


INTRODUCTION
Avian influenza viruses (AIVs) are serologically classified into 16 known haemagglutinin subtypes, termed H1–H16 [Reference Alexander1]. Notifiable avian influenza (NAI) in poultry is caused by H5 and H7 AIVs, these subtypes may mutate from low pathogenicity (LP) AIVs to the highly pathogenic (HP) AIVs which are characterized by high morbidity and mortality in gallinaceous poultry flocks [Reference Alexander and Brown2, 3]. Increased interest in the detection of AIV infections in the past decade has been prompted largely by the continuing poultry epidemic due to H5N1 HPAIVs and other HPAI outbreaks in poultry caused by H7 AIVs [Reference Alexander1, Reference Alexander4, Reference Brown5]. In addition, outbreaks of LPAI due to H5 and H7 LPAIVs continue to occur in poultry, often insidiously. Furthermore, non-NAIVs may elicit mild clinical signs in poultry. These include outbreaks due to H1, H4, H6 and H10 subtypes of AIV that have been recorded in recent years [Reference Brown5]. Among the non-NAIVs, viruses of H9 subtype are a concern across many parts of Asia, the Indian subcontinent and the Middle East, and have also been responsible for zoonotic infections [Reference Alexander1, Reference Brown5].
Virus isolation (VI) in embryonated fowls' eggs (EFEs) has been the classical ‘gold standard’ approach to the diagnosis of AI infections [6, 7]. In recent years real-time reverse transcription–polymerase chain reaction (rRT–PCR) has emerged as a highly sensitive and specific tool for AI laboratory diagnosis [Reference Hoffmann8]. Real-time RT–PCR offers rapid high-throughput testing during outbreaks and surveillance, and carefully optimized and validated protocols are considered to be at least as sensitive as VI [Reference Slomka9].
Published robust validations of AI rRT–PCRs have stressed the importance of careful assay design [Reference Hoffmann8, Reference Slomka9], thorough optimization, which includes a demonstration of rRT–PCR efficiency [Reference McPherson and Møller10], together with sensitivity and specificity assessments using AI laboratory isolates of all 16 H subtypes, as well as well-characterized AI clinical specimens [Reference Spackman11–Reference Monne15]. However, all these published rRT–PCR test validations have focused on testing single AI clinical specimens, and did not include the testing of pooled clinical specimens. The application of AI rRT–PCRs was described in detail for the investigation of two NAI poultry outbreaks that occurred in the UK during 2007 [Reference Slomka9]. During these two outbreaks, a total of about 9300 and 20 300 individual field samples were tested by AI rRT–PCR assay during the 4- and 5-wk duration of the H7N2 LPAI (May–June) and H5N1 HPAI (November) outbreaks, respectively [Reference Slomka9].
Because of the high sensitivity and specificity of optimized AI rRT–PCR methods they should be able to detect a single positive swab pooled with negative swabs as described in this study, but no investigations of pooling for AI rRT–PCR have been described to date. Pooling of poultry swabs would increase testing throughput during outbreak investigations, provided that the sensitivity and specificity of the AI rRT–PCR is equivalent to that observed during the testing of single swabs. Swab pooling may also provide an additional cost benefit in reducing the number of necessary AI rRT–PCR tests since there is a greater likelihood of having at least one positive bird included in a pooled sample, especially at low prevalence, compared to a sample from an individual bird.
However, if there is a reduction in sensitivity when positive samples are mixed with negative samples, then pooled sampling may be less sensitive, and may miss low prevalence infection. A dilution effect has been found in several cases where bacteriological detection by growth of cultures has been performed on pooled samples, e.g. Salmonella in poultry and pigs [Reference Arnold, Cook and Davies16–Reference Arnold, Carrique-Mas and Davies18] and E. coli O157 in cattle [Reference Arnold19]. In these studies, maximum-likelihood or Bayesian methods have been used to compare the likelihood of a pooled sample testing positive vs. the number of positive samples in the pool, and have found a decreasing likelihood of a sample testing positive as the number of positive samples in the pool decreases. Such a dilution effect due to pooling was not unexpected in these previous studies. For testing by rRT–PCR, pooling of swabs may introduce other compromising factors such as the likelihood of the presence of PCR inhibitors. Therefore, the purpose of this study was to determine whether pooling AI-positive swabs with negative swabs had a detrimental effect on the sensitivity of AI rRT–PCRs. Because AI poultry infections are diagnosed at flock level a further aim of this study was to compare the relative efficiency of pooling vs. sampling of individual swabs in identifying an infected flock.
