


Implications
Feeding excessive dietary protein to sows increases nitrogen excretion into the environment. Hence, it is important that the protein provision matches the actual amino acid requirement of the sow. This study showed that increased protein intake of hyper-prolific lactating sows prevents excessive BW loss and decreases body protein mobilization, as the extra dietary protein was utilized for milk production. Minimizing the body mobilization can ensure better performance of sows in the following reproductive cycle. The higher milk yield and nutrient concentrations resulted in higher litter gains, which is of great economic importance to the pig producer.
Introduction
Sows have become larger and leaner during the last decades, and their milk yield has increased (Hansen et al., Reference Hansen, Strathe, Kebreab, France and Theil2012) to cover the requirement of the large litters. These tremendous changes probably have increased the dietary requirement of protein and amino acids of the sow. For many years focus has been on determining the requirement of single amino acids, especially lysine, and not the requirement of balanced CP. The supply of protein above the requirement will cause deamination of the surplus amino acids and increased excretion of N into the environment (Hansen et al., Reference Hansen, Strathe, Theil and Kebreab2014; Huber et al., Reference Huber, de Lange, Krogh, Chamberlin and Trottier2015). In another part (Strathe et al., Reference Strathe, Bruun, Geertsen, Zerrahn and Hansen2017a) of the current study, the dietary standardized ileal digestible (SID) CP concentration for maximizing litter gain was determined to be 135 g/kg. Litter gain is regarded as an important determinant of nutrient requirements of lactating sows (Trottier et al., Reference Trottier, Johnston, de Lange and Farmer2015). However, it is also important to understand how the dietary provision of CP influences the mobilization of body tissues and milk production when determining the requirement of lactating sows. Sows often turn catabolic in order to maintain their milk production (Strathe et al., Reference Strathe, Bruun and Hansen2017b). Under-supplying dietary protein can cause a high mobilization of body protein (King et al., Reference King, Toner, Dove, Atwood and Brown1993; Mejia-Guadarrama et al., Reference Mejia-Guadarrama, Pasquier, Dourmad, Prunier and Quesnel2002), since milk production has a very high priority. Depleted body protein pools must be restored during the following gestation, which is energetically unfavourable (Dourmad et al., Reference Dourmad, Etienne and Noblet1996; Bender, Reference Bender2012) and should therefore be avoided. The extent of the catabolic state will be reflected in changes in body composition, plasma metabolite concentrations and milk production (Dourmad et al., Reference Dourmad, Noblet and Etienne1998). Measurements of plasma urea N (PUN) concentrations, milk composition and body composition can be used as alternative markers for the optimum SID CP (Coma et al., Reference Coma, Zimmerman and Carrion1996; Yang et al., Reference Yang, Pettigrew, Johnston, Shurson, Wheaton, White, Koketsu, Sower and Rathmacher2000), because it is not given that the metabolism of the sow is optimized at the same dietary SID CP concentration as the litter gain.
Hence, the objective of this experiment was to estimate the dietary SID CP requirement of lactating sows using litter gain as the main determinant supported by measurements on several traits reflecting sow metabolism. It was hypothesized that increased dietary protein provision would decrease body protein mobilization and improve milk production through increased yield and nutrient concentrations as well as affect sow plasma concentrations of metabolites related to protein and fat metabolism.
Material and methods
This experiment was carried out on a subsample of 92 sows from a larger trial with 544 sows (Strathe et al., Reference Strathe, Bruun, Geertsen, Zerrahn and Hansen2017a).
Treatments, animals and housing
The study was conducted in a commercial piggery using a total of 92 parity 2 and 3 sows (Danish Landrace × Danish Yorkshire, DanAvl) inseminated with Duroc semen (Hatting KS, Horsens, Denmark). The animals were studied from 4 days prepartum (−4.1 ± 1.6) to 24 days postpartum (24.4 ± 1.2) when piglets were weaned. Sows were randomly allocated to one out of six dietary treatments with increasing dietary provision of SID protein (Table 1) and stratified for parity in a complete block design from day 2 postpartum. Diets were formulated to be isoenergetic on net energy basis (EvaPig®, 2008). Sows were fed using a SpotMix feeding system (Schauer Agrotronic, Prambachkirchen, Austria), and meals for each sow were weighed and registered by the feeding system. From day 4 prepartum to day 2 postpartum all sows were fed the same commercial formulated lactation diet based on wheat, soybean meal and sugar beet pulp complying with Danish recommendations (Tybirk et al., Reference Tybirk, Sloth and Jørgensen2014). From day 2 to day 10 of lactation, sows were fed twice daily, and from day 10 onwards, sows were fed three times per day. The sows were fed 2.3 kg from day 2 postpartum, and feed allowance was gradually increased to 8.4 kg at day 17. Sows had drinking nipples in the trough and free access to water. Feed residuals were not recorded, but feed allowance was adjusted for individual sows daily, and the feed allowances mentioned above were used as maximum for the sows, so some sows would consume less.
