



Introduction
The Caatinga is the largest and most diverse seasonally dry tropical forest (SDTF) of the New World (Silva et al. Reference Silva, Leal and Tabarelli2017). Located in the semiarid region of northeastern Brazil, it presents many extreme environmental and meteorological parameters: high annual temperature and solar radiation, low relative humidity, irregular rain, and extended droughts (Prado Reference Prado, Leal, Tabarelli and da Silva2003).
The vegetation in the Caatinga has a wide variation across space and time and strongly correlates with rainfall; hence, most plants produce flowers and fruits during the short-wet season (Silva et al. Reference Silva, Leal and Tabarelli2017). In general, caatinga plant communities occur in mosaics of open xeric spiny shrublands, clusters of diverse cacti and succulents, tall dry forest stands, and vegetation associated with water bodies and rocky outcrops (Leal et al. Reference Leal, Tabarelli, Da Silva and Silva2003). Thus, the Caatinga has been classified and divided into several phytophysiognomies and vegetational units (see Velloso et al. Reference Velloso, Sampaio and Pareyn2002).
Bats comprehend almost half of the Caatinga mammal species, with records of 96 species and two endemics (Carmignotto and Astúa Reference Carmignotto, Astúa, da Silva, Leal and Tabarelli2017, Gutiérrez and Marinho-Filho Reference Gutiérrez and Marinho-Filho2017, Silva et al. Reference Silva, Delgado-Jaramillo, Souza Aguiar and Bernard2018). As in most Neotropical ecosystems, bat assemblages in the Caatinga contain members of all mammalian trophic guilds (frugivory, nectarivory, piscivory, carnivory, insectivory, and sanguinivory) (Paglia et al. Reference Paglia, Fonseca, Rylands, Herrmann, Aguiar, Chiarello, Leite, Costa, Siciliano, Kierulff, Tavares, Mittermeier and Patton2012). However, bat richness, guilds, and species composition can vary depending on the type of phytophysiognomies – or foraging habitat (Carvalho-Neto et al. Reference Carvalho-Neto, da Silva, Santos, Rohde, Garcia and Montes2016, Gregorin et al. Reference Gregorin, Carmignotto and Percequillo2008, Sá-Neto and Marinho-Filho Reference Sá-Neto and Marinho-Filho2013, Willig Reference Willig1983, Silva et al. Reference Silva, Delgado-Jaramillo, Souza Aguiar and Bernard2018).
Frugivorous bats are very diverse in the Caatinga, occurring from forested to disturbed areas (Mares et al. Reference Mares, Willig, Streilein and Lacher1981, Willig Reference Willig1983, Silva Reference Silva2007, Gregorin et al. Reference Gregorin, Carmignotto and Percequillo2008, Sá-Neto and Marinho-Filho Reference Sá-Neto and Marinho-Filho2013, Novaes and Laurindo Reference Novaes and Laurindo2014, Rocha et al. Reference Rocha, Ruiz-Esparza and Ferrari2018, Soares et al. Reference Soares, Rocha, Bocchiglieri and Ferrari2019), while nectarivores are also abundant, despite the low richness in certain localities (Willig Reference Willig1983, Sá-Neto and Marinho-Filho Reference Sá-Neto and Marinho-Filho2013, Novaes and Laurindo Reference Novaes and Laurindo2014, Cordero-Schmidt et al. Reference Cordero-Schmidt, Barbier, Vargas-Mena, Oliveira, Santos, Medellín, Rodríguez-Herrera and Venticinque2017, Rocha et al. Reference Rocha, Ruiz-Esparza and Ferrari2018, Soares et al. Reference Soares, Rocha, Bocchiglieri and Ferrari2019). Gleaning insectivorous and carnivorous bats, referred to as animalivorous, are rich in species but found in low abundances and associated with more forested areas (Mares et al. Reference Mares, Willig, Streilein and Lacher1981, Silva Reference Silva2007, Beltrâo et al. Reference Beltrão, Zeppelini, Fracasso and Lopez2015, Sá-Neto and Marinho-Filho Reference Sá-Neto and Marinho-Filho2013, Nogueira et al. Reference Nogueira, Pol, Pessôa, Oliveira and Peracchi2015, Silva et al. Reference Silva, Dias, Martins, Guedes, de Almeida, da Cruz, Serra-Freire, Damascena and Peracchi2015). Sanguinivorous bats are common in disturbed areas (Gregorin et al. Reference Gregorin, Carmignotto and Percequillo2008, Nogueira et al. Reference Nogueira, Pol, Pessôa, Oliveira and Peracchi2015, Rocha et al. Reference Rocha, Ruiz-Esparza and Ferrari2018, Soares et al. Reference Soares, Rocha, Bocchiglieri and Ferrari2019) and relatively less common in preserved caatinga forests (Vargas-Mena et al. Reference Vargas-Mena, Cordero-Schmidt, Bento, Rodríguez-Herrera, Medellín and Venticinque2018a, Vargas-Mena et al. Reference Vargas-Mena, Cordero-Schmidt, Rodríguez-Herrera, Medellín, Bento and Venticinque2020). As for open-space insectivorous bats, 50% of Caatingas’ bats belong to this feeding guild but reportedly uncommon due to sampling bias regarding the wide use of mist nets – these bats can detect and avoid nets easily (Gregorin et al. Reference Gregorin, Carmignotto and Percequillo2008, Novaes and Laurindo Reference Novaes and Laurindo2014, Beltrão et al. Reference Beltrão, Zeppelini, Fracasso and Lopez2015, Silva and Bernard Reference Silva and Bernard2017, Soares et al. Reference Soares, Rocha, Bocchiglieri and Ferrari2019). However, captures near water bodies and riparian vegetation can present a relatively high richness and abundance of insectivorous bats (Silva Reference Silva2007, Beltrão et al. Reference Beltrão, Zeppelini, Fracasso and Lopez2015, Novaes et al. Reference Novaes, Laurindo and Souza2015).