METHODS
Virus, experimental infections of birds and swabs
A/goose/England/07(H2N2) was isolated from a poultry farm during June 2007 in the UK, and was propagated by inoculation into 9- to 11-day-old EFEs to prepare the viral inoculum by standard methods [6, 7], with viral titre expressed as the median egg infectious dose per millilitre (EID50/ml) [Reference Reed and Muench20]. Twelve 3-week-old turkeys were each infected via the oculo-nasal route with 1×106 EID50 in 200 μl inoculum. Swabs [Urethral ENT, supplied by the Medical Wire and Equipment (MW and E) Company, UK] were obtained from the buccal and cloacal cavities of each turkey. These were collected prior to infection and recorded as 0 days post-infection (dpi), then daily from 1 dpi until 18 dpi. In addition, buccal and cloacal swabs were collected from 308 turkeys from healthy flocks farmed in the Irish Republic.
Turkey swabs: individual and pooled
Twenty buccal and 20 cloacal swabs from the healthy farmed turkeys were selected at random and expressed into 1 ml virus transport medium (VTM), which consisted of brain heart infusion broth and antibiotics (BHIB). These 40 swabs were tested by M gene rRT–PCRs to demonstrate the AI-free status of the healthy turkeys that were sampled in the Irish Republic. One hundred and eighty-four swabs were obtained from six experimentally infected turkeys with each bird sampled each day during 1–18 dpi, except one bird that was culled after it provided swabs at 1 and 2 dpi. This gave 92 buccal and 92 cloacal swabs which were similarly expressed into 1 ml BHIB. These 184 swabs were to be individually extracted for RNA and subsequent M gene rRT–PCR testing. One hundred and eight swabs from the remaining six directly infected turkeys (54 buccal, 54 cloacal) collected at 2–11 dpi were selected for pooling, whereby one of these swabs was pooled with four swabs obtained from the corresponding anatomical site from the healthy farmed turkeys. The study was designed to model a worst-case scenario, where one weak AI-positive swab is pooled together with four AI-negative swabs. Each of these 144 pools of five swabs was expressed into 1 ml BHIB.
RNA extraction and AI rRT–PCRs
Swab fluids (140 μl each) from single or pooled swabs were extracted for RNA using the Viral RNA Extraction kit (Qiagen, UK) adapted to the BioRobot 2000 (Qiagen) as described previously, which included positive AIV extraction controls [Reference Slomka14]. Two variants of the M gene rRT–PCR validated test for generic detection of global AIVs of all H subtypes [Reference Spackman11] were used to test extracted RNA. The first assay was performed according to the recommendation in the EU AI Diagnostic Manual [6], as described previously [Reference Slomka14], this will be referred to as the ‘wet’ M gene rRT–PCR. The second was a commercially available version of the M gene rRT–PCR [Reference Spackman11], which is prepared as a freeze-dried reagent with the probe shortened and modified to include an MGB quencher [Reference Petrauskene21], this will be referred to as the ‘bead’ M gene rRT–PCR. The bead M gene rRT–PCR also includes an internal positive control (IPC), which can reveal the presence of PCR inhibitors [Reference Petrauskene21]. RNA was extracted from the quantified preparation of A/goose/England/07(H2N2) and used to construct a tenfold dilution series of RNA, which was tested by both M gene rRT–PCRs in order to demonstrate acceptable efficiencies, i.e. in the range 90–110% to reflect optimal test performance whereby data may be interpreted quantitatively [Reference McPherson and Møller10]. A positive cut-off was used at a cycle threshold (Ct) of 36 [Reference Perkins and Swayne22], for each assay. All M gene rRT–PCR tests were performed using Mx3000 (Stratagene, USA) platforms that were identically calibrated and strictly maintained by the laboratory Quality Assurance programme to operate consistently to the same standard. Fluorescence outputs from M gene quantification were converted to Ct values by using the supplied MxPro software (Stratagene), with amplification plots also inspected visually to ensure uniformity in the generation of Ct values.
Serology
Sera were collected from turkeys before and after (18 and 21 days) infection with the H2N2 virus. These were tested for AI antibodies using two commercially available competitive ELISAs, namely the ID Screen® Influenza A Antibody Competition ELISA (ID Vet, France) and AI MultiS-Screen Ab Test (IDEXX, USA) according to the manufacturers' instructions.
