


The limited available data that trace succeeding generations of humans show that gestations of male twins produce, on average, relatively few grandchildren for their parents (Gabler & Voland, Reference Gabler and Voland1994; Lummaa, Reference Lummaa2001). The fitness deficit of these gestations arises principally, although not solely (Lummaa et al., Reference Lummaa, Pettay and Russell2007), from relatively high rates of infant and child mortality among male twins born into threatening environments.
Evolutionary theory posits that natural selection would conserve mechanisms that precluded a mother from investing her limited energy in gestating a fetus unlikely to survive to reproductive age if born (Forbes, Reference Forbes1997; Møller, Reference Møller1997; Stearns, Reference Stearns and Stearns1987; Wells, Reference Wells2000). Much literature speculates that fetuses signal their fitness to a non-conscious decisional biology that also senses environmental threats to maternal and infant wellbeing (Catalano et al., Reference Catalano, Margerison-Zilko, Goldman-Mellor, Pearl, Anderson, Saxton and Kharrazi2012; Erlebacher, Reference Erlebacher2010; Sales et al., Reference Sales, Grant, Catalano and Jabbour2011; Vigano et al., Reference Vigano, Mangioni, Pompei and Chiodo2003). This biology somehow combines information to estimate the yield of grandchildren per unit of maternal investment required to sustain the prospective infant in prevailing environments. If the yield falls below some criterion that varies from woman to woman but exhibits a central tendency in the population, the gestation has increased risk of spontaneously ending. Perhaps reflecting their relatively low fitness, male twin gestations exhibit higher incidence of fetal mortality than gestations of female twins and of singletons of either sex (Dailey et al., Reference Dailey, Jayakrishnan, Phipps, Raker and Chien2009; Melamed et al., Reference Melamed, Yogev and Glezerman2009; Mulla et al., Reference Mulla, Plavsic, Ortiz, Nuwayhid and Ananth2013; Rydhstroem & Heraib, Reference Rydhstroem and Heraib2001; Tan et al., Reference Tan, Wen, Walker, Fung, Demissie and Rhoads2004).
Kahneman (Reference Kahneman2011) has speculated that leaving some decisions outside of awareness may have conferred fitness benefits and, therefore, been conserved by natural selection. Mechanisms that spontaneously abort least fit gestations, for example, may conflict with countervailing traits that compel mothers, via cognition reinforced by culture, to invest in children regardless of fitness (Clutton-Brock, Reference Clutton-Brock1991). Mutations that separated pre- and postnatal maternal investment decisions, and that reduced the influence of awareness in the former, would offer a fitness advantage.
We suggest that, consistent with Kahneman's (Reference Kahneman2011) speculation, at least one decision fundamental to human fitness, whether to continue the gestation of a male twin, regularly proceeds without awareness and yet reveals preferences including the willingness to assume risk. We support this suggestion by showing that household surveys that predict financial risk taking in the population also predict biological risk taking as seen in gestational outcomes.
Little literature has pondered whether risk aversion in the non-conscious decisional biology that ends or extends human gestation varies over time with risk aversion in conscious assessments that inform, for example, economic choices. One study reports that decreases in the self-reported willingness of households to assume financial risk predict increases in the incidence of low weight male births in Sweden (Catalano et al., Reference Catalano, Karasek, Gemmill, Falconi, Goodman, Magganas and Hartig2014). The authors argue that the association arises because increasing risk aversion in decisional biology leads to spontaneous abortion of higher risk/lower reproductive return fetuses, but advanced medical technology ‘converts’ some fraction into live preterm, and therefore low weight, births. This argument leads to the suspicion that other, less clinically mediated indicators of selection against higher risk fetuses should also increase when the population reports declining willingness to assume financial risk. We test that suspicion using the twin ratio among male births.
We tested the hypothesis that the likelihood of a twin among males born in Sweden would fall below expected values when the intention of Swedes to assume economic risks similarly falls below expected values. We used Sweden for our test because it has among the very best vital statistics systems and twin registries in the world. Prior research validating selection in utero has, for this reason, also used Swedish data (Catalano & Bruckner, Reference Catalano and Bruckner2005, Reference Catalano and Bruckner2006; Catalano et al., Reference Catalano, Bruckner and Smith2008). Sweden, moreover, has a longstanding and highly respected survey of consumer intentions, from which we derived our independent variable.
