


Many abnormalities of regional brain activity have been reported in schizophrenia (Reference Liddle, Reveley and DeakinLiddle, 2000), but there is little consensus as to which abnormalities are stable and thus potential trait markers, and which are transient markers of current clinical state. In a recently completed study we observed that relative to control participants, clinically stable, medicated people with schizophrenia exhibited abnormal activity in the dorsolateral prefrontal cortex, cingulate gyrus and cerebellum during performance of the ‘N-back’ working memory task (Reference Mendrek, Kiehl and SmithMendrek et al, 2004). These results provided partial support for both the ‘cognitive dysmetria’ model of schizophrenia, which suggests that reduced activation in the fronto-thalamo-cerebellar circuitry is a core pathophysiological feature of the illness (Reference AndreasenAndreasen, 1999), and for the proposition that corticolimbic system dysregulation has a significant role in the formation of psychotic symptoms (Reference BenesBenes, 2000). The study reported here was designed to explore the pattern of cerebral function in a group of patients with first-episode schizophrenia during two stages of their illness – acute psychosis and partial remission – in order to determine which elements of the distorted circuitry are stable over time and which fluctuate with change in clinical state. Based on recent theories and empirical findings in schizophrenia research (Reference Lahti, Holcomb and MedoffLahti et al, 1995; Reference Andreasen, O'Leary and CizadloAndreasen et al, 1996; Reference Crespo-Facorro, Paradiso and AndreasenCrespo-Facorro et al, 1999; Reference Holcomb, Lahti and MedoffHolcomb et al, 2000), our hypothesis was that the disturbance of fronto-thalamo-cerebellar circuitry in patients with first-episode schizophrenia would remain under-activated over time, whereas the function of the cingulate gyrus would vary with changes in clinical status.
METHOD
Participants
Ten people with first-episode schizophrenia, recruited within a week of their admission to the psychiatric ward of the University of British Columbia Hospital in Vancouver, participated in the study. All patients were judged by the attending physicians to be competent to give informed consent and capable of cooperating during the imaging procedure. All patients initially diagnosed as having a brief psychotic disorder, schizophreniform disorder or schizophrenia were subsequently given a diagnosis of schizophrenia according to DSM–IV criteria (American Psychiatric Association, 1994). Patients underwent a functional magnetic resonance imaging (fMRI) brain scan during their first week of antipsychotic treatment and were scanned again after a further 6–8 weeks of treatment. The first scan took place after approximately 3 days of antipsychotic treatment to minimise the problem of acute agitation during the procedure although initial dosages were low to avoid excessive sedation. Patients were treated with atypical antipsychotic medication (either risperidone or olanzapine) throughout the study, at a dose judged to be optimal on clinical grounds. As a result of the study design, we were unable to detect the immediate consequences of antipsychotic treatment; on the other hand, because patients were receiving similar medication during the first and second scanning sessions, the observed systematic changes in brain function are likely to reflect the gradual changes in clinical status during sustained treatment, rather than immediate pharmacological effects. The data obtained from two patients were discarded from the final analysis because of failure to meet the predetermined acceptable level of accuracy in the task performed during scanning. At the beginning of the study, seven of the remaining eight patients were receiving treatment with risperidone (mean daily dosage 3 mg, s.d.=0.4) and one was taking olanzapine (10 mg); at the end of the study, seven were taking risperidone (mean daily dosage 2.7 mg, s.d.=0.3), two were taking olanzapine (mean daily dosage 12.5 mg, s.d.=2.5) and one patient was receiving both medications (1 mg risperidone and 10 mg olanzapine).
In addition to an interview and a review of case notes to corroborate the diagnosis of schizophrenia and assess changes in symptoms over time, all patients were assessed with the Signs and Symptoms of Psychotic Illness scale (SSPI; Reference Liddle, Ngan and DuffieldLiddle et al, 2002), which measures the severity of 20 signs and symptoms of acute and chronic psychotic illness; a score of 18 (s.d.=7) is typical of an acute psychotic state (Reference Liddle, Ngan and DuffieldLiddle et al, 2002). The group of patients in our study obtained a mean score of 19.85 (s.d.=10.72) during the first assessment with the SSPI (within the first week of treatment) and a mean score of 13.0 (s.d.=10.78) during the second assessment (after 6–8 weeks of treatment).
The group of eight healthy control participants, with no current or past psychotic illness and no psychotic illness in a first-degree relative, did not differ from the group of patients in terms of age, gender, parental socio-economic status or IQ (Table 1). Parental socio-economic status was assessed using the Hollingshead criteria for parental social position (Reference Hollingshead and RedlichHollingshead & Redlich, 1958) and IQ was measured using the Quick test (Reference Ammons and AmmonsAmmons & Ammons, 1962). All participants were right-handed according to the Annett Handedness scale (Reference AnnettAnnett, 1970). The study was approved by the ethics review committee of the University of British Columbia. All participants gave written consent after the experimental details were explained to them.
