


Introduction
Several characteristics of the zebrafish (Danio rerio) make it an ideal model for studying development and disease. The high homology between zebrafish and humans, along with the ease with which the zebrafish genome can be modified, allows for development and assessment of disease models that generally mimic human disease (Goldsmith & Jobin, Reference Goldsmith and Jobin2012). In addition, zebrafish are easy to breed, mature quickly, produce large numbers of offspring, and are relatively inexpensive to maintain, making them an attractive option for many researchers (Pickart & Klee, Reference Pickart and Klee2014). Perhaps the most unique advantage to the zebrafish is its external development, which is highly conducive for studies of ocular development (Link & Collery, Reference Link and Collery2015). Of particular importance to studies of vision, the cone-rich retinas and diurnal vision of zebrafish complement the studies normally conducted in nocturnal, rod-dominant rodents (Allison et al., Reference Allison, Haimberger, Hawryshyn and Temple2004; Chhetri et al., Reference Chhetri, Jacobson and Gueven2014).
Traditional confocal microscopy can be used with fluorescent markers and transgenic zebrafish lines to examine retinal structure and development both ex vivo (Li et al., Reference Li, Tsujimura, Kawamura and Dowling2012; Fraser et al., Reference Fraser, Duval, Wang and Allison2013; Raymond et al., Reference Raymond, Colvin, Jabeen, Nagashima, Barthel, Hadidjojo, Popova, Pejaver and Lubensky2014) and in vivo (Choi et al., Reference Choi, Law, Chien, Link and Wong2010; Watanabe et al., Reference Watanabe, Nishimura, Oka, Nomoto, Kon, Shintou, Hirano, Shimada, Umemoto, Kuroyanagi, Wang, Zhang, Nishimura, Miyazaki, Imamura and Tanaka2010; Clark et al., Reference Clark, Winter, Cohen and Link2011). Ex vivo studies of the zebrafish cone mosaic reveal a highly-organized tiling of four spectrally-distinct cone submosaics across the retina (Allison et al., Reference Allison, Barthel, Skebo, Takechi, Kawamura and Raymond2010; Salbreux et al., Reference Salbreux, Barthel, Raymond and Lubensky2012). Modification of a fluorescent stereomicroscope to include a custom gonioscopic lens has also enabled imaging of the cone mosaic in vivo (Duval et al., Reference Duval, Chung, Lehmann and Allison2013), however, the images are of a relatively small retinal area and contain significant edge distortion. To cover a larger retinal area, several labs have adapted other ophthalmic imaging modalities for use in zebrafish. In one approach, Cheng et al. (Reference Cheng, Yu, Wu, Huang, Chu, Farkas, Nicolau and Leif2010) and Bell et al. (Reference Bell, Xie, Yuan, Kaul, Hollyfield and Anand-Apte2014) used confocal scanning laser ophthalmoscopy to image the zebrafish retina in vivo. Fundus photography has also been used to image the optic disc, vasculature, and photoreceptors in the living zebrafish (Tschopp et al., Reference Tschopp, Takamiya, Cerveny, Gestri, Biehlmaier, Wilson, Strähle and Neuhauss2010), though images of the photoreceptors with this method were not of high resolution. In addition, a major problem with many of these in vivo techniques is uncertainty regarding the lateral scale of the images, which limits the accuracy of any quantitative analysis.
Optical coherence tomography (OCT) permits non-invasive, high-resolution imaging of the eye in a variety of animal models (McLellan & Rasmussen, Reference McLellan and Rasmussen2012). Compared to the previous techniques, OCT covers a wide field of view and has increased axial resolution (<5 microns). Recently, OCT has been applied to zebrafish to image the inner retina, lens, and anterior segment (Rao et al., Reference Rao, Verma, Patel and Gupta2006; Verma et al., Reference Verma, Rao, Suresh, Patel and Gupta2007; Bailey et al., Reference Bailey, Davis, Vance and Hyde2012; Weber et al., Reference Weber, Hochmann, Cimalla, Gärtner, Kuscha, Hans, Geffarth, Kaslin, Koch and Brand2013; Collery et al., Reference Collery, Veth, Dubis, Carroll and Link2014). Given the interest in the photoreceptor layer, in particular its regenerative capacity (Wan & Goldman, Reference Wan and Goldman2016), we wanted to use OCT to image the zebrafish cone mosaic. The Bioptigen OCT built-in software uses a straight slab to generate en face projections, but due to the inherent curvature of the retina, the straight slab produces en face images that are not specific to a single layer of interest. In addition, since the probes used were designed for imaging the mouse retina, the lateral scale of images obtained in zebrafish is unknown (though recent work from Bailey et al. (Reference Bailey, Davis, Vance and Hyde2012) provided an OCT-histology correlation on the axial scaling of zebrafish OCT imagery). Here we sought to improve the method for generating en face projections, as well as determine the lateral scale of OCT images obtained in zebrafish. Using our methods, we demonstrate that quantitative data from OCT images of the photoreceptor mosaic agree with previously published histology data. Application of these methods will enable longitudinal, quantifiable imaging of zebrafish retinal structure in healthy and disease states.