Materials and Methods
Data
We used the Micro Index of the Swedish Consumer Tendency Survey (MISCT) to gauge shifts in risk aversion (National Institute of Economic Research, 2012). The nationally representative telephone survey of Swedish consumers aged 16–84 years yields 1,500 interviews each month. This survey asks respondents a series of questions regarding the state of the economy as well as their personal and family finances, their expectations of the future course of these circumstances, and their plans to purchase consumer durables. The survey asks, for example, how likely respondents are to purchase durable goods in general, as well as make specific major purchases such as an automobile or dwelling.
The MISCT combines responses to questions on the present and anticipated (12 months in the future) state of personal finances and on intentions to purchase durable goods over the next 12 months. The focus on durable goods makes the survey particularly salient for our purposes because their consumption reportedly varies more with risk aversion than does the consumption of non-durables (Carroll et al., Reference Carroll, Fuhrer and Wilcox1994; Howrey, Reference Howrey2001; Huth et al., Reference Huth, Eppright and Taube1994). Responses to questions are then categorized as indicating expectations of greater or lesser risk in purchasing durable goods. The index equals the percent scored greater risk subtracted from the percent scored lesser risk.
We obtained monthly counts of twin and singleton births from the Swedish Medical Birth Registry (Swedish National Board of Health & Welfare, 2012). We requested monthly counts to allow sufficient temporal definition to test our theory but not so defined as to risk violations of anonymity that could arise from using data from shorter time periods. We began our analyses with the first month for which data were available for both the birth and consumer confidence measures (January 1993). Our analyses end with December 2010, the most recent month for which we could obtain data at the time of our request.
Design
Testing our hypothesis required us to specify, a priori, the expected temporal relationship between lower than expected consumer confidence and lower than expected twin male births. Our specification reflected current knowledge of the timing of unintentional fetal loss.
Fetal loss early in gestation remains difficult to observe, but research suggests that the highest rates appear in the first trimester, then decline until approximately the 16th week of gestation, when the risk plateaus for approximately 6 weeks, then rises again before birth (Ammon Avalos et al., Reference Ammon Avalos, Galindo and Li2012; Goldhaber & Fireman, Reference Goldhaber and Fireman1991).
Abortuses in the first trimester exhibit high rates of chromosomal abnormalities that make survival as infants, let alone reproduction as adults, highly unlikely regardless of environmental circumstances or maternal investment (Kline & Stein, Reference Kline, Stein, Edmonds and Bennet1987). These spontaneous abortions would, therefore, likely occur at virtually any level of risk aversion.
The distribution of spontaneous abortions among chromosomally normal fetuses also peaks, although less dramatically than among abnormal fetuses, around the 12th week and also declines to a plateau that extends from the 16th through the 24th week when it drops precipitously (Kline & Stein, Reference Kline, Stein, Edmonds and Bennet1987). Males predominate among the 16th to 24th week normal abortuses (Byrne et al., Reference Byrne, Warburton, Opitz and Reynolds1987; Hassold et al., Reference Hassold, Quillen and Yamane1983). Based on these data, we speculated that although non-conscious decisional biology works throughout pregnancy, we would detect its effects primarily in the 16th through 24th weeks. Earlier effects among the frequent losses due to chromosomal abnormalities and later effects among relatively rare losses may both prove difficult to detect.
Twins gestate for a shorter period than singletons (Alexander et al., Reference Alexander, Kogan, Martin and Papiernik1998; Kiely, Reference Kiely1998). Data suggest that Swedish twin births increase at the 34th week of gestation with most occurring by the 38th week (Cheung et al., Reference Cheung, Yip and Karlberg2000). The effect of rising risk aversion in gestational biology should, therefore, first appear ten weeks, or about 2.5 months, after a decline in the MISCT. This early effect would appear in the cohort exposed at the 24th week of gestation (i.e., latest week in the critical period), and born at the 34th week (i.e., earliest week of probable birth period). The last effect would appear 22 weeks, or roughly 5.5 months, after the shock in the cohort exposed at the 16th week of gestation (i.e., earliest week in the critical period) and born the 38th week (i.e., latest week of the probable birth period).