Only about 10% of the Caatinga has been adequately sampled regarding bats (see Bernard et al. Reference Bernard, Aguiar and Machado2011, Carmignotto and Astúa Reference Carmignotto, Astúa, da Silva, Leal and Tabarelli2017). Thus, regions like the northeast of the Caatinga, where the state of Rio Grande do Norte is located, require more bat inventories and monitoring (Bernard et al. Reference Bernard, Aguiar and Machado2011, Carmignotto and Astúa Reference Carmignotto, Astúa, da Silva, Leal and Tabarelli2017, Vargas-Mena et al. Reference Vargas-Mena, Alves-Pereira, Barros, Barbier, Cordero-Schmidt, Lima, Rodríguez-Herrera and Venticinque2018b). A recent effort to fill this data gap found that nearly 70% (32 species) of the state’s bat diversity occurs in the Caatinga dry forest of the state (Vargas-Mena et al. Reference Vargas-Mena, Alves-Pereira, Barros, Barbier, Cordero-Schmidt, Lima, Rodríguez-Herrera and Venticinque2018b). Nevertheless, data on the species distribution and community structure of bats are scarce in this region of the Caatinga.
Gathering baseline data about diversity patterns, trophic guilds, and species composition can provide a novel insight into the structure of bat assemblages in the northeastermost region of the Caatinga. Such data can also pinpoint important areas or habitats that may be vital for the conservation of local bat populations. Unfortunately, the Caatinga has been profoundly altered in the last decades by the conversion of areas of native vegetation into human-made ecosystems, chronic disturbance by overexploitation of native species, and introduction of exotic species of plants and animals. Consequently, areas with tall forest stands that were once the most dominant physiognomy today are mostly occupied by open physiognomies and secondary forests (Silva et al. Reference Silva, Leal and Tabarelli2017).
In this study, our objective was to describe the assemblage structure of bats by analysing the richness, abundance, trophic guild representation, and species composition in different caatinga habitats in six sites in the northeastermost region of the Caatinga dry forest. We expect that the structure of bat assemblages and the species distribution will differ according to the habitat type. We look forward that this study will help better understand the structure of Caatinga’s bat assemblages and its species composition, bringing valuable contributions to future conservation actions of local bat populations and their foraging habitats.
Methodology
Study area
We conducted this study in the northeastermost region of the Caatinga dry forest in northeastern Brazil (Figure 1). Specifically in the state of Rio Grande do Norte, the Caatinga corresponds to 91% of its territory (Serviço Florestal Brasileiro 2020). The region has a semiarid climate (Diniz and Pereira Reference Diniz and Pereira2015) and BShw (hot and dry) Köppen classification (Alvares et al. Reference Alvares, Stape, Sentelhas, Gonçalves and Sparovek2013). The rainy season is short from January to June with an average annual precipitation of 400–1100 mm. The dry season extends up to 8–9 months from June or July to December, with a mean annual precipitation of < 50 mm (Diniz and Pereira Reference Diniz and Pereira2015).

Figure 1. Location map of sites and distribution of habitats that were surveyed for bats in the northeastermost region of the Caatinga dry forest from 2017 to 2019. a) Biogeographical distribution of the Caatinga in northeastern Brazil, and b) geopolitical limits of the state of Rio Grande do Norte where this study was conducted.
We surveyed six sites ≥ 55 km apart (Figure 1) in the state of Rio Grande do Norte and selected three protected and three non-protected areas based on logistics, access, and capture permits.
Surveyed sites were as follows:
Açú National Forest (ANF): it is a protected area with sustainable use of natural resources (IUCN management category IV) of 215.25 ha located in Assú municipality (5º34’S, 36º54’W). It has a plain and smooth topography (altitude: 18–100 m) with a vegetation of 50 years of preservation predominantly of shrubby and secondary arboreal caatinga, Copernicia palm groves are found at the margins of the Piató lake, and a diverse garden/orchard is located at the rangers/visitors’ facilities.
Seridó Ecological Station (SES): it is a strict nature reserve (IUCN management category Ia) of 1123.61 ha (06º95’S, 37º39’W) with a flat topography with some ‘inselbergs’ (isolated hills that stands above well-developed plains). Altitude varies from 200 to 385 m. Water bodies rely on two semi-perennial lakes and one artificial dam near the main facilities, and streams are intermittent. Vegetation is predominantly of shrubby and riparian caatingas, with medium caatinga forest near inselbergs foothills, and plants associated with granitic outcrops.
Furna Feia National Park (FFNP): it is a national park (IUCN management category: II) of 8494 ha located in the municipalities of Mossoró and Baraúna (5º04’S, 37º32’W). Topography is plain and smooth (altitude from 70 to 280 m). Water bodies are scarce, and streams, lagoons, and underground rivers occur intermittently. Large karstic calcareous outcrops are found with records of 206 caves (Bento et al. Reference Bento, Cruz, Santos, Freitas, Campos and Souza2013). The unit is one of the few areas in the state that is home to old-growth caatinga forests, but secondary shrubby caatingas are common in semi-open areas and near rocky outcrops.