Table 1 Demographic characteristics of the study sample

| Control group | Schizophrenia group | |
|---|---|---|
| Age, years: mean (s.d.) | 28 (8) | 30 (9) |
| Gender (male/female), n/n | 6/2 | 6/2 |
| IQ: mean (s.d.)1 | 101 (7.3) | 99 (4.9) |
| Socio-economic status: mean (s.d.)2 | 2.63 (1.2) | 2.75 (1.8) |
Behavioural test and data analysis
The behavioural task used in our study was one used previously in neuroimaging investigations of working memory (Reference Awh, Jonides and SmithAwh et al, 1996; Reference Cohen, Perlstein and BraverCohen et al, 1997; Reference Jonides, Schumacher and SmithJonides et al, 1997). This version of the n-back task consisted of a screen display of a succession of letters; the person tested was required to press a button whenever the designated letter appeared. In our study, participants either had to press the button whenever the letter X appeared (the ‘0-back’ task) or press it any time they saw a letter identical to one presented two screens earlier (the ‘2-back’ task). Each letter was displayed for a duration of 250 ms with an inter-stimulus interval of 2 s. Participants were required to complete two runs of alternating 30 s periods of 0-back and 2-back testing, separated by 20 s rest periods; each run lasted about 7 min. Both tasks involved similar sensory processing of information and a similar amount of motor activity. Before scanning, participants were given full instructions and a 3 min practice session, in which they had to reach a 70% accuracy level. Despite successful practice, two patients failed to reach the required 60% level of accuracy during performance of the task in the scanner and their data were discarded from the analysis. All participants performed the task on two separate occasions, 6–8 weeks apart.
The behavioural data were analysed using between–within repeated measures, condition (0-back, 2-back) by group (patients, controls) by scanning session (first, second), factorial analyses of variance (ANOVAs). Separate ANOVAs were used to evaluate performance accuracy and reaction time data. Errors of omission were defined as a failure to respond to the target stimulus within 1500 ms of stimulus onset; errors of commission were defined as a response to a non-target stimulus within the same time frame.
Imaging procedure and analysis
Imaging was performed using a standard clinical GE 1.5 tesla whole-body MRI scanner (General Electric, Milwaukee, Wisconsin, USA) fitted with a Horizon echo-speed upgrade. Whole-brain echo-planar fMRI was performed using a gradient echo pulse sequence (repetition time 3000 ms, echo time 40 ms, flip angle 90°, field of view 24 cm × 24 cm, 64 × 64 matrix, band width 62.5 kHz, 3.75 × 3.75 mm inplane resolution, 5 mm slice thickness, 29 slices). The functional images acquired in each run were reconstructed offline and subsequently realigned, motion corrected and normalised into the MRI template and stereotactic space (Reference Talairach and TournouxTalairach & Tournoux, 1988) using Statistical Parametric Mapping software (SPM99; Wellcome Department of Cognitive Neurology, Institute of Neurology, London). The motion estimates for individual participants did not exceed 3 mm or 3°, and there was no significant difference between the groups. The realigned and normalised images were smoothed with an 8 mm full-width at half-maximum Gaussian filter.
The statistical analyses were performed using a random effects model as implemented in SPM99 for UNIX. In the computation of this analysis, first the observed time courses of image intensities were temporarily filtered to remove noise associated with low-frequency confounds such as respiration. In addition, each type of epoch (i.e. 2-back, 0-back and rest) was modelled by a boxcar waveform with a temporal delay of 6 s to account for the relatively slow onset of the haemodynamic response. Then, single images for each participant in each session were created based on the 2-back v. 0-back and 0-back v. rest contrasts. These contrast images were subsequently entered into a second-level random-effects analysis, which employed t-tests to assess the significance of the planned comparisons between conditions and between groups. The mean difference in cerebral activation between the 2-back and 0-back conditions and the difference between the 0-back and rest condition within each group and within each scanning session were assessed using one-sample t-tests. These analyses were performed for the entire brain volume at the cluster level (P≤0.05 corrected for multiple comparisons, thresholded at P≤0.001) as implemented in SPM99 (Reference Friston, Worsely and FracowiackFriston et al, 1994). The differences between groups in the contrast between 2-back and 0-back and between 0-back and rest in each session were assessed using two-sample t-tests. In addition to the search in the entire brain volume for the within-group analysis, in order to increase statistical sensitivity and to reduce the probability of type 2 error in assessing our hypothesis regarding the fronto-thalamo-cerebellar and corticolimbic systems, the between-group as well as the within-group between-session comparisons were restricted to voxels contained within predefined regions of interest. These regions of interest were eight spheres 12 mm in radius centred on the loci of peak activation in the brain regions relevant to our hypothesis (previously identified in a pilot study of a separate group of 11 healthy volunteers), sited in the bilateral dorsolateral prefrontal cortex, bilateral thalamus, bilateral cerebellum and the anterior and posterior cingulate gyrus (Table 2). In testing for the significance of changes in these regions, we applied the criterion of P≤0.005. In order to avoid global normalisation artefacts (Reference AnderssonAndersson, 1997; Reference Aguirre, Zarahn and D'EspositoAguirre et al, 1998; Reference Desjardins, Kiehl and LiddleDesjardins et al, 2001), this procedure was not performed.