Statistical analysis
We aimed to explore whether pooling of swabs affected the Ct value of M gene rRT–PCRs. If the introduction of inhibitors or other detrimental factors due to pooling were to increase the Ct value, then there would potentially be a reduction in the sensitivity of testing pools compared to testing individual swabs. Specifically, we aimed to determine whether the Ct value of a pooled sample was significantly greater than that of an individual sample.
A secondary aim was to compare the power of flock-level detection for a given number of samples of pooled and individual sampling, and it was therefore necessary to determine the sensitivity of pooled and individual sampling relative to the number of days that a bird had been infected. Therefore, we determined whether there was any temporal pattern to the Ct value compared to the number of days post-infection for each swab type (cloacal and buccal). Preliminary analysis of the data using linear regression of the Ct scores from infected turkeys indicated a statistically significant increase in Ct scores with the number of days since infection (P<0·001) (buccal swabs from 1 dpi and cloacal from 4 dpi, with R 2 values of 0·74, 0·55, 0·81 and 0·74 for the ‘wet’ buccal, ‘wet’ cloacal, ‘bead’ buccal and ‘bead’ cloacal, respectively) and therefore a model of the following form was fitted for each swab type
 $$\mu _{\rm b} \lpar t\rpar \equals \alpha _{\rm b} \plus \beta _{\rm b} t\comma$$
$$\mu _{\rm b} \lpar t\rpar \equals \alpha _{\rm b} \plus \beta _{\rm b} t\comma$$for t>latent (non-shedding) period, where μb represents the mean Ct value of a buccal swab t days after infection, and where αb, βb are unknown parameters estimated from the linear regression. The subscript b is used to represent buccal swabs, and the same formula applied to cloacal swabs, with subscript c in place of subscript b throughout.
The impact on the Ct value of the pooled swabs (one positive diluted with four negative samples) compared to the single swabs was estimated by fitting a model of the following form for the buccal pools
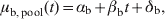 $$\mu _{{\rm b\comma pool}} \lpar t\rpar \equals \alpha _{\rm b} \plus \beta _{\rm b} t \plus \delta _{\rm b} \comma$$
$$\mu _{{\rm b\comma pool}} \lpar t\rpar \equals \alpha _{\rm b} \plus \beta _{\rm b} t \plus \delta _{\rm b} \comma$$The aim of the analysis was then to determine whether δi >0 (i=b, c), which would imply a significant impact on the Ct value of pooling, and thus an effect on the sensitivity of pools compared to individual samples.
There were two possible outcomes for the swab result of each bird at time t; either it would have a Ct value, which was assumed to follow a normal distribution with mean μi and standard deviation σi or have too little viral RNA to be detected and thus have no Ct value at all. For the latter, a logistic regression model was fitted to the number of samples having a Ct score at each time point, assuming that the number of samples with a Ct score followed a binomial distribution with n equal to the number of samples and p such that:
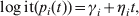 $$\log {\rm it} \lpar p_{i} \lpar t\rpar \rpar \equals \gamma _{i} \plus \eta _{i} t\comma$$
$$\log {\rm it} \lpar p_{i} \lpar t\rpar \rpar \equals \gamma _{i} \plus \eta _{i} t\comma$$where γi,ηi(i=b, c) are unknown parameters determining the likelihood of a swab having a Ct score vs. the time since infection. The expected sensitivity of rRT–PCR applied to an individual swab at t dpi is then given by the product of (i) the probability that the swab has a Ct value [(given by p i(t)] and (ii) the probability that, given it has a Ct value, the Ct value is below the positive cut-off of 36 [Reference Spackman11, Reference Slomka23] (i.e. the probability that μi(t)<36, which is evaluated using the cumulative distribution function of the normal distribution.
The model was fitted in WinBUGS 3.1, using a burn-in of 1000 iterations and a run of 5000 iterations to obtain model results. Three Markov Chains were run with random initial values and the Gelman–Rubin statistic used to assess convergence. WinBUGS 3.1 uses Monte Carlo Markov Chain methods to obtain the probability density of each parameter which is a combination of prior assumptions of the parameter and the likelihood of the parameter from the data, i.e. prior knowledge of each parameter can be incorporated into the analysis and means that the final estimate is a weighted average of both. In this case, we made no strong assumptions about each parameter and used relatively non-informative priors, via normal distributions with mean 0 and variance of 1000. However, the method was still used since it provided very useful indicators of the uncertainty in each parameter.