Monthly changes in consumer tendencies logically occur in the time between an earlier and later survey, implying that an unexpected shift to greater risk aversion occurred in the month before its report. Its effect, if any, on the decisional biology of gestation would, therefore, manifest as unexpectedly few male twin births approximately two to six months after the consumer confidence index unexpectedly declines.
Analyses
We tested our hypothesis in four steps. First, we estimated the expected monthly values of the MISCT so that we could identify months in which the Swedish population reported unexpectedly high risk aversion (i.e., unexpectedly low intention to purchase durable goods). Statistical tests of association typically assume that the expected value of any variable equals the mean of all its observations. Variables measured over time, however, often violate this assumption because they exhibit autocorrelation in the form of secular trends, cycles, or the tendency to remain elevated or depressed, or to oscillate, after high or low values. The expected value of an observation in such a series is not the mean of all observations but rather the value predicted by autocorrelation.
We used the method devised by Box and Jenkins to detect and model autocorrelation in the monthly MISCT (Box et al., Reference Box, Jenkins and Reinsel2008). The method –– Autoregressive, Integrated, Moving Average (ARIMA) modeling –– identifies which of a very large family of mathematical expressions best predicts measurements made over time. The method assumes that the measurements are passed through an unobserved filter, or model, that imposes autocorrelation upon them. The approach looks for patterns in the data that narrow the search to likely filters and then applies estimates of ‘fit’ to identify the best fitting model. These models yield expected values that approximate what would have occurred if the circumstances that produced earlier values remained unchanged.
The residuals (i.e., difference between the expected and observed values) of the ARIMA model for the MISCT estimate the degree to which monthly risk aversion among Swedes either increased or decreased from expected levels. Consistent with our focus on risk aversion, we scored as one those months with unexpectedly low consumer confidence (i.e., those with negatively signed residuals from the ARIMA models estimated in step 1), and we scored all other months zero. The resulting variable identifies months in which the Swedish population reports risk aversion greater than expected from history (i.e., from autocorrelation).
Second, we calculated the monthly odds of a twin among male live births (excluding multiple births greater than twins). Third, we applied Box-Jenkins methods to the odds to detect and model autocorrelation. In other words, we arrived at the expected odds of a male twin in any month from ‘history’ (i.e., autocorrelation).
Fourth, we tested our hypothesis by estimating the equation formed by adding, as a predictor, the binary variable derived in step 1 to the model developed in step 3. We specified the equation such that unexpectedly high risk aversion predicted the odds of a twin among male births in the same month as risk aversion, as well as up to eight months later. Our theory predicts that at least one coefficient for cohorts born two through six months after periods of unexpectedly great risk aversion will be significantly less than zero. We set significance conservatively at p < .025 (single-tailed test) because we inferred support for the hypothesis if any of the five coefficients were significantly less than zero.
Results
The monthly odds of a twin among live male births over our test period ranged from 0.019 to 0.043 with a mean of 0.029. The monthly MISCT ranged from -21.3 to 18.8 with a mean of 5.4.
Figure 1 shows the observed and expected values of the MISCT. The best fitting ARIMA model of the MISCT included taking first differences (i.e., value at month t subtracted from month t-1) to adjust for several periods of secular trend during the test period (e.g., April 1995 to April 2000). The model also included a moving average parameter indicating short term ‘memory’, in which unusually high or low differences preceded high or low differences of declining absolute size by one month.