Felipe Guerra (FG): it is located in the municipality of Felipe Guerra in the Apodí Plateau (5º35’S, 37º41’W). The relief is smooth (altitude from 30 to 100 m) and the Apodí river flows from south to north, and intermittent streams can be found. Extensive calcareous rocky outcrops are found with records of 496 underground cavities – the largest in the state (Bento et al. Reference Bento, Cruz, Freitas and Campos2015). Vegetation is mainly of shrubby/arboreal and riparian caatingas, plant species associated with rocky outcrops, and extensive palm groves of the endemic Copernicia prunifera at the Apodí river margins.
Martins: it is located in the municipality of Martins (6º05’S, 37º54’W) and has a rocky and mountainous topography with plateaus ranging from 400 to 800 m above sea level. Remnants of montane semideciduous forest are found on top of the plateaus with an orographic rainfall of 1200 mm/year, contrasting with the surrounding Caatinga vegetation at lower altitudes (Porto et al. Reference Porto, Cabral and Tabarelli2004).
Lajes: it is located in the municipality of Lajes (5º42’S, 36º14’W) and is mountainous with a rugged relief and rocky terrain (altitude from 300-600 m). Streams are scarce and intermittent. It harbours one of the state’s largest fragments of arboreal and shrubby caatingas with large clusters of cacti, terrestrial bromeliads, and Leguminosae trees (hereafter, Cacti forests).
Data collection
In each site, we captured bats using mist nets ranging from 3 to 12 m long × 2.5 m high (20 mm mesh size with five shelves) and nets of 14 m long × 3m high (25 mm, four shelves). Depending on the logistics of the site, we selected two to five capture points for a total of 22 capture points throughout the six sites. The location and number of capture points in each site can be found in Supplementary Material 1. In each capture point, we deployed from 150 to 220 m of mist nets, which remained open for 6.5 h from dusk until midnight (17:30–24:00 h). Nets were arranged in sets of three to five nets in zigzag pattern on trails, roads, semi-open areas, and near water bodies. Because the capture data of this study was also used in another study regarding phytophagous bats, we deployed in all sites one set of mist nets near food resources when available (Cacti or trees/bushes with flowers or fruits).
In the six sites, we surveyed 99 nights from February 2017 to June 2019. The number of nights and sampling effort differed in each site due to logistics constraints. However, all sites were surveyed at least twice a year to cover the rainy and dry seasons, except in Lajes, which was sampled almost every month from February 2017 to February 2019 due to easy access and proximity to the state’s capital (Supplementary Material 1).
Each capture point was classified into an habitat type based on those described by Mares et al. (Reference Mares, Willig, Streilein and Lacher1981) and Prado (Reference Prado, Leal, Tabarelli and da Silva2003). We followed these habitat definitions because they represent potential and plausible foraging habitats for bats in the Caatinga. Selected habitats were shrubby caatinga with 26 sampled nights, riparian caatinga with 42, medium caatinga forest with 10, nine nights in rocky outcrops, five nights in Copernicia groves, three nights in humid forest enclave, and two nights in gardens/orchards. For detailed habitat description, see Table 1. Number of sampled nights, capture effort, and specific month surveyed in each habitat can be found in Supplementary Material 2, and habitat photos can be found in Supplementary Material 3.
Table 1. Description of caatinga habitats selected for bat data collection in the Caatinga of Rio Grande do Norte, northeastern Brazil based on Mares et al. (Reference Mares, Willig, Streilein and Lacher1981) and Prado (Reference Prado, Leal, Tabarelli and da Silva2003)

Bat capture and handle protocols followed the guidelines of the American Society of Mammalogists for the use of wild mammals in research (Sikes et al. Reference Sikes2016). Mist nets were not left unattended for > 15 min, and entangled bats were carefully removed immediately to minimise injury and stress. After extraction, bats were held in cotton bags and identified through biometric data and other specific traits using the identification key of South American bats by Díaz et al. (Reference Díaz, Solari, Aguirre, Aguiar and Barquez2016). Before release, all bats were marked in the lower back by cutting the rumps, hair with scissors for recapture control. Specimens with doubtful identification or new occurrences were collected and deposited in the Prof. Adalberto Varela’s Mammal collection of the Universidade do Rio Grande do Norte (UFRN). The Brazilian environmental agency issued collecting permits (SISBIO licence number 48325-2 MMA, IBAMA, and ICMBIO).
Since bats can occupy different trophic levels, classifying species into guilds can help to understand the structure and composition of bat assemblages. Thus, we classified the recorded species into trophic guilds following Hill and Smith (Reference Hill and Smith1984) based on the available natural history information about the primary diet of the bats in Brazil (Reis et al. Reference Reis, Peracchi, Batista, Lima and Pereira2017). Some bat species (e.g., subfamily Phyllostominae) that feed on both insects and vertebrates were classified as gleaning animalivores based on Giannini and Kalko (Reference Giannini and Kalko2005). Therefore, trophic guilds selected were aerial insectivores, gleaning animalivores, piscivores, frugivores, nectarivores, omnivores, and sanguinivores.
We calculated the bat-capture effort of each habitat following Straube and Bianconi (Reference Straube and Bianconi2002), which multiplies the total mist-net area with the number of hours that nets were open during each night. As capture effort differed among habitats, we corrected bat-capture data for capture effort as follows: [captures N/(mist-netting hours * metres2) * 1000]. Capture effort for each habitat and sites can be found in Supplementary Material 2 and 4, respectively.
Data analysis
To evaluate the sampling efficiency of each area and habitat, we used individual-based species accumulation curves using EstimateS Software 9.1.0. The sample order was randomised 1000 times to eliminate the influence of the order in which days were added to the total number of individuals. If computed curves approach an asymptote, the effort has been sufficient to collect most species of an area (Moreno and Halffter Reference Moreno and Halffter2001).