Table 2 Talairach coordinates of maximally activated voxels during the 2-back v. 0-back condition by 11 healthy volunteers in the pilot study

| Region of interest | Talairach coordinates | ||
|---|---|---|---|
| x | y | z | |
| 2-back > 0-back | |||
| Left middle frontal gyrus (BA 46) | -48 | 34 | 20 |
| Right middle frontal gyrus (BA 46) | 52 | 34 | 20 |
| Left cerebellum | -38 | -60 | -25 |
| Right cerebellum | 34 | -60 | -30 |
| Left thalamus | -12 | -6 | 10 |
| Right thalamus | 16 | -4 | 10 |
| 0-back > 2-back | |||
| Left posterior cingulate gyrus (BA 23) | -4 | -54 | 20 |
| Left anterior cingulate gyrus (BA 24) | -4 | 48 | 10 |
RESULTS
Behavioural data
The paired t-tests revealed that in the schizophrenia group the participants’ symptoms, assessed using the SSPI scale during the first week of treatment, diminished significantly after 6–8 weeks of antipsychotic medication: t (7)=9.26, P<0.001. The between–within repeated measures factorial ANOVA for the errors of omission during performance of the working memory task in two scanning sessions showed that patients performed significantly worse than control participants (main effect of group, F (1, 14)=14.1, P<0.01) and that both groups made significantly fewer errors in the 0-back task than in the 2-back task (main effect of condition, F (1, 14)=21.58, P<0.001). There was no group × condition interaction (P>0.05) but a statistically significant condition × scanning session interaction (F (1, 14)=6.96, P<0.05) was present, reflecting a reduction in error rate during the second scanning session. The performance accuracy is illustrated in Fig. 1. The separate ANOVA for the errors of commission did not reveal any statistically significant results. Finally, the ANOVA of reaction time data showed that it took significantly longer to respond correctly in the 2-back task than in the 0-back task (main effect of condition, F (1, 14)=48.52, P<0.001) and that patients were slower at responding than were control participants (main effect of group, F (1, 14)=30.02, P<0.001). A significant group × condition interaction was also present (F (1, 14)=7.3, P<0.05).

Fig. 1 Mean number of errors of omission by first-episode patients (squares) and control participants (circles) during the n-back task in the first (solid lines) and second (dashed lines) scanning sessions.
Imaging data
The analyses of the pattern of cerebral activity during the 2-back v. 0-back condition in healthy participants demonstrated significant activations in bilateral prefrontal, premotor and parietal cortex, as well as in bilateral cerebellum and thalamus (Table 3, Fig. 2a). The 0-back v. rest comparison revealed significant findings in the supplementary motor area, but not in any of the above-mentioned regions implicated in a working memory function (Table 3, Fig. 2b). The same pattern of activity was apparent in these participants, during the second scanning session 6–8 weeks later (Table 3, Fig. 2c, d).

Fig. 2 Group statistical parametric maps of activation during performance of the n-back task by control participants in the first scanning session (top panel: (a) 2-back v. 0-back, (b) 0-back v. rest) and the second scanning session (bottom panel: (c) 2-back v. 0-back, (d) 0-back v. rest) (P≤0.001, uncorrected).
Table 3 Significant areas of activation in control participants during the behavioural tasks

| Region of interest | Talairach coordinates | Local maxima z score | ||
|---|---|---|---|---|
| x | y | z | ||
| First scanning session | ||||
| 2-back v. 0-back | ||||
| Right inferior parietal lobule (BA 40) | 36 | -48 | 40 | 6.01 |
| Right middle frontal gyrus (BA 6) | 48 | 8 | 32 | 5.88 |
| Left inferior parietal lobule (BA 40) | -40 | -52 | 44 | 5.57 |
| Left middle frontal gyrus (BA 6) | -24 | -4 | 52 | 5.48 |
| Left cerebellum | -40 | -60 | -40 | 5.14 |
| Left thalamus | -16 | -4 | 4 | 4.61 |
| Right cerebellum | 24 | -64 | -40 | 4.33 |
| Right thalamus | 8 | -24 | 8 | 4.00 |
| 0-back v. rest | ||||
| Left superior frontal gyrus (BA 6) | -8 | -8 | 56 | 5.82 |
| Right superior frontal gyrus (BA 6) | 8 | -8 | 64 | 5.43 |
| Second scanning session | ||||
| 2-back v. 0-back | ||||
| Left superior parietal lobule (BA 7) | -28 | -68 | 44 | 6.31 |
| Right superior parietal lobule (BA 7) | 36 | -64 | 44 | 5.91 |
| Left middle frontal gyrus (BA 6) | -20 | -4 | 52 | 5.83 |
| Left cerebellum | -32 | -56 | -40 | 5.58 |
| Right middle frontal gyrus (BA 6) | 32 | -4 | 56 | 5.28 |
| Right cerebellum | 40 | -68 | -36 | 5.35 |
| Left thalamus | -8 | -24 | 12 | 4.99 |
| Right thalamus | 8 | -20 | 16 | 4.83 |
| 0-back v. rest | ||||
| Left superior frontal gyrus (BA 6) | -8 | -8 | 60 | 5.26 |
| Right superior frontal gyrus (BA 6) | 8 | -4 | 64 | 4.90 |
In contrast, the comparison between the 2-back and 0-back tasks in the acutely ill patients revealed activation only in a few isolated clusters of the prefrontal and parietal cortex (Table 4, Fig. 3a). Analysis of 0-back v. rest demonstrated significant and widespread pattern of activations in the bilateral prefrontal and parietal cortex, thalamus and cerebellum (Table 4, Fig. 3b). This ‘shifted’ pattern of activity, with extensive activation during the 0-back task relative to rest and minimal increase in activation from 0-back to 2-back, normalised over time with treatment, although not completely. Thus, after 6–8 weeks of treatment, accompanied by a significant improvement in symptoms, the activation during the 0-back v. rest condition diminished (Table 4, Fig. 3c) and the activation of 2-back v. 0-back had increased (Table 4, Fig. 3d).