Materials and methods
To determine scaling of OCT images, axial length and retinal imaging were performed on seven fli1:eGFP zebrafish, which have fluorescently labeled vascular endothelial cells (Lawson & Weinstein, Reference Lawson and Weinstein2002). Zebrafish were anesthetized with 0.016% tricaine methanesulfonate, positioned on the imaging stage, and secured using a clay restraint (Collery et al., Reference Collery, Veth, Dubis, Carroll and Link2014). Axial length was determined using the optical path length measured from images obtained using the Bioptigen Envisu R2200 SD-OCT (Bioptigen, Research Triangle Park, NC) with a broadband source (central wavelength 878.4 nm, 186.3 nm bandwidth; Superlum, Enterprise Park, Cork, Ireland) and a 12-mm telecentric lens as previously described (Collery et al., Reference Collery, Veth, Dubis, Carroll and Link2014). High-resolution images of the retina were obtained using the mouse retina probe. Volume scans were nominally 1.2 × 1.2 mm with isotropic sampling (750 A scans/B scan; 750 B scans). Raw OCT scans for retinal images were exported and processed using a custom OCT volume viewer (Java software, Oracle Corporation, Redwood Shores, CA), in which an adjustable contour is used to generate en face summed volume projection (SVP) images (Flatter et al., Reference Flatter, Cooper, Dubow, Pinhas, Singh, Kapur, Shah, Walsh, Hong, Weinberg, Stepien, Wirostko, Robison, Dubra, Rosen, Connor and Carroll2014; Scoles et al., Reference Scoles, Flatter, Cooper, Langlo, Robison, Neitz, Weinberg, Pennesi, Han, Dubra and Carroll2016). For a given B-scan, 3–15 control points were added to the initial contour, where each control point is manually adjusted to follow the contour of the layer(s) of interest. Contour thickness was adjusted to the maximum width of the retinal sublamina of interest, typically 10–20 pixels. The contour was manually adjusted for each B-scan in the volume, correcting for local changes in layer topography and gross changes in axial position due to breathing artifacts. The OCT volume viewing code is available on request. Multiple en face images can be generated for each OCT volume, resulting in images of different retinal features (e.g., inner retinal vasculature and photoreceptor mosaic).
Following OCT imaging, the fli1:eGFP fish were terminally anesthetized and decapitated, and heads were fixed in 4% formaldehyde overnight. The eyes were enucleated and the anterior segment removed from the eyecup. The eyecups were imaged with a Nikon Eclipse E600FN confocal fluorescent microscope with a Nikon D-eclipse C1 camera attachment (Nikon, Tokyo) to view the fluorescently labeled vasculature of the retina. Of the 14 eyes imaged, nine eyes (from seven fish) had minimal post-mortem distortion and were included for subsequent analysis. From microscopy images, the distance between blood vessel branch points was measured in microns using Nikon's EZ-C1 3.90 Free Viewer, with a minimum of three measurements per eye. Accuracy of the fluorescent microscopy measurements was confirmed by imaging a calibration slide at the same magnification. Identical blood vessel branch point measurements were made on the en face OCT images using ImageJ (Schneider et al., Reference Schneider, Rasband and Eliceiri2012). The ratio of ex vivo (microns) to in vivo (pixels) distance was calculated for each corresponding measurement and averaged for each eye. This micron/pixel ratio was used to calculate the actual scan width by multiplying it by the number of A scans/B scan (the scan width in pixels). The ratio of the actual scan width to the nominal scan width (calculated based on the known scan angle and assumed mouse optics) was then plotted against axial length to determine lateral scaling of OCT images as a function of eye length.