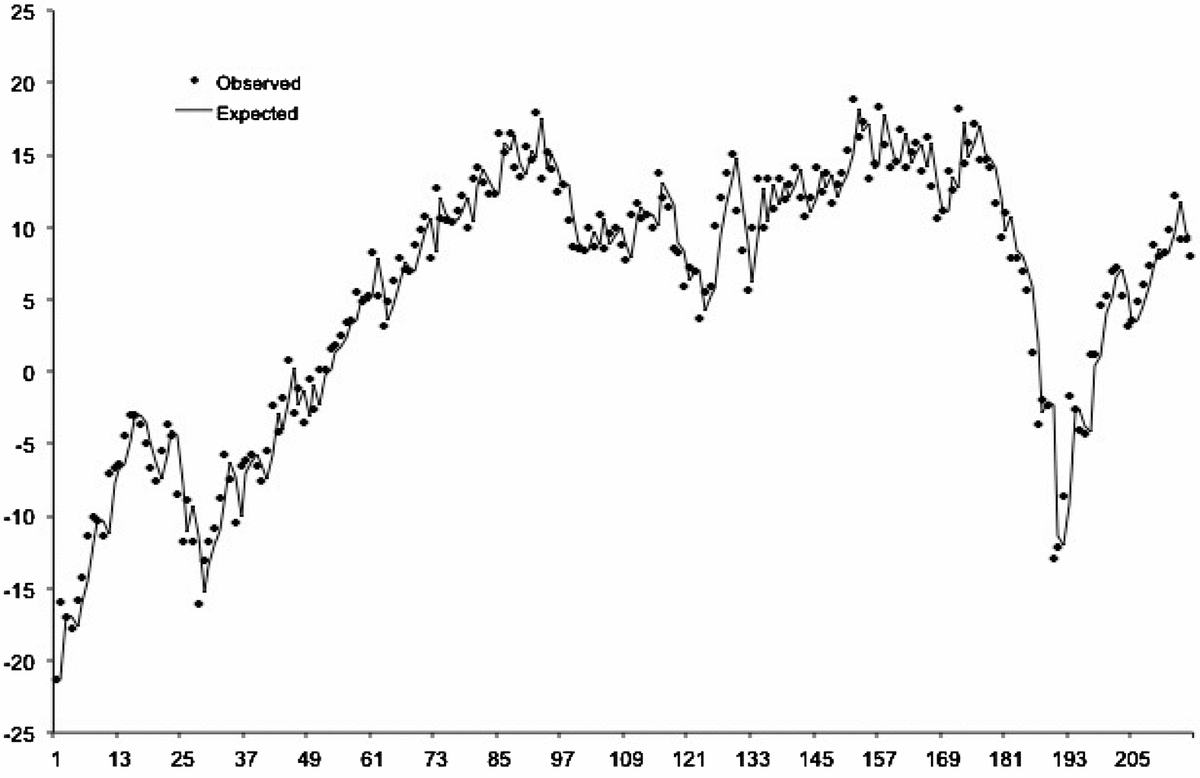
FIGURE 1 Expected (from autocorrelation) and observed values of the Micro Index of the Swedish Consumer Tendency Survey for 216 months beginning January 1993 and ending December 2010.
Figure 2 shows the observed and expected values of the monthly odds of a twin among male births. The best fitting ARIMA model of this series included autoregressive parameters at 12 and 23 months, indicating seasonality in the odds of a twin among male births.
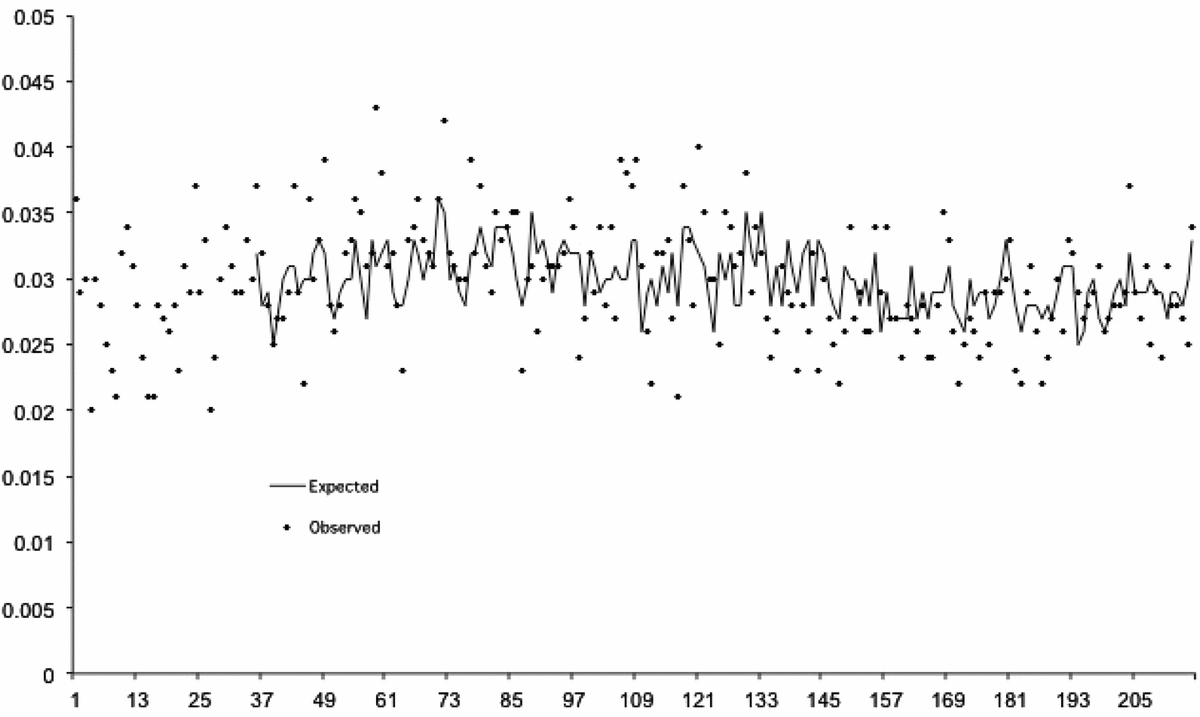
FIGURE 2 Expected (from autocorrelation) and observed monthly odds of a twin among males born in Sweden for 216 months beginning January 1993 and ending December 2010 (expected values begin later because 25 months lost to modeling).