Fig. 3 Group statistical parametric maps of activation during performance of the n-back task by patients in the first scanning session (top panel: (a) 2-back v. 0-back, (b) 0-back v. rest) and the second scanning session (bottom panel: (c) 2-back v. 0-back, (d) 0-back v. rest) (P≤0.001, uncorrected).
Table 4 Significant areas of activation in participants with schizophrenia during the behavioural tasks

| Region of interest | Talairach coordinates | Local maxima z score | ||
|---|---|---|---|---|
| x | y | z | ||
| First scanning session | ||||
| 2-back v. 0-back | ||||
| Left superior frontal gyrus (BA 6) | -4 | 12 | 56 | 4.40 |
| Left inferior frontal gyrus (BA 47) | -40 | 16 | -8 | 4.26 |
| Right inferior frontal gyrus (BA 44) | 52 | 12 | 16 | 4.00 |
| Right inferior parietal lobule (BA 40) | 44 | -44 | 44 | 4.11 |
| Left inferior parietal lobule (BA 40) | -40 | -48 | 44 | 3.35 |
| 0-back v. rest | ||||
| Right middle temporal gyrus (BA 21) | 52 | -48 | 0 | 5.10 |
| Right cerebellum | 4 | -76 | -40 | 4.99 |
| Left middle frontal gyrus (BA 6) | -44 | -6 | 40 | 4.98 |
| Left superior parietal lobule (BA 7) | -36 | -44 | 48 | 4.91 |
| Right inferior parietal lobule (BA 40) | 48 | -52 | 40 | 4.60 |
| Second scanning session | ||||
| 2-back v. 0-back | ||||
| Left superior parietal lobule (BA 7) | -28 | -68 | 44 | 6.31 |
| Right inferior parietal lobule (BA 40) | 36 | -64 | 44 | 5.91 |
| Left middle frontal gyrus (BA 6) | -20 | -4 | 52 | 5.83 |
| Right middle frontal gyrus (BA 6) | 32 | -4 | 56 | 5.28 |
| Right cerebellum | 28 | -64 | -36 | 4.65 |
| Left cerebellum | -32 | -64 | -36 | 4.63 |
| Right thalamus | 20 | -20 | 16 | 4.51 |
| 0-back v. rest | ||||
| Left insula | -32 | -24 | 28 | 4.29 |
| Left superior frontal gyrus (BA 6) | -12 | -12 | 68 | 4.23 |
The above phenomenon is illustrated by the magnitude of activation of the left dorsolateral prefrontal cortex in the different conditions in the two groups of participants (Fig. 4). Thus, healthy participants showed no activation of this brain area during the 0-back task but significant activation during the 2-back task, resulting in a highly significant difference between the two conditions. In comparison, patients with acute psychosis activated dorsolateral prefrontal cortex to a comparable level during both tasks, resulting in a non-significant difference between the two. In effect, the patients exhibited relative hyperfrontality during the 0-back task compared with controls. Importantly, at this session they did not achieve the magnitude of activation present in healthy participants during the 2-back task. In patients in remission the difference in dorsolateral prefrontal cortex activation was detectable between the two test conditions, but again this brain area did not attain the levels of activity apparent in controls during the 2-back task.