An additional five wild type zebrafish were imaged following the protocol above to gather data on the peripheral photoreceptor mosaic, which was defined as 20–50 microns from the optic nerve and corresponded to the transition from disorganized embryonic growth to crystalline adult growth. En face images were generated using the OCT volume viewer (Flatter et al., Reference Flatter, Cooper, Dubow, Pinhas, Singh, Kapur, Shah, Walsh, Hong, Weinberg, Stepien, Wirostko, Robison, Dubra, Rosen, Connor and Carroll2014; Scoles et al., Reference Scoles, Flatter, Cooper, Langlo, Robison, Neitz, Weinberg, Pennesi, Han, Dubra and Carroll2016). Contours were positioned at the most anterior photoreceptor layer to generate en face images of the UV cone layer and at the most posterior photoreceptor layer to generate en face images of the red/green cone layer (Branchek & Bremiller, Reference Branchek and Bremiller1984). Cones were identified using a semi-automated algorithm with manual correction (Garrioch et al., Reference Garrioch, Langlo, Dubis, Cooper, Dubra and Carroll2012). Density and mosaic geometry were assessed from the resultant cone coordinates using a custom program as previously described (Cooper et al., Reference Cooper, Wilk, Tarima and Carroll2016).
Results
Using the custom volume viewer, distinct sublaminas are easily visualized and are uniform across the image. Fig. 1 illustrates representative SVP image quality that can be obtained using the custom OCT volume viewer compared to the built-in software. By creating a custom contour (Fig. 1B), we can generate SVP images of a single retinal layer (UV cone layer shown in Fig. 1D), compared to the corresponding image generated from a flat slab as done with the built-in Bioptigen software (Fig. 1A and 1C). The custom software generates significantly improved images compared to commercially available software.

Fig. 1. Visualization of the zebrafish cone mosaic using en face volume projection images. (A, C) The straight slab method in the native Bioptigen analysis software results in an en face volume projection with different retinal layers included at different locations within the volume, thus the cone mosaic is only visible in some parts of the en face image. (B, D) Using a custom contour that follows a single outer hyper-reflective band results in an en face projection of that layer, in this case the UV cone mosaic.
Using en face images generated at the level above the nerve fiber layer, we were able to identify distinct branch points in the retinal blood vessels (Fig. 2A). The corresponding ex vivo measurements are shown in Fig. 2B. The ratio of the actual scan width to the nominal scan width (scaling coefficient) was plotted as a function of axial length (Fig. 2C). There is a significant positive correlation between the scaling coefficient and axial length (r = 0.98, P < 0.0001; Pearson's correlation), such that as axial length increases, the actual retinal area covered by the scan increases. A linear fit to these data provides a mathematical formula for calculating the scaling coefficient (s) for any zebrafish OCT based on axial length. This can be calculated as follows:
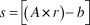 $$s = \left[ {\left( {A \times r} \right) - b} \right]$$
$$s = \left[ {\left( {A \times r} \right) - b} \right]$$
where A is the axial length in millimeters, and b and r are fixed values 0.2187 and 0.4059, respectively. It should be noted that this relationship has been adjusted to account for differences in scan settings and is therefore independent of the field of view (FOV), number of A scans/B scan, or number of B scans used.

Fig. 2. Deriving the lateral scale of in vivo OCT images of the fli1:eGFP zebrafish retina. (A) En face image generated by positioning the custom contour within the retinal nerve fiber layer. Measurements (in pixels) were taken between multiple blood vessel branch points (white dots). (B) Corresponding ex vivo fluorescent microscopy image of the same retina, with measurements (in µm) taken between the same blood vessel branch points in (A). The OCT:microscopy measurements were averaged for each eye and used to determine the size of the OCT scan in µm. A scaling coefficient for each scan was calculated as the ratio between the actual size of the OCT scan to the nominal OCT scan size (in this case, 1200 µm). (C) The scaling coefficient for each scan was plotted against the axial length for that eye and fit with a linear model. Error bars represent one standard deviation for each eye.