Table 1 Dietary composition (as-fed) of diets fed to sows from day 2 postpartum to weaning

Lys=lysine; Met=methionine; Thr=threonine; Trp=tryptophan; Leu=leucine; Ile=isoleucine; His=histidine; Phe=phenylalanine; Val=valine; SID=standardized ileal digestible.
1 Leci E Basic, Evilec ApS, Sandbjergvej 26, 6000 Kolding, Denmark.
2 Provided per kilogram of the diet: 8900 IU vitamin A; 890 IU vitamin D3; 167 mg dl-alfa-tocopherol, 4.24 mg vitamin K3; 2.23 mg vitamin B1; 5.57 mg vitamin B2; 3.34 mg vitamin B6; 0.02 mg vitamin B12; 16.70 mg d-pantothenic acid; 22.26 mg niacin; 1.67 mg folic acid; 89.04 mg iron (FeSO4); 13.00 mg copper (CuSO4); 44.52 mg manganese (MnO); 0.22 mg iodine (Ca(IO3)2); 0.37 mg selenium (Na2SeO3).
3 Phyzyme XP provided 500 phytase activity (FTU) per kilogram of diet (Danisco Animal Nutrition DuPont, Marlborough, UK).
4 SID values used in diet formulation were from Pedersen and Boisen (Reference Pedersen and Boisen2002).
5 Feed samples were sampled regularly during the experimental period, and 12 feed samples were analysed per dietary treatment.
6 Calculated from the analysed composition and digestibilities used when formulating the diets.
Each week a batch of six sows was selected for the experiment, moved to the farrowing unit day 4 prepartum and placed in individual farrowing crates. Twenty-four hours postpartum (day 2) the litters of the experimental sows were standardized to 14 piglets (average piglet weight 1.76 ± 0.24 kg). Apart from the observations made in the experiment all animals were managed according to the general routines of the herd. Health was monitored by the stock personnel, and normal herd practices for management, treatments and vaccinations were followed. When dead or very weak piglets were removed, the date and weight of the excluded piglets were recorded.
Data collection, measurements and chemical analyses
Information on the collection of feed samples and analyses is given in Strathe et al. (Reference Strathe, Bruun, Geertsen, Zerrahn and Hansen2017a). On 544 sows, BW and back fat (BF) thickness were measured at day 2 postpartum and at weaning, and litter weight was recorded at litter standardization and at weaning (Strathe et al., Reference Strathe, Bruun, Geertsen, Zerrahn and Hansen2017a).
On 70 sows (10 to 12 per dietary treatment), blood, urine and milk samples were collected. Blood samples were collected in 10 ml EDTA tubes (BD Vacutainer, Plymouth, UK) from the jugular vein at days −4, 3, 10, 17, and 24 postpartum 4 h after the morning feeding. Samples were placed on ice and centrifuged for 10 min (1560×g) at room temperature, and plasma was then harvested and stored at −20°C in 1.5-ml microcentrifuge tubes (Kruuse, Langeskov, Denmark) until analysis. Plasma was analysed for concentrations of PUN, creatinine, glucose, lactate and non-esterified fatty acids (NEFA), triglycerides, cholesterol, total protein, albumin, alanine amino transferase (ALT), aspartate amino transferase (AST), gamma-glutamyl transferase (GGT) and alkaline phosphatase (Advia 1800 Chemistry System; Siemens, Denmark).
Milk samples were collected on days 3, 10 and 17 postpartum. Piglets were removed from the sow for at least 30 min, and 2 ml oxytocin (Oxytocin ‘Intervet’ Vet, 10 IE/ml; Intervet International B. V., Boxmeer, Netherlands) was injected intramuscularly to induce milk letdown. Milk was sampled from four to five teats and stored in 50-ml tubes (Sarstedt, Nümbrecht, Germany) at −20°C until analysis for DM, lactose, fat and true protein (MilcoScan FT2, Foss Electric, Denmark).