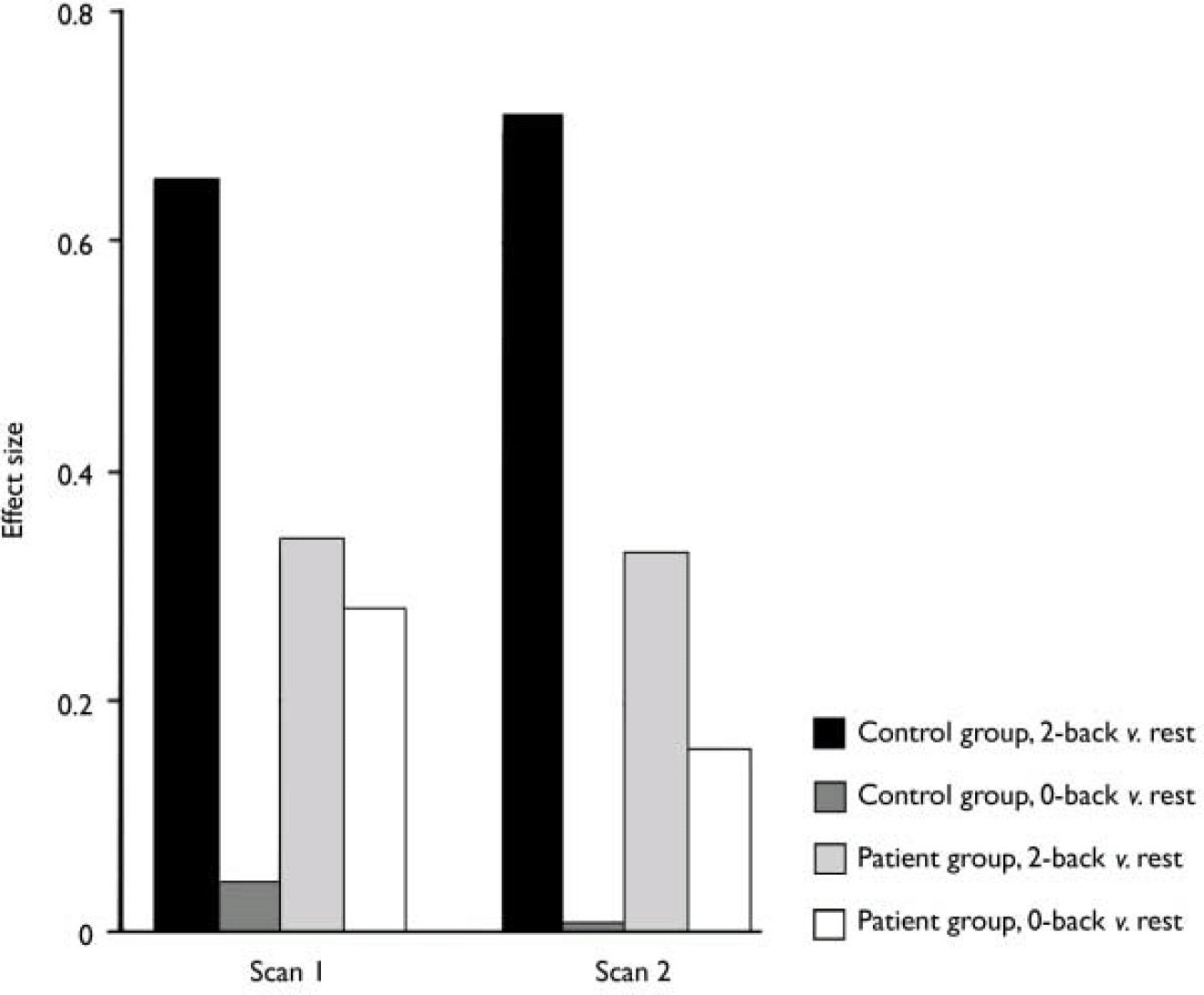
Fig. 4 Left dorsolateral prefrontal cortex activation during the 2-back and 0-back tasks v. rest in the two scanning sessions.
The between-group analysis of data obtained during the first scanning session revealed that in comparison with control participants, patients exhibited less activity in the specified regions of interest in the bilateral dorsolateral prefrontal cortex, thalamus, cerebellum and posterior cingulate cortex during the 2-back v. 0-back condition. In contrast, the comparison between 0-back and rest revealed greater activity in patients relative to controls in all of the above structures except the left thalamus and right cerebellum (Table 5). The between-group analysis of data obtained at the second scanning session revealed that patients exhibited less activity in specified regions of interest in the left dorsolateral prefrontal cortex, left thalamus and right cerebellum during the 2-back v. 0-back condition and relatively more activity in the left dorsolateral prefrontal cortex during 0-back v. rest (Table 5).
Table 5 Areas of significant difference between patient and control groups during task performance based on random-effects analysis in eight regions of interest (P≤0.005)

| Region of interest | Talairach coordinates | z score | ||
|---|---|---|---|---|
| x | y | z | ||
| First scanning session | ||||
| 2-back v. 0-back, controls > patients | ||||
| Left thalamus | -4 | -4 | 4 | 3.79 |
| Right thalamus | 16 | -16 | 8 | 3.67 |
| Left middle frontal gyrus (BA 46) | -40 | 32 | 16 | 3.58 |
| Right cerebellum | 28 | -56 | -36 | 3.56 |
| Left cerebellum | -40 | -64 | -20 | 3.17 |
| Right middle frontal gyrus (BA 46) | 44 | 44 | 24 | 3.15 |
| Right posterior cingulate gyrus (BA 23) | 12 | -52 | 20 | 3.05 |
| 0-back v. rest, patients > controls | ||||
| Left middle frontal gyrus (BA 46) | -52 | 32 | 16 | 4.42 |
| Right middle frontal gyrus (BA 45) | 48 | 28 | 12 | 3.82 |
| Left cerebellum | -40 | -64 | -32 | 3.80 |
| Right posterior cingulate gyrus (BA 23) | 8 | -52 | 20 | 3.17 |
| Right thalamus | 8 | 4 | 8 | 3.10 |
| Second scanning session | ||||
| 2-back v. 0-back, controls > patients | ||||
| Left thalamus | -16 | -4 | 4 | 3.03 |
| Right cerebellum | 28 | -52 | -36 | 2.81 |
| Left middle frontal gyrus (BA 9) | -52 | 24 | 28 | 2.65 |
| 0-back v. rest, patients > controls | ||||
| Left middle frontal gyrus (BA 45) | -52 | 32 | 12 | 3.37 |
Finally, the comparison of regions of interest between the first and second scanning sessions in healthy participants did not reveal any significant differences, whereas the same comparison in patients revealed significant changes in the activation pattern over time in the right dorsolateral prefrontal cortex, right thalamus, left cerebellum and posterior cingulate (Table 6).