Wild-type zebrafish were imaged and analyzed as described above for the fli1-eGFP fish. Using the scaling formula [eqn (1)] and the axial length, we determined the size of the OCT scan for each eye. The en face images of the cone mosaic produced with this method show similar packing arrangements and photoreceptor tiering that have been previously observed ex vivo (Allison et al., Reference Allison, Barthel, Skebo, Takechi, Kawamura and Raymond2010; Tarboush et al., Reference Tarboush, Chapman and Connaughton2012; Ramsey & Perkins, Reference Ramsey and Perkins2013). Contours generated at the innermost photoreceptor layer (corresponding to the UV photoreceptor layer) (Fig. 3A) and the outermost photoreceptor layer (corresponding to the red/green photoreceptor layer) (Fig. 3B), produce en face images with non-overlapping cone structures (Fig. 3C and 3D). A false color overlay (Fig. 3E and 3F) shows that these separate photoreceptor layers are interleaved with one another as seen in histology (Salbreux et al., Reference Salbreux, Barthel, Raymond and Lubensky2012). We are able to observe the disorganized packing near the optic nerve, made up of cones that formed during larval development (Fig. 4C and 4E), and the crystalline organization of the cones that developed as the fish transitioned to adulthood (Fig. 4B and 4D). Analysis of the peripheral UV cone sublamina shows a highly crystalline photoreceptor mosaic with 67.6 ± 11.0% of UV cones displaying 6-sided geometry. Within the images analyzed, we observed regions of even greater regularity, with some patches of the mosaic having 96% six-sided Voronoi cells; these patches were as large as 0.09 mm2. Average UV cone density was found to be 19,557 ± 4,716 cones/mm2, with values ranging from 12,593 to 25,756 cones/mm2. These values agree with previously published wild-type histology values ranging from about 6,000 to 23,000 cones/mm2 (Allison et al., Reference Allison, Barthel, Skebo, Takechi, Kawamura and Raymond2010; Salbreux et al., Reference Salbreux, Barthel, Raymond and Lubensky2012; Duval et al., Reference Duval, Chung, Lehmann and Allison2013; Raymond et al., Reference Raymond, Colvin, Jabeen, Nagashima, Barthel, Hadidjojo, Popova, Pejaver and Lubensky2014).

Fig. 3. Visualization of the UV and red/green photoreceptor layers. (A) B scan with contour placed at the UV layer and (B) the same B scan with the contour moved to the red/green layer. The resultant images generated produce en face images of (C) the UV cone layer and (D) the red/green cone layer. A false-colored overlay (E) of these two layers showing the alternating/interleaved geometry of these photoreceptor sublamina. Scale bar = 200 µm. (F) A cropped region shows the interleaved red/green and UV cone mosaics.

Fig. 4. Regional differences in the topography of the zebrafish cone mosaic. (A) Montage of five en face images of the adult zebrafish UV cone sublamina. Scale bar = 200 µm. Cropped images of the (B) peripheral adult growth and (C) parapapillary larval growth, showing differences in the percentage of six-sided cells within the cone mosaic. The corresponding Voronoi diagrams indicate the number of sides for each Voronoi cell (purple = 4; light blue = 5; green = 6; yellow = 7; red = 8; dark blue = 9). (D) The peripheral adult growth (blue box in A) shows a regular, crystalline mosaic, indicated by the high number of six-sided Voronoi cells (green cells). (E) In contrast, the parapapillary larval growth (orange box in A) shows disorganized, random packing.
Discussion
Here we present a method for deriving accurate, quantitative measures of the zebrafish cone mosaic. The main improvements of our approach include utilizing a custom contour method to generate en face images from the OCT volumes as well as deriving accurate estimates of the lateral scale of the in vivo images acquired using this system. Individual differences in axial length (which affect optical magnification) have been shown to affect the accuracy of OCT measurements in the human retina (Odell et al., Reference Odell, Dubis, Lever, Stepien and Carroll2011; Parthasarathy & Bhende, Reference Parthasarathy and Bhende2015). In animals, correction of the lateral magnification of OCT images in the rat eye has been done by developing a schematic model eye (Lozano & Twa, Reference Lozano and Twa2013), and correction of the axial scale of OCT images in the zebrafish eye has been done using correlative histology (Bailey et al., Reference Bailey, Davis, Vance and Hyde2012); however, to our knowledge, lateral scaling has not previously been determined for zebrafish. It should be noted that our method applies only to this specific OCT system (Bioptigen OCT with mouse probe), and a similar one-time calibration would be needed to determine scaling for different OCT systems. As a result of our methodology, we were able to make quantitative in vivo measurements of the adult zebrafish cone mosaic.