The Deviance Information Criterion (DIC) [Reference Spiegelhalter24] was used [a Bayesian equivalent of Akaike's Information Criterion (AIC), which compares non-nested models on the basis of their likelihood values, with a penalty for having more parameters]. The DIC was used to determine whether there was any significant difference in the inhibitory effect on the Ct value of pooling between (i) the two different assay types used to test the swabs (wet and bead M gene rRT–PCRs), and (ii) the two different sample types, buccal and cloacal swabs. Model comparisons were made using DIC [Reference Spiegelhalter24], which is a Bayesian analogue of AIC. To assist in the interpretation of the DIC, a DIC weight (w DIC) was calculated for each model being compared, which gives an estimate of the probability that each model is the best model for the data at hand, and is calculated according to
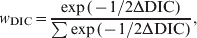 $$w_{{\rm DIC}} \equals {{\exp \lpar {{ \minus 1} \mathord{\left/ {\vphantom {{ \minus 1} {2{\rm \rmDelta DIC}}}} \right. \kern-\nulldelimiterspace} {2{\rm \rmDelta DIC}}}\rpar } \over {\sum {\exp \lpar \minus {1 \mathord{\left/ {\vphantom {1 {2{\rm \rmDelta DIC}}}} \right. \kern-\nulldelimiterspace} {2{\rm \rmDelta DIC}}}\rpar } }}\comma$$
$$w_{{\rm DIC}} \equals {{\exp \lpar {{ \minus 1} \mathord{\left/ {\vphantom {{ \minus 1} {2{\rm \rmDelta DIC}}}} \right. \kern-\nulldelimiterspace} {2{\rm \rmDelta DIC}}}\rpar } \over {\sum {\exp \lpar \minus {1 \mathord{\left/ {\vphantom {1 {2{\rm \rmDelta DIC}}}} \right. \kern-\nulldelimiterspace} {2{\rm \rmDelta DIC}}}\rpar } }}\comma$$where ΔDIC was the difference between the model in question and the minimum value of the DIC for the models being compared, and the denominator was the sum of the differences over all the models being compared.
The outputs of the Bayesian model were used to infer the likelihood of detection of an infected flock by each swab type vs. the number of days since infection. In turn, this likelihood of detection at bird level was then extrapolated to flock level by employing a deterministic differential equation that predicted the prevalence of infection each day for a number of days post-infection. In order to account for the reducing probability of detection as the time since infection increased for each bird, 17 different infectious classes were used, each representing the proportion of birds that had been infected for i=2, …, 18 days (i=1 corresponds to the latent period when birds were not infectious).
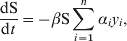 $${{{\rm dS}} \over {{\rm d}t}} \equals \minus \beta {\rm S}\sum\limits_{i \equals \setnum{1}}^{n} {\alpha _{i} y_{i} } \comma$$
$${{{\rm dS}} \over {{\rm d}t}} \equals \minus \beta {\rm S}\sum\limits_{i \equals \setnum{1}}^{n} {\alpha _{i} y_{i} } \comma$$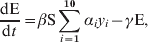 $${{{\rm dE}} \over {{\rm d}t}} \equals \beta {\rm S}\sum\limits_{i \equals \setnum{1}}^{\setnum{10}} {\alpha _{i} y_{i} } \minus \gamma {\rm E\comma }$$
$${{{\rm dE}} \over {{\rm d}t}} \equals \beta {\rm S}\sum\limits_{i \equals \setnum{1}}^{\setnum{10}} {\alpha _{i} y_{i} } \minus \gamma {\rm E\comma }$$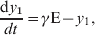 $${{{\rm d}y_{\setnum{1}} } \over {dt}} \equals \gamma {\rm E} \minus y_{1}\comma$$
$${{{\rm d}y_{\setnum{1}} } \over {dt}} \equals \gamma {\rm E} \minus y_{1}\comma$$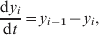 $${{{\rm d}y_{i} } \over {{\rm d}t}} \equals y_{i \minus \setnum{1}} \minus y_{i} \comma$$
$${{{\rm d}y_{i} } \over {{\rm d}t}} \equals y_{i \minus \setnum{1}} \minus y_{i} \comma$$where S represents susceptible birds, E represents exposed birds (infected but not infectious), and y i (i=1, …, n) represents infectious birds having been infectious for i days. The relative infectiousness of each infectious bird was given by αi, which was given by the likelihood of virus excretion, having been infectious for i days, which was assumed equal to the probability that the bird would be detected by rRT–PCR. The transmission rate β was fixed such that R 0 was given by 5·5, equal to the mean of that found in LPAI outbreaks in turkey flocks in Italy [Reference Comin25], and was thus set at 5·5/14·63=0·38, where 14·63 is the mean infectious period. The latent period was set to 1 day, since rRT–PCR detection occurred the day after infection in the experimentally infected birds (the longer latent period of 4 days for cloacal shedding was accounted for in the probability of detection). The main findings from the transmission model in terms of the relative performance of pooled vs. individual samples are not sensitive to the choice of transmission parameters (results not shown).