We used rank abundance plots to analyse the species abundance distribution and species evenness/dominance between all seven habitats. The rank abundance plot’s shape can indicate dominance or evenness in a community or sample (Matthews and Whittaker Reference Matthews and Whittaker2015). Steep plots of the set of bars that fits the graph signify assemblages with high dominance, while shallower slopes indicate higher evenness within the assemblages.
To analyse possible association between the habitats and one or more bat species in the assemblage, we ran a correspondence analysis (CA). This analysis simultaneously provides an ordination of the habitats and ordination for the species. It provides a graphical presentation of the direction of the changes in the assemblage structure of each habitat along the axes and the relative importance of each species for the assemblage (Ter Braak Reference Ter Braak, Jongman, Ter Braak and Van Tongeren1995). The CA was done only with habitats containing more than 100 captures and using the species abundance data corrected by the effort. Recaptured individuals were not considered for analysis. All analyses were done using SYSTAT software version 12.
Results
We captured 1575 individuals of 31 species belonging to seven trophic guilds distributed in five families with a net capture effort of 239 665 hm2 (Table 1) (Supplementary Material 5). Phyllostomidae was the family with more species recorded (20 species), followed by Molossidae (5), Noctilionidae (2), Vespertilionidae (2), Mormoopidae (1), and Emballonuridae (1). With the sampling effort corrected, the most abundant captured species corresponded to Artibeus planirostris (258 individuals per 1000 hm2), Glossophaga soricina (46), Desmodus rotundus (45), Lonchophylla mordax (30), and Myotis lavali (20).
The humid forest enclave, riparian, and shrubby caatinga presented adequate sampling effort as accumulation curves nearly reached an asymptote. Other habitats showed steep curves, and greater effort seems to be needed (Figure 2b). The riparian caatinga was the habitat where more species were recorded with 21 species, followed by the shrubby caatinga (17), rocky outcrops (15), humid forest enclaves (11), medium caatinga forests and Copernicia groves with 10 species each, and gardens/orchards with six species. After effort correction, the shrubby caatingas was the habitat that had the highest abundance of bats with 224.3 individuals per 1000 hm2, followed by riparian caatingas with 196.0, rocky outcrops with 113.6., humid forest enclaves with 94.4, orchards/gardens with 35.2, medium caatinga forests with 33.9, and Copernicia groves with 31.1 individuals.
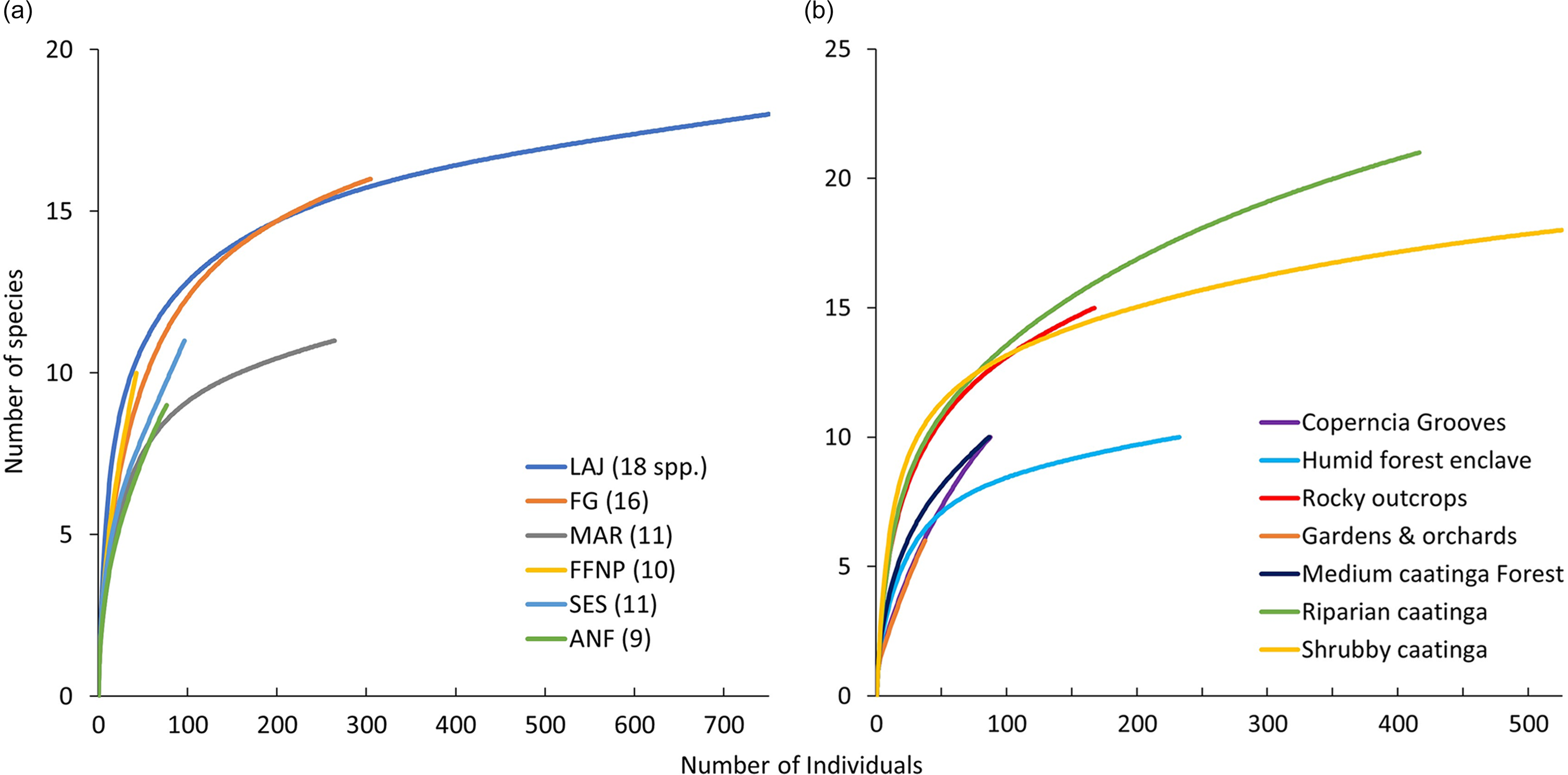
Figure 2. Individual-based species accumulation curves of a) the six surveyed sites: Lajes (LAJ), Felipe Guerra (FG), Martins (MAR), Furna Feia National Park (FFNP), Seridó Ecological Station (SES), Açú National Forest (ANF), and b) of the seven surveyed habitats in the northeastermost region of the Caatinga dry forest (Rio Grande do Norte state) in northeastern Brazil from 2017 to 2019. Curves were randomised 1000 times.