The current study had some limitations. First, analysis of the cone mosaic was limited to the UV sublamina, which has larger cones that are spaced farther apart than the other sublamina (Robinson et al., Reference Robinson, Schmitt, Harosi, Reece and Dowling1993). Although the red/green (L/M) cone sublamina can be imaged using this method (Huckenpahler et al., Reference Huckenpahler, Wilk, Cooper, Carroll, Link and Collery2016), the small size and compact spacing are sometimes below the resolution of the OCT. Similarly, the blue (S) cone sublamina is infrequently distinguished with our current system. System improvements such as using a bore designed specifically for the zebrafish eye, or employing adaptive optics OCT (AO-OCT) for imaging (Jian et al., Reference Jian, Zawadzki and Sarunic2013; Levine et al., Reference Levine, Zawadzki, Cheng, Simon, Pugh and Burns2013) could improve the resolution. A second limitation is that the creation of custom contours for en face images is a time-consuming process, as the contours are adjusted for each B scan within the volume. Combining our en face method with automated segmentation algorithms (Garvin et al., Reference Garvin, Abramoff, Wu, Russell, Burns and Sonka2009; Wang et al., Reference Wang, Kardon, Kupersmith and Garvin2012; Chiu et al., Reference Chiu, Lokhnygina, Dubis, Dubra, Carroll, Izatt and Farsiu2013) could significantly improve processing time. A final limitation is the inability to combine fluorescent labeling with this imaging approach, though molecular labels that manipulate light scattering, such as gold nanoparticles, have been successfully used with OCT (Hayashi et al., Reference Hayashi, Naseri, Pennesi and de Juan2009; de la Zerda et al., Reference de la Zerda, Prabhulkar, Perez, Ruggeri, Paranjape, Habte, Gambhir and Awdeh2015). Such tools may improve the ability to label specific cell populations within the OCT volumes (and thus the en face images).
Despite these limitations, OCT permits non-invasive, high resolution imaging of the eye in a variety of animal models (McLellan & Rasmussen, Reference McLellan and Rasmussen2012). Although OCT has been largely relegated to measuring retinal thickness, the ability to generate accurately scaled en face images increases the application of this method for structural and functional studies. In humans, OCT en face images are used to identify and track retinal lesions and to monitor disease progression (Sallo et al., Reference Sallo, Peto, Egan, Wolf-Schnurrbusch, Clemons, Gillies, Pauleikhoff, Rubin, Chew and Bird2012; Ferrara et al., Reference Ferrara, Mohler, Waheed, Adhi, Liu, Grulkowski, Kraus, Baumal, Hornegger, Fujimoto and Duker2014; Flatter et al., Reference Flatter, Cooper, Dubow, Pinhas, Singh, Kapur, Shah, Walsh, Hong, Weinberg, Stepien, Wirostko, Robison, Dubra, Rosen, Connor and Carroll2014; Mohammad et al., Reference Mohammad, Wanek, Zelkha, Lim, Chen and Shahidi2014; Hood et al., Reference Hood, Fortune, Mavrommatis, Reynaud, Ramachandran, Ritch, Rosen, Muhammad, Dubra and Chui2015). Studies in frogs and mice have applied en face imaging to study phototropic changes and dysfunction in photoreceptors (Zhang et al., Reference Zhang, Lu, Curcio and Yao2012; Lu et al., Reference Lu, Levy, Zhang, Pittler and Yao2013) and ex vivo en face imaging has been used to study photoreceptor development in zebrafish (Mitchell et al., Reference Mitchell, Stevens, Frey, Hunter, Ashino, Kawamura and Stenkamp2015). The in vivo nature of the method described here permits longitudinal assessment of the mosaic in the same animal over time and we foresee this technique having wide application to developmental biology, drug discovery, and disease modeling. We anticipate that researchers would likely use this technique to study longitudinal changes to the cone mosaic in normal development, or in response to disease. However, given the popularity of the zebrafish for drug screening, we also envision a more clinical application where OCT imaging is used to monitor drug oculotoxicity or efficacy prior to FDA approval. Moving forward, applying these imaging methods and robust analysis tools in a variety of disease models could significantly improve our understanding of retinal structure and disease.
Acknowledgments
The authors would like to thank Christine Skumatz, Christopher Langlo, and Joe Vance for assistance with this study. Research reported in this publication was supported by the National Eye Institute under award numbers R01EY016060, T32EY014537, P30EY001931 and the National Institute of General Medical Sciences under award number T32GM080202. This investigation was conducted in a facility constructed with support from the Research Facilities Improvement Program, Grant Number C06RR016511, from the National Center for Research Resources, National Institutes of Health. The content is solely the responsibility of the authors and does not necessarily represent the official views of the National Institutes of Health. Additional support was provided by an unrestricted departmental grant from Research to Prevent Blindness.





 Open access
Open access