RESULTS
Establishment of AI negativity in healthy farmed turkeys
Twenty buccal and 20 cloacal swabs were selected at random from the 480 swabs that had been collected from the healthy farmed turkeys. RNA was extracted robotically and tested by both the ‘wet’ and ‘bead’ M gene rRT–PCRs. All 40 swabs registered a ‘no Ct’ result by both M gene rRT–PCRs. This was in accord with EU guidelines to test a minimum of 20 birds for AI from each suspect epidemiological unit [6].
Viral shedding and serology in turkeys directly infected with H2N2
The fit of the linear regression model for the mean Ct value vs. the number of days since infection is shown for buccal swabs (Fig. 1a) and cloacal swabs (Fig. 1b), where the mean Ct was calculated for up to six individual samples/pools per day for each swab type (depending on how many samples had a Ct value on that day; swabs that registered ‘no Ct’ were omitted from the calculation). Cloacal shedding commenced later than buccal shedding (3–4 dpi compared to 1 dpi; Table 1) and after 4 dpi, had a higher mean than buccal shedding (Fig. 1, Table 1), and also had greater variability, especially for the individual samples. The IPC included in the bead M gene rRT–PCR registered only one ‘no Ct’ result among all 328 tested swabs (single and pooled). This one instance of clear inhibition occurred in one pool of cloacal swabs where the cloacal swab from one experimentally infected turkey was obtained at 2 dpi, and the M gene target also registered ‘no Ct’ by both wet and bead assays. All the other cloacal swabs (single and pooled) obtained from the other 11 infected turkeys at 2 dpi similarly registered ‘no Ct’ for the M gene target in the bead assay, and this strongly inferred absence of cloacal shedding at this early time point in any of the birds (Fig 1, Table 1). The IPC successfully registered Ct values in the range 27·63–30·24 [mean 29·13, median 29·13, mode 28·75 (n=8)] for the remaining 415 bead M gene tests (Supplementary Fig. S1). Antibody testing of sera drawn at 18 and 21 dpi with the two commercial AI antibody detection ELISAs revealed AI seroconversion in all infected turkeys (data not shown).

Fig. 1. The mean Ct value (excluding data points where ‘no Ct’ was recorded) of the individual and pooled (a) buccal and (b) cloacal swab samples vs. days since infection of H2N2 LPAI in turkeys for PCR detection using wet and bead M gene assays.
Table 1. Mean cycle threshold (Ct) values for individual swabs and pooled swabs (one positive swab diluted with four negative swabs) from turkeys experimentally infected with H2N2 low pathogenicity avian influenza

—, Indicates that no birds were tested.
‘No Ct’ means that none of the birds tested recorded a Ct value.
Results are given as mean (standard deviation).
Values given without a standard deviation are single swabs.
Simple determination of the extent of analyte loss due to pooling
The volume of four 1-ml aliquots of BHIB were checked by measurement to the nearest 5 μl prior to the addition of single swabs, and then measured again following expression and withdrawal of the swabs. The mean volume loss due to the introduction and withdrawal of single swabs was calculated as 25 μl per 1 ml BHIB aliquot (Supplementary Table S1). The same measurements were done for four 1 ml BHIB aliquots into which a pool of five swabs were added, expressed and withdrawn. For the pools of five swabs the mean analyte loss was 124 μl per 1 ml BHIB aliquot (Supplementary Table S1). Therefore pooling of five swabs results in an increased BHIB volume loss of ∼100 μl compared to that observed for single swabs.
Statistical approach to determine whether pooling is detrimental to M gene rRT–PCR testing of turkey swabs
Proportion positive by swab and assay type
The bead M gene assay generally had lower Ct values than the wet M gene assay [Fig. 1 and compare estimates of αb, αc (the constants in the linear regression that determine the fitted Ct values) in Tables 2 and 3]. Using the estimates from the linear regression of Ct value and probability of a Ct value vs. days post-infection (Tables 2 and 3, Supplementary Figs S2 and S3), the sensitivity of each swab can be estimated (Fig. 2). This showed a decreasing probability of detection the longer the birds had been infected, and the much higher probability of detection using the bead than the wet M gene assay. For the bead assay there was still a ∼20% chance of detection with each swab type at 18 dpi.