Table 6 Areas of significant difference between patients’ first and second scanning sessions during task performance, based on random-effects analysis in eight regions of interest (P≤0.005)

| Region of interest | Talairach coordinates | z score | ||
|---|---|---|---|---|
| x | y | z | ||
| Relative underactivation in first session | ||||
| 2-back v. 0-back | ||||
| Left cerebellum | -36 | -64 | -16 | 2.82 |
| Left posterior cingulate gyrus (BA 23) | -8 | -52 | 20 | 2.82 |
| Right middle frontal gyrus (BA 46) | 44 | 44 | 20 | 2.76 |
| Right thalamus | 24 | -4 | 16 | 2.64 |
| Relative overactivation in first session | ||||
| 0-back v. rest | ||||
| Left posterior cingulate gyrus (BA 23) | -8 | -52 | 24 | 3.21 |
| Left cerebellum | -40 | -68 | -20 | 3.00 |
| Right thalamus | 16 | -4 | 0 | 2.69 |
DISCUSSION
Principal within-group findings
The results obtained in the group of healthy participants were consistent with our previous findings (Reference Mendrek, Kiehl and SmithMendrek et al, 2004) and with other studies of working memory function in healthy volunteers (Reference D'Esposito, Detre and AlsopD’Esposito et al, 1995; Reference Awh, Jonides and SmithAwh et al, 1996; Reference Cohen, Perlstein and BraverCohen et al, 1997; Reference Jonides, Schumacher and SmithJonides et al, 1997). Thus, the healthy participants exhibited widespread activation in the bilateral prefrontal cortex, parietal cortex, thalamus and cerebellum during the 2-back task v. 0-back condition, and significant activation in a circumscribed region of supplementary motor area during 0-back v. rest (see Fig. 2, Table 3). In marked contrast, the participants with acute psychosis exhibited little activation during the 2-back v. 0-back condition, but extensive activation during 0-back v. rest condition in several of the areas activated in control participants during the 2-back task – namely the bilateral prefrontal cortex, parietal cortex, thalamus and cerebellum (see Fig. 3, Table 4). Thus, the lack of substantial difference between 2-back and 0-back arose from the fact that both tasks produced similar patterns of neural activation in patients with acute psychosis; in other words, the normally undemanding 0-back task was challenging enough for patients to recruit the available cerebral resources to near-maximal capacity, so that there was no possibility of enhancing cerebral function during the 2-back task. The finding of augmented cerebral activation during the 0-back condition is congruent with previous neuroimaging studies in which various working memory tasks enhanced prefrontal cortex activation in patients who performed above the chance level but worse than a control group (Manoach et al, Reference Manoach, Press and Thangaraj1999, Reference Manoach, Gollub and Benson2000; Reference Callicott, Bertolino and MattayCallicott et al, 2000). It is also consistent with the report by Ramsey et al (Reference Ramsey, Koning and Welles2002), who suggested that the inefficiency of neural communication in schizophrenia results in excessive recruitment of neural systems in patients relative to control participants during comparable performances on cognitive tasks.
The pattern of neural activation during performance of the n-back task normalised to some extent over time with antipsychotic treatment and improvement in the clinical state of the patients. Thus, in the second scanning session, patients did not exhibit any anomalous overactivation during the 0-back v. rest condition, but the level of cerebral activation observed during the 2-back v. 0-back condition was still diminished relative to control participants.
Principal between-group findings
The between-group region of interest analyses confirmed and extended the within-group results. Compared with control participants, patients with acute psychosis exhibited significant underactivation bilaterally in the dorsolateral prefrontal cortex, cerebellum, thalamus and posterior cingulate in the 2-back v. 0-back condition, while showing overactivation in all of these regions (with the exception of right cerebellum and left thalamus) during 0-back v. rest. The relative underactivation during the 2-back task coupled with the lack of overactivation during the 0-back task implies true underutilisation of the right cerebellum and left thalamus in acute psychosis (as opposed to maximal activation during 0-back and no additional increase during the 2-back task). Moreover, both right cerebellum and left thalamus remained underused in the second scanning session in patients with partially remitted disease relative to control participants, pointing to the stable nature of this disturbance. This interpretation is supported by the observation of a similar pattern of decreased activation in clinically stable, medicated patients (Reference Mendrek, Kiehl and SmithMendrek et al, 2004). Nevertheless, it must be emphasised that our findings cannot exclude significant effects on regions outside the set of preselected regions of interest.