The rank abundance distributions in all habitats presented steep plots with high dominance of phyllostomid bats (Figure 3). However, the species composition differed among all habitats from the second to fifth abundance ranks. Some species were abundant in just one or more habitats, while others were absent or rare. A. planirostris was the most abundant species (rank = 1) and dominated all seven habitats. This species was dominant in the riparian caatinga and humid forest enclaves and presenting a relative superabundance with the other well-sampled habitats. Moreover, this pattern was also observed in the poor-sampled habitats (e.g., Copernicia groves, median caatinga forests, and gardens and orchards). All these habitats presented a high slope variation between abundance bars (Figure 3). However, in the rocky outcrops and shrubby caatinga, the dominance of A. planirostris was relatively lower, and other species presented similar abundances in the subsequent ranks (ranks = 2–5). Of all habitats, the shrubby caatingas presented a relatively more even assemblage than the other habitats, with A. planirostris in the first in rank but with a high abundance of the nectarivores G. soricina, L. mordax, X. vieirai, and L. inexpectata.

Figure 3. Bat species rank abundance distribution in seven surveyed habitats in the northeastermost region of the Caatinga dry forest (Rio Grande do Norte state) in northeastern Brazil from 2017 to 2019. Species acronyms correspond to the top five most abundant species. Acronyms follows in alphabetic order: Artibeus planirostris (A.p.), Carollia perspicillata (C.p.), Desmodus rotundus (D.r.), Glossophaga soricina (G.s.), Lonchophylla inexpectata (L.i.), Lonchophylla mordax (L.m.), Molossus molossus (M.m.), Myotis lavali (M.l.), Noctilio leporinus (N.l.), Peropteryx macrotis (P.m.), Phyllostomus discolor (P.d.), Platyrrhinus lineatus (P.l.) Sturnira lilium (S.l.), Trachops cirrhosis (T.c.), Xeronycteris vieirai (X.v.).
Differences at trophic guild were found among habitats (see Supplementary Material 6b). The shrubby caatingas had the largest proportion of nectarivores (50% of captures) and the habitat with the largest proportion of animalivorous bats (9%). In the riparian caatingas, frugivores (37%) and nectarivores (34%) composed most of the assemblage, followed by insectivores (12%). Frugivores dominated in humid forest enclaves (92%) and in Copernicia groves (81%), piscivores only occurred in the latter (7%). In orchards and gardens, frugivores dominated, followed by insectivores with 16% of captures. Similarly, frugivores dominated (65%) in medium caatinga forests, but sanguinivores were the second to dominate the captures (17%). Lastly, in rocky outcrops, the highest capture proportions were distributed in frugivores (33%), nectarivores (23%), and sanguinivores (23%).
The CA indicated differences in the species composition between habitats (Figure 4). Axis 1 of the CA accounted for 67.8% of the variation, and axis 2 accounted for 21.9%. The CA separated the species and habitats into three groups in relation to both axes. The humid forest enclave was grouped in the negative values of axis 1 and presented a high association with frugivorous species (A. planirostris, Carollia perspicillata, Sturnira lilium, and Platyrrhinus lineatus), including the sole records of Pyllostomus hastatus, Anoura geoffroyi, Artibeus lituratus, and S. lilium. The rocky outcrops were found distant from other habitats in the positive values of axis 1 and in the negative values of the axis 2. This habitat presented an association with a diverse array of species of guilds like the omnivore Phyllostomus discolor, the animalivorous Micronycteris bats, and insectivores Pteronotus gymnonotus, Peropteryx macrotis, and Eumops sp. Both riparian and shrubby caatingas were located closely in the ordination and presented a similar species composition. However, riparian caatingas demonstrated a closer association with insectivorous bats with some exclusive species like Molossops temminckii, Molossus molossus, and Rhogeessa hussoni. In contrast, shrubby caatingas showed a close association with nectarivores such as Lonchophyllas bats and X. vieriai, the latter, together with Neoplatymops mattogrossensis, were species exclusive to this habitat. The species A. planirostris and G. soricina were located relatively close to the centroid of the ordination, indicating a widespread occurrence in all habitats. Other species, like M. lavali and Tonatia bidens, seemed to favour riparian and shrubby caatingas, while vampire bats (D. rotundus and D. ecaudata) and L. mordax favoured rocky outcrops and shrubby caatingas.

Figure 4. Axes 1 and 2 of correspondence analysis of matrix of four caatinga habitats and 29 bats species in Rio Grande do Norte, Brazil from 2017 to 2019. Axis 1 explains 67.8% and axis 2 explains 21.9% of the observed variation.