Fig. 2. The estimated sensitivity of buccal and cloacal swabs for detection of H2N2 by wet and bead M gene assays in turkeys vs. the number of days since infection.
Table 2. List of parameters from a Bayesian model applied to individual and pooled buccal and cloacal swab data from turkeys infected with H2N2 for the wet M gene assay

Ct, Cycle threshold.
Table 3. List of parameters from a Bayesian model applied to individual and pooled buccal and cloacal swab data from turkeys infected with H2N2 for the M gene bead assay

Ct, Cycle threshold.
Impact of pooling on Ct value
In terms of the inhibitory effect of pooling by assay type, there was a significant improvement in model fit (according to DIC) in having a different inhibitory effect between the assay types (P<0·05). Furthermore, the comparison of the inhibitory effect between the buccal and cloacal swabs (i.e. δb, δc) indicated no difference between the swab types for the wet M gene assay, but did show an important difference for the bead M gene assay (P<0·01). Therefore in the final model, δb was assumed equal to δc for the wet assay (Table 2) but allowed to differ from δc in the bead assay (Table 3). It was also assumed that δb, δc could differ between assay types.
For the wet M gene assay, there was a small estimated increase in Ct value (δb, δc =0·07, Table 1) from the PCR for the pooled samples compared to the individual samples. However, the credible interval contained zero and thus there was no important inhibitory effect found for the wet assay. For the bead M gene assay, although the median of δc was larger than for the wet M gene assay, there was similarly no important inhibitory effect for the cloacal swabs (Table 3). The pooled buccal swabs tested by the bead M gene assay actually had a significantly lower Ct value than the individual swabs, and were more likely to be positive than the individual swabs (Table 3); this is possibly due to a small number of outlying values for the individual swabs for the bead assay which may have made the mean Ct value for the individual samples higher.
Pooling vs. individual sampling for flock-level detection
Since there was no inhibitory effect of pooling, the likelihood of a bird testing positive by a pool of five buccal swabs was assumed equal to 1 – (1 – θb(i))5, where θb(i) represents the sensitivity that an infected bird is detected by buccal swab rRT–PCR having been infectious for i days. The equivalent formula holds for cloacal swabs with subscript b changed to c.
The lack of an inhibitory effect means that four pools of five would have the same sensitivity to detect infection in a flock as 20 individual cultures, meaning that the same sensitivity could be maintained using pools as by testing individual swabs but with only 20% of the number of rRT–PCR tests.
The use of the bead instead of the wet M gene assay is predicted to increase the window in which the infected flock would be detected by rRT–PCR by ∼5 days (Fig. 3) (although the length of the extension would depend upon the within-flock dynamics, which will vary between virus strains). Increasing the number of pools of five to 20, i.e. equal to the number of tests for individual sampling except 100 birds would be tested, would again increase the length of the window in which the flock would be detected by a further 4 days or so. There was no benefit in increasing the number of pools in the first few days after the flock was infected since using only four pools of five had a very high probability of detecting the flock once excretion of virus had started in infected birds.

Fig. 3. The estimated sensitivity of detection of a LPAI infected turkey flock: (i) sampling 20 birds with both buccal and cloacal swabs, tested with wet M gene assay, (ii) or four pools of buccal swabs and four pools of cloacal swabs, each with five birds per pool, and each tested with bead M gene assay, and (iii) testing 20 pools of five swabs with bead M gene assay.
DISCUSSION
The results of the present study indicate that no factors relating to potential inhibition or indeed other related or undefined factors have resulted in any significant compromise due to pooling of turkey swabs. The successful detection of the IPC in the tests conducted by the bead M gene assay indicated that PCR inhibition was not a noticeable factor in the testing of all RNA extracts from all the swabs (individual and pooled) in this study. This is consistent with absence of rRT–PCR inhibitors in extracted RNA from chicken oro-pharyngeal and cloacal swabs reported previously, although inhibition was observed for wild bird cloacal swabs [Reference Das26]. These findings were determined using swabs collected from 12 turkeys experimentally infected with a LPAI virus, A/goose/England/07(H2N2). Earlier attempts to compare the rRT–PCR testing of individual and pooled swabs by using artificially reconstructed swabs produced inconsistent results (data not shown), hence it was considered important to design a comparison based on in vivo generated swabs. Another key aspect of this study was that all the turkeys were successfully infected with this H2N2 isolate, as demonstrated by seroconversion and sustained daily viral shedding (both buccal and cloacal) observed by testing of the swabs by M gene rRT–PCRs. In addition, the quantity of viral RNA that corresponded to the shedding titre was considered to be typical of that observed in swabs obtained in LPAI field infections, i.e. with Ct values >30 [Reference Slomka9] at the majority of sampling time points (Fig. 1). This consistent, low level of shedding provided assurance that the swabs taken for pooling were of low titre. Therefore pooling of one weak positive swab with four negative swabs reconstructed a scenario which may be encountered in the field as a worst case for successful detection, i.e. when the prevalence of AI positives in a flock may be low, and the sampled positive swabs are at low viral load.