Table 1 shows the results of estimating the test equation described in the fourth step above. Consistent with our hypothesis, the odds of a twin among male births fell below expected values two months after Swedes reported unexpectedly low consumer confidence.
TABLE 1 Coefficients and Standard Errors for Model Predicting Odds of a Twin Among Male Births in Sweden for 216 Months Beginning January 1993 and Ending December 2010
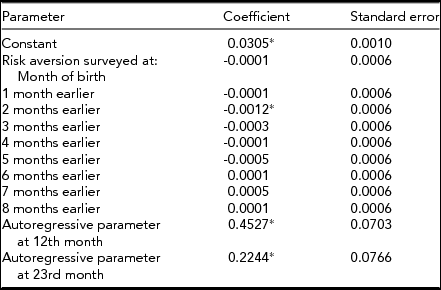
*p < .025; 1-tailed test.
We conducted several robustness tests of our analyses. First, we used the methods of Chang et al. (Reference Chang, Tiao and Chen1988) to detect and adjust for outliers in the dependent variable that may have inflated our confidence intervals and led to false acceptance of the null for birth cohorts other than those 2 months after unexpectedly high risk aversion. We detected five outliers, but adjusting for them did not change our findings.
Second, we pared the parameters with non-significant coefficients from the model and estimated it again. The coefficient for the cohort born two months after unexpectedly high risk aversion remained significantly less than zero.
Third, we replaced our binary risk aversion variable in the pared model with a variable in which we left the negatively signed residuals of the MISCT in their original continuous values rather than rescoring them to 1s. Estimating this model tested a linear dose response to risk aversion. The results of the test did not change.
As a fourth check on the robustness of our findings, we tested our hypothesis using the ‘common filtering’ approach. We ‘filtered’ the male twin ratio through the best fitting model of the stationary MISCT scores. An association between the residuals of applying the same model to both series cannot be attributed to autocorrelation (because the residuals of the model derived from and applied to the MISCT series will have no autocorrelation) and will generalize back to the original series (because both variables were identically transformed). Applying common filtering to our two variables revealed that the residuals of the male twin ratio varied positively with the residuals of the MCCI observed two months earlier. This is the same result we derived from our method.
To express our results in the familiar ‘effect on odds’ metric, we transformed our dependent variable to natural logarithms and estimated our original test equation again. The coefficient for births two months after unexpectedly great aversion to risk (i.e., -0.0359) suggested that the odds of a twin among male births fell by approximately 3.5% in the affected cohorts. We arrived at this estimate by subtracting the antilog of the coefficient from 1, and multiplying the difference by 100.
Discussion
We found that the odds of a twin in monthly male birth cohorts declined two months following unexpected increases in self-reported risk aversion in the Swedish population. This implies that males in twin gestations were more vulnerable to loss when the population became more risk averse.