Discussion
The spatial heterogeneity due to the natural occurrence of habitat mosaics in the Caatinga has been attributed as a factor that shapes the structure of bat assemblages in this dry forest (Carvalho-Neto et al. Reference Carvalho-Neto, da Silva, Santos, Rohde, Garcia and Montes2016, Willig Reference Willig1983). In the present study, we have found evidence that bat assemblages’ structure and species distribution changes according to the habitat type. As expected, we found differences in richness (Figure 2), abundance (Figure 3), species composition (Figure 4), and trophic guilds (Supplementary Materials 6) among the studied habitats. Such variation might be explained by the fact that each habitat provides specific environmental conditions and resources to be exploited by a distinct array of species with different life history traits (e.g., diet or roost generalists or specialists) and ecological adaptations (e.g., wing morphology, type of echolocation, and sensorial capacities) (Kunz Reference Kunz and Kunz1982, Kunz and Pierson Reference Kunz, Pierson and Nowak1994, Kalko Reference Kalko1998, Denzinger and Schnitzler Reference Denzinger and Schnitzler2013, Norberg and Rayner Reference Norberg and Rayner2016). In other regions of the Caatinga, differences in the structure of bat assemblages among habitats have also been reported (Gregorin et al. Reference Gregorin, Carmignotto and Percequillo2008, Novaes and Laurindo Reference Novaes and Laurindo2014, Rocha et al. Reference Rocha, Ruiz-Esparza and Ferrari2018, Sá-Neto and Marinho-Filho Reference Sá-Neto and Marinho-Filho2013, Willig Reference Willig1983). Thus, this indicates that this pattern is consistent for the northeastermost region of the Caatinga dry forest. As well, we found a high abundance of the frugivore A. planirostris in the region and likely explained by its ecological plasticity in its diet and roost selection.
Our results indicated a high variation in the richness per habitat (6–21 species). The habitats with the highest richness were riparian and shrubby caatingas and the rocky outcrops. The high richness associated with these habitats was not only due to the greatest sampling effort carried out in these areas, as indicated by the accumulation curves (Figure 2b). The high richness detected in these habitats might be explained by the presence of a high heterogeneity of resources (e.g., food, roost, and foraging areas). Consequently, more exploitable niches for a wider array of bat species than in less heterogeneous habitats (MacArthur and MacArthur Reference MacArthur and MacArthur1961, Connor and McCoy Reference Connor and McCoy1979).
The difference in the species composition of each habitat suggests that certain bat species and guilds preferred to forage more in certain specific habitat. Although some species were widespread (e.g., D. rotundus, G. soricina, A. planirostris, and M. lavali), their abundance ranking position varied between habitats (Figure 3) while other species were abundant in specific habitats (e.g., X. vieirai in shrubby and M. lavali in riparian). Additionally, the identity of rare species differed among habitats and some presented exclusive species. The reason of this pattern may be related to the differences in the vegetational structure and plant species composition of each foraging habitat. This was corroborated by the CA when we analysed the four best-surveyed habitats (Figure 4). The axis 1 of the CA (which explained 67.8% of the total variation) separated the humid forests enclaves. These enclaves have a well-developed vegetational stratification, from those semi-open habitats formed by mosaics of open habitat patches with different degrees of canopy closure, such as rocky outcrops, riparian, and shrubby caatingas.
Even though semi-open habitats presented relatively similar structures, the occurrence of particular of resources might be the reason of the structural differences in their bat assemblages. For instance, the shrubby caatinga besides having a high species richness, it presented the highest association of nectarivorous bats (Figure 3, 4 and Supplementary Material 6b). Shrubby caatingas are remarkably diverse in chiropterophilic plants, especially in columnar- and shrubby-cacti species (e.g., Pilosocereus spp. and Cereus jamacaru), and terrestrial bromeliads (Encholirium spectabile), among other species. These plants species provide abundant nectar and pollen resources in year-round basis to the diverse guild of nectarivores that forages in this habitat (Cordero-Schmidt et al. Reference Cordero-Schmidt, Barbier, Vargas-Mena, Oliveira, Santos, Medellín, Rodríguez-Herrera and Venticinque2017, Cordero-Schmidt et al. Reference Cordero-Schmidt, Maruyama, Vargas-Mena, Oliveira, Santos, Medellín, Rodríguez-Herrera and Venticinque2021). In other areas of Caatinga, nectarivorous bats are common but represented by one or two species (Rocha et al. Reference Rocha, Ruiz-Esparza and Ferrari2018, Sá-Neto and Marinho-Filho 2013, Willig Reference Willig1983, Silva Reference Silva2007, Soares et al. Reference Soares, Rocha, Bocchiglieri and Ferrari2019), while our studied shrubby caatinga was richer (4 spp.). It is worth highlighting the shrubby caatinga of Lajes, specifically in the Serra do Feiticeiro mountains. Here, shrubby caatingas are composed of extensive aggregations of columnar cacti (mostly Pilosocereus pachycladus) forming ‘cacti forests’ where all species of nectarivorous bats were recorded, including three endemics (L. mordax, L. inexpectata, and X. vieirai). Thus, nectarivorous bats seem to play a vital role in maintaining these unique cacti forests through pollination (Cordero-Schmidt et al. Reference Cordero-Schmidt, Maruyama, Vargas-Mena, Oliveira, Santos, Medellín, Rodríguez-Herrera and Venticinque2021).