Gallinaceous poultry flocks that are infected with HPAI display rapid progression to clinical disease and mortality within 48–72 h of introduction of infection, as has been regularly observed in experimental in vivo studies [Reference Slomka14, Reference Perkins and Swayne22]. At the time of clearly apparent disease and death, a high proportion of birds are known to be infected and shedding at high viral loads, e.g. Ct 20–25 in AI rRT–PCR tests. Such high levels of HPAI shedding have been observed also in field-infected galliformes, with high infection prevalences having been recorded in diseased galliforme flocks [Reference Slomka9, Reference Slomka23]. The testing of pools in such a high viral load scenario should certainly not compromise detection of HPAIV infection in diseased galliformes. The domestic duck is an important host for H5N1 HPAI in many Asian countries, in which clinical signs may not always be apparent [Reference Kim27], hence surveillance testing of this species for infection is critical to prevent the emergence of new outbreaks. Domestic ducks infected with H5N1 HPAIVs may shed virus at a range of titres from the oropharyngeal and cloacal sites [Reference Kim28, Reference Sturm-Ramirez29], although field investigations in Vietnam showed the titre of duck shedding to be lower than that observed in chickens [Reference Perkins and Swayne22]. However, the present findings would suggest that pooling of duck swabs for AI rRT–PCR testing would be a legitimate approach for surveillance and outbreak investigations where domestic duck flocks are suspected of being infected. Since there could be differences between shedding patterns for different host/strain combinations, it would be valuable to extend such studies to different combinations of poultry species and other LPAIV isolates for further verification of pooling.
It is important to emphasize how the pools were prepared, i.e. they were collected and pooled in a dry state prior to expression of all five swabs together into 1 ml VTM, and a portion of this was extracted to prepare RNA for testing by an optimized and validated AI rRT–PCR. The effect on the loss of target analyte due to pooling appears to be small, whereby the effect of adding, expressing and withdrawing a pool of five swabs resulted in a volume loss that was 100 μl greater than that observed for a single swab (Supplementary Table S1). This is a volume loss relative to the starting BHIB volume of 1 ml, but may be considered to be proportionately small. Therefore the described pooling approach should not result in any significant dilution or other analyte loss effects, particularly as the total volume of the five swabs is small compared to the volume of VTM into which they were expressed.
Existing recommendations pertaining to the pooling of clinical specimens have been put into practice for the pooling of tissues obtained from any available carcasses [6, 7], and in the case of diseased galliformes infected with H5N1 HPAIV, the high viral titres resulting from the systemic spread of such infections have resulted in successful AI rRT–PCR detection [Reference Slomka9]. Pooling of swabs for VI testing has been put into practice in the context of liquid pools, i.e. where individual swabs are first expressed into 1 ml volumes of VTM, and five equal volumes are pooled from five individual VTMs, all obtained from the same anatomical site and from the same species in the same epidemiological unit. Portions of each liquid pool were then inoculated into EFEs [Reference Slomka9] and sufficient volumes are necessary to inoculate into four or five EFEs and for any repeat VI testing. However, it is clear that a fivefold dilution effect is a consequence of this liquid pooling approach if, for example, one positive swab is pooled with four negatives, and in the case of a low-titre positive swab this may compromise the success of VI. Therefore liquid pooling requires caution. An example of this was observed in the investigation of an outbreak of H9N1 LPAI in UK chickens in late 2010 (C. D. Parker, unpublished observations). Sensitive molecular approaches were critical in successfully characterizing the infecting H9N1 subtype because all attempts to isolate viable H9N1 virus by a liquid pooling approach from this outbreak had failed. This may have related to the very low titres of the chicken swabs that were reflected by high Ct values for the individual swabs, and the liquid pooling approach could have further diluted any stronger swabs that may have yielded a positive VI result if tested individually. It is important to emphasize that the current study differed in that five swabs were pooled into 1 ml VTM, so no obvious dilution factor should have caused any compromise in AI detection by rRT–PCR. Although this was not modelled in the present study, it is possible that a beneficial concentration effect may result in cases where two or more positive swabs are included in a pool of five.