Scholarship in psychology, economics and neuroscience assumes that humans have the capacity to make decisions after estimating and comparing the risks and rewards of available options. Nearly all this work focuses on decisions humans report making (Kahneman, Reference Kahneman2011), but some have studied choices made outside awareness (Dijksterhuis & Aarts, Reference Dijksterhuis and Aarts2010). Wegner and Smart (Reference Wegner and Smart1997), for example, describe a class of decisions that the agent cannot report making even though the choices reveal his or her preferences. Our findings imply such a connection between population risk sentiment and the non-consciously controlled biologic circumstance of fetal loss.
Our test benefits from a large study population observed over many months. We included all male singleton and twin births in 216 Swedish monthly cohorts born from January 1993 through December 2010. This long series of monthly data allowed us to use the most rigorous methods available to control for autocorrelation in both our independent and dependent variables. Our finding cannot, therefore, arise spuriously from shared trends, seasonality, or regression to expected levels after high or low values.
Our analyses have limitations. The data available to us did not permit separating male twins into those from male–male and male–female gestations. Although available data suggest that any gestation producing a male twin ultimately yields fewer grandchildren than do other gestations, male–male gestations appear to take a heavier toll on a mother's health than do mixed sex twin gestations (Lummaa, Reference Lummaa2001). Indeed, male fetal sex is associated with preterm delivery, and male–male twin sets display additional higher risk than male–female pairs (Dailey et al., Reference Dailey, Jayakrishnan, Phipps, Raker and Chien2009; Melamed et al., Reference Melamed, Yogev and Glezerman2009; Rydhstroem & Heraib, Reference Rydhstroem and Heraib2001; Tan et al., Reference Tan, Wen, Walker, Fung, Demissie and Rhoads2004). Males in a male–male pair are likely lower birth weight than those with a female twin (Melamed et al., Reference Melamed, Yogev and Glezerman2009). This circumstance intimates that self-reported risk aversion may predict selection in utero against the male twin sets more than mixed-sex pairs. Females from mixed-sex twin gestations exhibit lower fitness than those from same sex sets (Lummaa et al., Reference Lummaa, Pettay and Russell2007) implying that selection against males in mixed sex gestations may also be adaptive in threatening environments.
Our theory suggests that the odds of a twin among male births should gauge selection in utero over time. The non-conscious decisional biology of gestation could, however, also target less fit singletons for spontaneous abortion. Our estimate of the effect of unexpected risk aversion on the odds of a twin among male births could, therefore, be biased downward in that the risk aversion also removes less fit singletons from the denominator of the odds.
Further theory development should explore the implications of risk aversion for female twins. The argument we offer for using gestations of male twins as targets for selection in utero does not apply to female twins. Unlike gestations involving male twins, those producing female twins ultimately yield as many, and in stressful circumstances may yield more, grandchildren than do female singletons (Lummaa, Reference Lummaa2001). This circumstance implies that gestations of female singletons may spontaneously abort before those of female twins with increasing risk aversion in the population (Catalano et al., Reference Catalano, Saxton, Bruckner, Goldman and Anderson2009).
Our findings build on a connection noted between selection in utero and ‘portfolio theory’ (Catalano et al., Reference Catalano, Karasek, Gemmill, Falconi, Goodman, Magganas and Hartig2014; Forbes, Reference Forbes2009; Kahneman & Tversky, Reference Kahneman and Tversky1979: Markowitz, Reference Markowitz1952). This theory suggests that a mother has a portfolio of living and/or potential children among whom she must invest her limited time and energy. At critical points in each pregnancy, her non-conscious decisional biology makes choices about whether to continue investing in the gestation given the current environment and as a reflection of other opportunities in her portfolio. Because that non-conscious determination has a profound effect on the fitness of the mother and of her offspring, natural selection would conserve mutations that served an adaptive advantage (Dawkins & Carlisle, Reference Dawkins and Carlisle1976).
Conclusions
While our findings may not intuitively appear as risk-factor epidemiology intended to inform clinical intervention, they contribute to our understanding of temporal variation in the characteristics, including fitness, of human birth cohorts. We use an evolutionary approach to improve our understanding of a complex and poorly understood public health phenomenon –– fetal loss of male twins. In doing so, we have provided important insights about risk aversion and human gestations that are rarely discussed in clinical settings.
Acknowledgments
The Robert Wood Johnson Health and Society Scholars Program funded the research described in this article. Additional funding was provided through NICHD grant #HD07275.