On another subsample of 60 sows (10 per treatment), body water content was measured by the deuterium oxide (D2O) dilution technique (Theil et al., Reference Theil, Nielsen, Kristensen, Labouriau, Danielsen, Lauridsen and Jakobsen2002) on days 3 and 24 postpartum. An initial blood sample was drawn from the jugular vein into a 4-ml lithium-heparinized tube (BD Vacutainer, Plymouth, UK) just before D2O injection, and then 5 h after injection with D2O, a second sample was taken. A dose of 0.2 ml (10% solution of D2O; Sigma Aldrich, Brøndby, Denmark) per kg BW was injected i.m. in the neck of the sow. The exact amount of D2O injected was determined by weighing the syringe before and after injection. The blood samples were centrifuged for 10 min (1560×g) at room temperature, and plasma was then recovered and stored in 1.5-ml microcentrifuge tubes (Kruuse, Langeskov, Denmark) at −20°C until analysis. The total D2O space was determined as described by Theil et al. (Reference Theil, Nielsen, Kristensen, Labouriau, Danielsen, Lauridsen and Jakobsen2002).
Calculations and statistical analysis
Calculations and statistical analyses were carried out using the statistical software R (R Core Team, Vienna, Austria) with the individual sow regarded as the experimental unit. Average daily gain (ADG) of the litter was calculated from litter size and weight at standardization and at weaning. Weights of dead piglets or piglets removed from the litter during the experiment were included in the litter gain. Milk yield was estimated from ADG of the litter and litter size (data not shown) using equations by Hansen et al. (Reference Hansen, Strathe, Kebreab, France and Theil2012). The content of body water was estimated based on the measured concentrations of D2O in plasma. From body water, BF and BW, the body protein, fat and ash contents were estimated using equations by Rozeboom et al. (Reference Rozeboom, Pettigrew, Moser, Cornelius and el Kandelgy1994). In the analysis of the dose–response data, the dietary SID CP concentrations were used as an explanatory variable. Based on the recordings of individual daily feed intakes of the sows, chemical analysis of dietary composition, and digestibility coefficients of CP of the six diets, an average dietary SID CP concentration was calculated for the individual sow.
Data were fitted using a mixed-effects model in ANOVA form using the nlme package in R to describe the effect of the six dietary treatments. Feed intake, sow BW and BF, sow body composition and changes in body composition, and litter weight were analysed using model (1). The interaction between diet and parity was tested, but the interaction and the effect of parity were not significant and therefore eliminated in the final model:
 $${Y_{ijk}} = \mu + {\alpha _i} + {\beta _j} + {\varepsilon _{ijk}}$$
(1)
$${Y_{ijk}} = \mu + {\alpha _i} + {\beta _j} + {\varepsilon _{ijk}}$$
(1)
where Y
ijk
is the response variable measured on the k th sow (k = 1, …, n
ij
) in the jth block (j = 1, 2, …, 16) receiving the i th diet (i = 1, …, 6). Here µ is the overall mean, α
i
is the fixed effect of diet, β
j
is the random effect of block that was assumed to be N(0,
 $\sigma _B^2$
), and ε
ijk
is the random error component, which was assumed to be N(0,
$\sigma _B^2$
), and ε
ijk
is the random error component, which was assumed to be N(0,
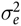 $\sigma _e^2$
).
$\sigma _e^2$
).
When analysing milk yield and composition, daily secretions and concentrations of plasma metabolites, a repeated-measures model was fitted to data. The three-way interaction between diet, parity and day were included in model (2), but none of the three-way or two-way interactions and the effect of parity were significant and therefore excluded from the final model:
 $${Y_{ijkl}} = {\rm{ }}\mu {\rm{ }} + {\alpha _i} + {\beta _j} + {\gamma _k} + {\nu _l} + {\varepsilon _{ijkl}}$$
(2)
$${Y_{ijkl}} = {\rm{ }}\mu {\rm{ }} + {\alpha _i} + {\beta _j} + {\gamma _k} + {\nu _l} + {\varepsilon _{ijkl}}$$
(2)
where Y
ijkl
is the response variable measured on the lth sow (l = 1, …, n
ijk
) in the kth block (k = 1, 2, …, 16) receiving the ith diet (i = 1, …, 6) on the jth day postpartum (j = 1, …, 5). Here µ is the overall mean, α
i
is the fixed effect of diet, β
j
is the fixed effect of day postpartum, γ
k
is the random effect of block that was assumed to be N(0,
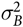 $\sigma _B^2$
), ν
l
is the random effect of sow that was assumed to be N(0,
$\sigma _B^2$
), ν
l
is the random effect of sow that was assumed to be N(0,
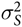 $\sigma _S^2$
), and ε
ijkl
is the random error component that was assumed to be N(0,
$\sigma _S^2$
), and ε
ijkl
is the random error component that was assumed to be N(0,
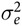 $\sigma _e^2$
). Plasma concentrations of lactate, NEFA, alkaline phosphatase and GGT were log-transformed, because of lacking variance homogeneity. Results are given as least squares means and SE. Log-transformed data were back-transformed and given as least squares means and 95% confidence interval (CI). Statistical significance was declared at P < 0.05 and tendency at P < 0.10.