This study included testing pooled swabs by optimized and validated M gene rRT–PCRs, and the original validation studies all included detection of the target analyte in clinical specimens [Reference Slomka9, Reference Spackman11, Reference Lee and Suarez12, Reference Petrauskene21]. Validation of these tests also included demonstration of very high specificity, so that clinical specimens that were VI negative but rRT–PCR positive could be confidently accepted as genuine AIV positives [Reference Slomka9, Reference Petrauskene21]. Therefore the high sensitivity and specificity of these rRT–PCRs cannot be undervalued, and may well have been an attribute in the current study which has shown them to be successful in the detection of AI in pooled clinical specimens.
According to the EU AI Diagnostic Manual [6], the sampling design for the collection of swabs from 20 birds per epidemiological unit is based on the detection of at least one positive bird with 95% confidence if prevalence is ⩾5%. In the course of outbreak-related surveillance in the UK, up to 60 birds per epidemiological unit are swabbed in order to obtain greater statistical certainty, particularly as the prevalence of infection may be low. This was observed at one ‘dangerous contact’ premises that was tested during the November 2007 H5N1 HPAI outbreak in free-range turkeys in the UK, where H5 rRT–PCR revealed three infected birds among the 60 which were sampled [Reference Slomka9]. Although the testing of pooled swabs appears to be valid even at early time points following introduction into a flock when infection may be at a low prevalence and viral shedding at low titres (Fig. 3), it remains prudent to carefully consider any results from any flocks that may register only indeterminate AI rRT–PCR results in the Ct range 36·01–39·99. This would necessitate a request for repeat swabbing of the suspect flock, when the prevalence of infection and viral shedding should be higher.
Results from the Bayesian analysis in this study also revealed that the bead M gene rRT–PCR was more sensitive than the wet M gene rRT–PCR for both single and pooled swabs. The bead M gene rRT–PCR has demonstrated advantages in the testing of clades 2.3.2 and 2.3.4 H5N1 HPAI viruses encountered in poultry outbreaks in Vietnam during 2009, where data analysis which used a Bayesian model, also revealed a superior sensitivity for bead compared to wet M gene rRT–PCRs [Reference Slomka23]. It must be emphasized, however, that the compromised sensitivity of the wet M gene rRT–PCR in testing the 2009 Vietnamese clinical specimens was due to three nucleotide mismatches in the reverse primer sequence. The same mismatches also occurred in the bead M gene rRT–PCR, but it would appear that differences in the core chemistries and thermocyling conditions in the two M gene assays [Reference Slomka14, Reference Petrauskene21] restored sensitivity of detection by the bead M gene rRT–PCR [Reference Slomka23].
The results indicate a potentially extended period after infection for which virological sampling could detect H2N2 excretion in individual birds (Fig. 2). This is because of a few birds that have intermittent excretion of virus up to 18 dpi (and potentially beyond). These birds had relatively high Ct values at >2 weeks after infection, close to the positive cut-off at Ct 36, and would not have been detected by the wet M gene assay, suggesting that the use of a more sensitive test could increase the window in which virological sampling would detect infection in a flock. Furthermore, it is important for determining the effectiveness of virological sampling during an outbreak, to be able to determine the window in which virological sampling could detect infected birds. This study indicates that the distribution of the infectious period is determined from experimental and field data, rather than a point value, since point values for the infectious period are likely to provide poor predictions of the period of flock infectiousness; a few birds with long infectious periods could make an important difference to the likelihood of detection in the tail of an outbreak.
To progress this work further, it would be useful to be able to relate the sensitivity of the rRT–PCR tests and the Ct scores to viral shedding. This would enable the sensitivity of both individual and pooled samples to be predicted over the time course of infection within an individual bird, and if transmission rates were known, over a flock. Such a model would provide testable predictions that could be verified by within-flock transmission experiments.
SUPPLEMENTARY MATERIAL
For supplementary material accompanying this paper visit http://dx.doi.org/10.1017/S0950268812001811.
ACKNOWLEDGEMENTS
The authors wish to thank the UK Department of the Environment, Food and Rural Affairs for funding the AI Reference Laboratory at AHVLA-Weybridge, which supported this work under grant SV3400. Dr Dennis Alexander's comments on the manuscript are also acknowledged.
DECLARATION OF INTEREST
None.