Like the right cerebellum and left thalamus, the left dorsolateral prefrontal cortex also remained functionally suppressed, although to a lesser degree (see below and Fig. 4 for explanation of this anomaly). In contrast, activation of the right dorsolateral prefrontal cortex, right thalamus, left cerebellum and posterior cingulate normalised over time with antipsychotic treatment and attenuation of symptoms, to a level that was not significantly different from that in the healthy group, suggesting that disturbance of these sites represents a state marker for acute exacerbation of schizophrenia.
The question of changes in the pattern of cerebral activity accompanying the resolution of psychosis was also addressed with the between-session analysis of the patients with schizophrenia, which revealed significant change in activation over time in the right dorsolateral prefrontal cortex, right thalamus, left cerebellum and posterior cingulate. It is important to point out that in these patients the disease was in partial rather than full remission during the second scanning session, and further normalisation of function of these structures could be anticipated with the complete resolution of symptoms.
Lateralisation effect
Overall, the results revealed that the more persistent abnormalities of the acute psychotic state were localised in the left cerebral hemisphere and right cerebellum, whereas more transient features were localised in the right cerebral hemisphere and left cerebellum. However, again it is essential to point out that this conclusion is restricted to areas that were included in the region of interest analysis. The observed relationship between cerebral and cerebellar abnormalities is consistent with existence of contralateral connections between the dorsolateral prefrontal cortex and cerebellum relayed by thalamic nuclei (Reference Middleton and StrickMiddleton & Strick, 2001). Accordingly, the concurrent dysfunction of the dorsolateral prefrontal cortex, cerebellum and thalamus in our schizophrenia group may represent underlying disturbance in the connectivity between these structures. This finding, together with other reports (e.g. Reference Andreasen, O'Leary and CizadloAndreasen et al, 1996; Reference Wiser, Andreasen and O'LearyWiser et al, 1998; Reference Crespo-Facorro, Paradiso and AndreasenCrespo-Facorro et al, 1999), provides support for the ‘cognitive dysmetria’ theory of schizophrenia (Reference AndreasenAndreasen, 1999), while the presence of lateralisation effect adds a new layer of complexity to the model. Moreover, the observation of more persistent abnormalities in the left hemisphere is in line with evidence from structural abnormality studies in schizophrenia, in which most of the studies that found significant asymmetry reported greater abnormality in the left hemisphere (Reference Hopkins, Lewis, Harrison and RobertsHopkins & Lewis, 2000). The persistence of the left hemisphere abnormalities and the transience of the right hemisphere abnormalities during performance of the n-back task are also broadly consistent with the hemispheric imbalance model proposed by Gruzelier (Reference Gruzelier1984). On the basis of evidence from electrophysiological studies, Gruzelier proposed that in schizophrenia symptoms reflecting excitation (which tend to be transient) are associated with right hemisphere dysfunction, whereas symptoms reflecting withdrawal (which tend to be more persistent) are associated with relative underactivity of the left hemisphere.
Prefrontal cortex findings
Our results shed new light on the debate regarding prefrontal function in schizophrenia (Reference Weinberger and BermanWeinberger & Berman, 1996; Reference ManoachManoach, 2003) and may help to reconcile the inconsistent findings of a number of fMRI studies, some of which demonstrated diminished prefrontal activation during working memory tasks in people with schizophrenia relative to control participants (Reference Callicott, Ramsey and TallentCallicott et al, 1998; Reference Carter, Perlstein and GanguliCarter et al, 1998; Reference Stevens, Goldman-Rakic and GoreStevens et al, 1998; Reference Perlstein, Carter and NollPerlstein et al, 2001), whereas others found enhanced activation (Manoach et al, Reference Manoach, Press and Thangaraj1999, Reference Manoach, Gollub and Benson2000; Reference Callicott, Bertolino and MattayCallicott et al, 2000) or no difference between the groups (Reference Honey, Bullmore and SharmaHoney et al, 2002). We observed both overactivation of the prefrontal cortex during the 0-back task and underactivation during the 2-back task in patients relative to controls. This finding fits well with empirical evidence of a nonlinear, inverted U-shaped response in dorsolateral prefrontal cortex function to parametrically increasing working memory difficulty. Thus, in healthy volunteers dorsolateral prefrontal cortex activation increases together with the working memory load; but although the initial reduction in working memory capacity may be associated with relative overactivation of this brain region (Reference Rypma and D'EspositoRypma & D’Esposito, 1999), further decline in the capacity to process information is accompanied by its relative underactivation (Reference Callicott, Mattay and BertolinoCallicott et al, 1999). Manoach (Reference Manoach2003) has proposed that this inverted U-shaped function is shifted to the left in people with schizophrenia, such that the increase, plateau and eventual decrease in activation can be observed with lower working memory loads than in healthy individuals. Our data suggest that in addition to this leftwards shift there might be also a downwards shift in dorsolateral prefrontal cortex function in schizophrenia. Specifically, although prefrontal dysfunction in the patient group was modulated to a certain degree by clinical status and the type of presented challenge, there appears also to exist a persistent abnormal limitation of left dorsolateral prefrontal cortex activation in schizophrenia. This limitation might be related to structural anomaly of this region in schizophrenia reported by Selemon et al (Reference Selemon, Rajkowska and Goldman-Rakic1995) and Rajkowska et al (Reference Rajkowska, Selemon and Goldman-Rakic1998).