Besides the abundant food resources for bats found in specific habitats, abundant roost resource is also a factor to consider in the structural processes of bat assemblages in this region of the Caatinga. Rocky outcrops, besides being very heterogeneous, with semi-open foraging areas with patches of shrubs, cacti, and Leguminosae trees, are abundant in underground cavities. For instance, in the limestone outcrops of Felipe Guerra, more than 350 caves have been recorded (Bento et al. Reference Bento, Cruz, Freitas and Campos2015). Consequently, in this habitat we found a high abundance of cave-roosting bats belonging to various trophic guilds like the frugivore A. planirostris, the sanguinivore D. rotundus and the nectarivores L. mordax, and G. soricina (Figure 3). Also, a high association of other species, such as the animalivore Micronycteris sanborni, the omnivore Phyllostomus discolor, and the insectivores Pteronotus gymnonotus and Peropteryx macrotis, all cave-roosting bats recorded in RN (Vargas-Mena et al. Reference Vargas-Mena, Cordero-Schmidt, Bento, Rodríguez-Herrera, Medellín and Venticinque2018a, Reference Vargas-Mena, Alves-Pereira, Barros, Barbier, Cordero-Schmidt, Lima, Rodríguez-Herrera and Venticinque2018b). Thus, this habitat provides an important roosting resource for local populations of several species.
Humid forest enclaves presented the most distinct assemblage structure of all habitats. Frugivorous bats were rich (A. planirostris, A. lituratus, S. lilium, C. perspicillata, and P. lineatus) and abundant (91 % of captures, Supplementary Material 6b) and harboured several exclusive species (Pyllostomus hastatus, Anoura geoffroyi, Artibeus lituratus, and S. lilium). This frugivore dominance is consistent with other humid forest enclaves across the Caatinga (Sousa et al. Reference Sousa, Langguth, Gimenez, Pôrto, Tabarelli and Machado2004, Silva Reference Silva2007, Novaes et al. Reference Novaes, Laurindo, Oliveira, Barreto and Avilla2013, Rocha et al. Reference Rocha, Ruiz-Esparza and Ferrari2018). In these enclaves, 80% of plants are zoochoric species, whereas, in adjacent areas of Caatinga, only 44% are zoochoric, while most are anemochoric (Griz and Machado Reference Griz and Machado2001, Vicente et al. Reference Vicente, Santos, Tabarelli, Leal, Tabarelli and Silva2003, Machado et al. Reference Machado, Prata and Mello2012). Thus, the abundant fruit resources available are likely key for maintaining frugivore populations in this unique habitat.
The riparian caatinga might represent the best example of how Caatinga’s natural heterogeneity can favour a high relative richness and abundance of bats in the assemblage. This habitat harbors a diverse plant community with a mixture of plant species of adjacent habitats and riparian species, plus a critical resource in this semiarid dry forest, such as water bodies. In our study, besides this habitat harbouring the highest richness (17 spp.), we recorded the highest richness (7 spp.) and abundance of insectivorous bats (e.g., Myotis lavali). Therefore, the richness of insectivores contributes relatively more to the richness of this habitat (Table 2). This pattern is similar to that reported by Novaes et al. (Reference Novaes, Laurindo and Souza2015) in areas near water bodies in northwestern caatinga. In arid and semiarid ecosystems, insectivorous bats are abundant near water bodies since they forage for aquatic emergent insects and to drink water (Grindal et al. Reference Grindal, Morissette and Brigham1999, Korine and Pinshow Reference Korine and Pinshow2004, Costa et al. Reference Costa, Luz and Esbérard2012, Razgour et al. 2010). Riparian caatingas might function as an ‘oasis’ for insectivores and other guilds, especially during the harsh dry season, but this potential role still needs further investigation.
Table 2. Species composition and number of captured of individuals (without effort correction) per caatinga habitat surveyed during 2017–2019 in the Caatinga of Rio Grande do Norte State. Orch/gard means orchards and gardens
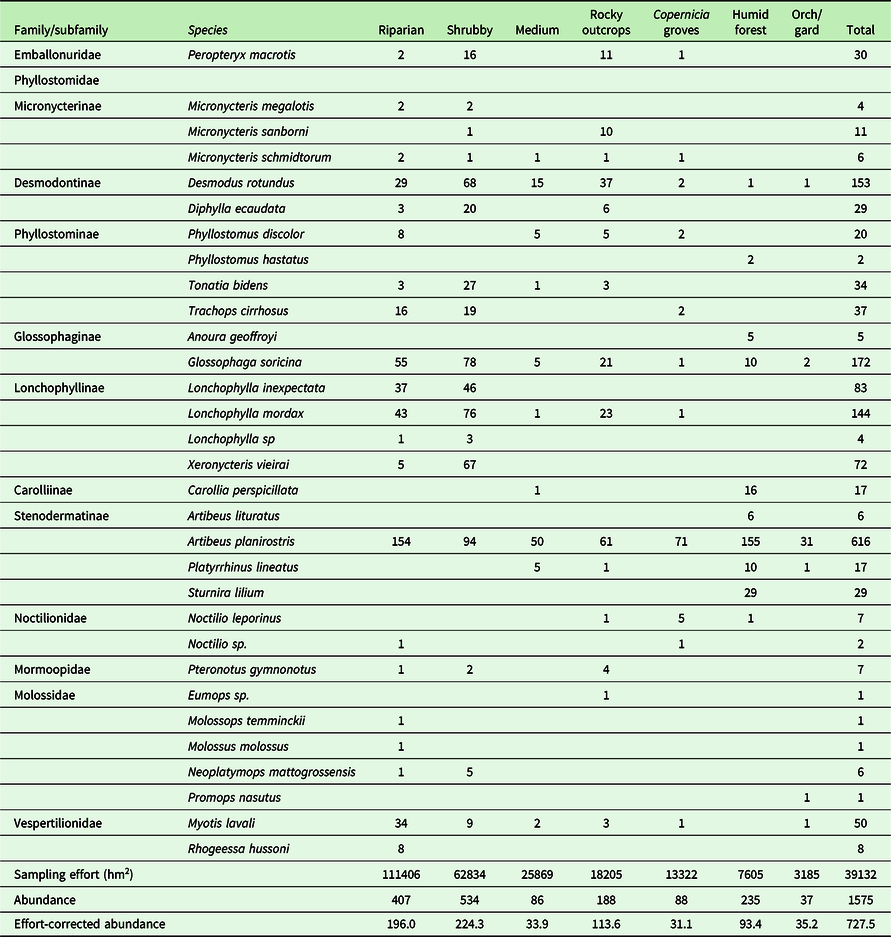
