification and restriction digest of the variable region of class 1 integrons from 76 isolates allowed clustering into 5 types. The reasons why some intI1-positive isolates did not harbor gene cassettes may be that mutations happened at the 3′CS; or the variable region was too long to be amplified. Many studies reported different gene cassette in intI1-positive S. aureus isolates, including aadA2 (59%), dfrA12-orfF-aadA2 (49%), aacA4-cmlA1 (3%), dfrA17-aadA5 (1%), dfrA1-aadA1(45%), and aadA1(26%) (Xia et al., Reference Xia, Xu, Qin, Li, Liu, Kang, Gu and Ma2016; Li et al., 2018). Sometimes, the gene cassettes in class 1 integrons did not account for the total resistance phenotypes for erythromycin, tetracycline and penicillin (Li et al., 2018). We did not sequence the gene cassettes in this study but used index cluster analysis to estimate the association between intI1 and antimicrobial resistance phenotype and genotype. Results showed that, consistent with the penicillin resistance phenotype, incidence of int1 was also related to blaz gene. Similarly, a strong association was shown between int1 and penicillin-resistant phenotypes in E. coli isolated from swine (Zhang et al., Reference Zhang, Li, Wang, Qi, Wang, Xu, Liu, Zhang and Guo2020). This association suggests that these resistance mechanisms are co-selected, and warrants further investigations on how these determinants disseminating in dairy herds.
In conclusion, a high occurrence of AMR against erythromycin, ampicillin and penicillin in S. aureus isolated from clinical bovine mastitis in large dairy farms was detected, and the most frequent resistance genes were blaZ and tetD genes. Twelve percent of S. aureus isolates were mecA-positive. In addition, class 1 integrons were commonly detected in S. aureus and associated with penicillin resistance. This suggests that Class 1 integrons and penicillin resistance are genetically linked in S. aureus isolated from bovine mastitis in China.
Supplementary material
The supplementary material for this article can be found at https://doi.org/10.1017/S0022029924000566
Acknowledgments
This research was financially supported by the Basic Research Project-Youth Project (202001AU070098); National Natural Science Foundation of China (31902335); Yunnan Fundamental Research Projects (grant no. 202301AS070081), Rural Vitalization Science and technology Project -Gejiu Cattle Industry Science and Technology Mission in Yunnan Province (grant no. 202304BI090007).