Present results in the light of previous longitudinal studies
This study is one of the first longitudinal investigations of a cohort of patients at two different stages of their illness, a study design that has become more feasible with the widespread availability of non-invasive fMRI technology. Partly because of the challenging nature of these studies, only a few such reports in schizophrenia have been published (Reference Honey, Bullmore and SoniHoney et al, 1999; Reference Manoach, Halpern and KramerManoach et al, 2001; Reference Stephan, Magnotta and WhiteStephan et al, 2001). Honey et al (Reference Honey, Bullmore and Soni1999) demonstrated that after switching from typical to atypical antipsychotic medication, patients exhibited increased prefrontal and parietal activation during a working memory task. Stephan et al (Reference Stephan, Magnotta and White2001) tested two groups of people with schizophrenia, one drug-free and one treated with olanzapine and found that the pharmacological treatment normalised cerebellar functional connectivity during a simple motor task. Manoach et al (Reference Manoach, Halpern and Kramer2001) studied test–retest reliability of working memory performance in clinically stable patients with schizophrenia and in healthy volunteers and found that even given reliable task performance and a stable clinical status, individual participants showed variability in cerebral activation, although the group-averaged activation pattern did not differ between the first and second scanning sessions.
Study limitations
Our results are subject to some limitations. For example, the fact that patients were treated throughout the course of the study with antipsychotic medication does not allow us to distinguish changes in brain function attributable to long-term primary pharmacological effects from changes attributable to alteration in clinical state. Specifically, some of our results might be interpreted as evidence that medication ameliorates abnormality in one network (right fronto-thalamo-cerebellar) but not in another (left fronto-thalamo-cerebellar). However, we consider it unlikely that all of the effects we observed reflect the effects of medication. In particular, the differences between patients and controls in the first scanning session are unlikely to be due to medication, as these differences were in the opposite direction to the changes that occurred during sustained medication. Moreover, the differences between the groups in cerebral activation could have arisen partly through the significantly inferior performance on the n-back task of patients relative to control participants. However, although the behavioural differences could have partially contributed to the differential pattern of cerebral activations, overall this explanation is too simplistic, because the performance of patients was inferior on both the 0-back and the 2-back tasks, whereas cerebral activation was increased in the first task and decreased in the second. Furthermore, although the patients’ working memory did improve between the first and second scanning sessions, there was no significant correlation between change in the task performance and cerebral activity. Finally, since we specifically investigated working memory we cannot draw any conclusion about the functional substrates of other cognitive processes.
To summarise, the overall findings of our study suggest that underutilisation of the left dorsolateral prefrontal cortex, left thalamus and right cerebellum represents a stable, potential trait marker of schizophrenia, whereas disturbances in the right dorsolateral prefrontal cortex, right thalamus, left cerebellum and cingulate gyrus are a state-related phenomenon.
Clinical Implications and Limitations
Clinical Implications
-
▪ Underactivations of the left dorsolateral prefrontal cortex, left thalamus and right cerebellum represent a stable potential trait marker of schizophrenia.
-
▪ Abnormalities in the right dorsolateral prefrontal cortex, right thalamus, left cerebellum and cingulate gyrus normalise with improvement in clinical status and thus represent a state-related phenomenon.
-
▪ Identification of trait-related abnormalities could contribute to development of more reliable diagnosis of schizophrenia, whereas identification of state-related changes might help in evaluation of treatments for schizophrenia and/or may serve as predictors of treatment outcome.
Limitations
-
▪ The findings cannot exclude significant effects on regions outside of the set of preselected regions of interest.
-
▪ Patients were treated throughout the course of the study with antipsychotic medications and changes in brain function attributable to pharmacological effects cannot be distinguished from changes attributable to change in clinical state.
-
▪ Because we studied patients with the specific task of working memory we cannot draw any conclusions about the functional substrates of other cognitive processes.
Acknowledgements
The study was supported by grants from the Medical Research Council of Canada and the Norma Calder Schizophrenia Research Foundation, and took place at the Department of Psychiatry of the University of British Columbia. The assistance of Drs Bruce Forster, Alex MacKay and Andra Smith and the invaluable help of magnetic resonance imaging technicians Jennifer McCord, Sylvia Rennenberg and Trudy Shaw and research assistant Cameron Anderson are gratefully acknowledged. We also thank all those who participated in the study.











eLetters
No eLetters have been published for this article.