Bats belonging to the family Phyllostomidae represented the highest abundance and dominance in all habitats, as expected, since they represent more than 50% of all recorded bats in the Caatinga (Silva et al. Reference Silva, Delgado-Jaramillo, Souza Aguiar and Bernard2018). Notably, A. planirostris presented considerable dominance in all habitats and a superabundance at the riparian caatinga and the humid forest enclaves (Figure 3), including in relatively poor-sampled habitats like in Copernicia groves and gardens and orchards. In general, A. planirostris is a widespread species that occurs from forested areas habitats to urban areas (Hollis et al. Reference Hollis, Spix and Bat2005). This bat in the Caatinga can roost in underground cavities (Vargas-Mena et al. Reference Vargas-Mena, Cordero-Schmidt, Bento, Rodríguez-Herrera, Medellín and Venticinque2018a) and tree foliage (Vargas-Mena com. pers.) and is reported to feed on a wide variety of food items such as fruits of native and exotic plants, insects, nectar, and even leaves (Carvalho et al. Reference Carvalho, Maas, Peracchi and Gomes2016, Cordero-Schmidt et al. Reference Cordero-Schmidt, Medeiros-Guimarães, Vargas-Mena, Carvalho, Ferreira, Rodríguez-Herrera and Venticinque2016, Hollis et al. Reference Hollis, Spix and Bat2005, Rocha et al. Reference Rocha, Domingos-Melo, Zappi and Machado2019). This roost and diet flexibility likely provides advantages to occupy more habitats than other frugivores in Caatinga’s harsh environment.
In other areas of Caatinga, A. planirostris has been reported as the most abundant species (Feijó and Rocha Reference Feijó and Rocha2017, Gregorin et al. Reference Gregorin, Carmignotto and Percequillo2008, Silva Reference Silva2007), but also reported as absent in the top most abundant species (Novaes and Laurindo Reference Novaes and Laurindo2014, Rocha et a l. Reference Rocha, Ruiz-Esparza and Ferrari2018, Soares et al. Reference Soares, Rocha, Bocchiglieri and Ferrari2019). Its superabundance in some areas and rarity in others might be related to the seasonality of food and capture probability, but these contrasting patterns along its geographic distribution need further investigation. On the other hand, Carollia perspicillata was extremely rare in the studied habitats, which is surprising since it is one of the most abundant species in other Caatinga regions (Beltrão et al. Reference Beltrão, Zeppelini, Fracasso and Lopez2015, Feijó and Rocha Reference Feijó and Rocha2017, Gregorin et al. Reference Gregorin, Carmignotto and Percequillo2008, Novaes and Laurindo Reference Novaes and Laurindo2014, Silva et al. Reference Silva, Dias, Martins, Guedes, de Almeida, da Cruz, Serra-Freire, Damascena and Peracchi2015, Soares et al. Reference Soares, Rocha, Bocchiglieri and Ferrari2019, Willig Reference Willig1983). In our studied sites, plants and fruits on which the species prefers to eat were never observed (e.g., Piperaceae) (Flemming Reference Flemming1988), likely explaining its rarity in this region of the Caatinga. This species seems to be more common in more humid areas (e.g., humid forest enclaves).
We found diverse bat assemblages in protected areas, highlighting their value in the conservation of bat populations and their habitats in RN. However, more sampling effort is necessary in all protected areas (Figure 2a), and these sites should be a priority in future bat inventories. Habitats located in non-protected areas, like the shrubby Caatinga in Lajes and the humid forest enclaves in Martins, should be considered for conservation actions since, in their assemblage, the dominant species are likely critical for the maintenance and regeneration process of these unique habitats through seed dispersal and pollination. Also, undersampled habitats like Copernicia groves should be explored more, including the potential role of A. planirostris in the seed dispersion of Copernicia prunifera, an endemic and important socio-economical palm in the Caatinga (Rocha et al. Reference Rocha, Silva, Dantas and Vieira2015). Finally, we encourage using acoustics recordings and roost search as complementary methods to mist-netting studies in the task to understand better the bat fauna of the heterogenous Caatinga dry forest.
Supplementary material
To view supplementary material for this article, please visit https://doi.org/10.1017/S0266467423000317
Acknowledgements
We would like to thank J.B. Lima, I. Mendes-Freitas, J.P.M. Araújo, V. Santana, and M.J. Bezerra for their patience and invaluable help during the field data collection and the Wildlife Conservation Society Brazil for key support in field logistics.
Financial support
This study was supported by the Conselho Nacional de Desenvolvimento Científico e Tecnológico (J.C.V.M, E.C.S, B.R.H., Special Visiting Researcher-PVE grant number: 401467/2014-7; and E.M.V, productivity grant number #308040/2017-1), the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (J.C.V.M., E.C.S. post-graduation scholarships), and The Rufford Foundation (Grant: 25057-1).
Competing interests
The authors declare none.