$\sigma _e^2$
). Plasma concentrations of lactate, NEFA, alkaline phosphatase and GGT were log-transformed, because of lacking variance homogeneity. Results are given as least squares means and SE. Log-transformed data were back-transformed and given as least squares means and 95% confidence interval (CI). Statistical significance was declared at P < 0.05 and tendency at P < 0.10.
All response variables were analysed using either a linear dose–response function or nonlinear dose–response function, being the linear broken-line and quadratic broken-line. Here dietary CP concentration was treated as a continuous variable. The nonlinear dose–response models were fitted using nonlinear mixed-effects models, where the random effect of block was incorporated into the plateau parameter of the models as described by Robbins et al. (Reference Robbins, Saxton and Southern2006). Taking the linear plateau model as an example, then the nonlinear mixed-effect model was formulated as:
 $${y_{ij}} = \left\{ {\matrix{ {k*\left( {R - C{P_{ij}}} \right);C{P_{ij}} < R} \cr {{y_{{\rm{max}}}} + {b_i};C{P_{ij}} > R} \cr } } \right. + {e_{ij}}$$
(3)
$${y_{ij}} = \left\{ {\matrix{ {k*\left( {R - C{P_{ij}}} \right);C{P_{ij}} < R} \cr {{y_{{\rm{max}}}} + {b_i};C{P_{ij}} > R} \cr } } \right. + {e_{ij}}$$
(3)
where y
ij
denotes response variable for the jth sow in the ith block. Population parameters are the slope k in response to dietary CP
ij
below the requirement R and y
max is plateau level, where the dietary CP
ij
is not limiting sow performance. The random effect b
i
captures the block-to-block variation in sow performance and is assumed to be N(0,
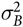 $\sigma _B^2$
), and ε
ij
is the random error component, which was assumed to be N(0,
$\sigma _B^2$
), and ε
ij
is the random error component, which was assumed to be N(0,
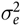 $\sigma _e^2$
)
$\sigma _e^2$
)
The model output was evaluated using three criteria: (1) comparison of Akaike Information Criteria and −2 log likelihood fit statistics for nested models, (2) models with breakpoint outside the range of the tested SID CP concentrations was regarded inappropriate, and (3) evaluation of visual fit to data. The nonlinear models were compared to the linear model using the abovementioned criteria. However, only results of variables with a significant effect of dietary CP concentration are shown in the Results section. All data were used to fit the models, but in Figures 1 and 2 the least square means for the six groups are also presented together with the best model as described above.

Figure 1 Changes in the contents of body water, protein and ash from day 3 to day 24 postpartum with increased standard ilea digestible (SID) dietary protein. The content of body water was determined using the deuterium dilution technique (Theil et al., Reference Theil, Nielsen, Kristensen, Labouriau, Danielsen, Lauridsen and Jakobsen2002). Body protein and ash were estimated from body water, BW and back fat thickness. The loss is given as the percentage of tissue mass at day 3. Data were fitted using linear regression, linear broken-line and quadratic broken-line models using nonlinear mixed-effects models. The model with the best fit is presented in the figure. Data points in the figures are least square means and SE of the six dietary groups, and the linear broken-line model or the linear regression model using data of individual sows are also presented. (a) Body water loss decreased linearly (−0.3 × (SID CP – 130) × (SID
 ${\rm{CP}} \le 130$
) – 2.5) until 130 g SID CP/kg at a loss of 2.5% (P<0.001), (b) body protein decreased linearly (−0.2 × (SID CP – 128) × (SID
${\rm{CP}} \le 130$
) – 2.5) until 130 g SID CP/kg at a loss of 2.5% (P<0.001), (b) body protein decreased linearly (−0.2 × (SID CP – 128) × (SID
 ${\rm{CP}} \le 128$
) + 1.5) until 128 g SID CP/kg at a loss of 1.5% (P<0.001) and (c) body ash loss decreased linearly (Y = 25.5−0.23 × SID CP (g/kg); P<0.001). Negative values indicate a gain.
${\rm{CP}} \le 128$
) + 1.5) until 128 g SID CP/kg at a loss of 1.5% (P<0.001) and (c) body ash loss decreased linearly (Y = 25.5−0.23 × SID CP (g/kg); P<0.001). Negative values indicate a gain.

Figure 2 Effect of dietary standardized ileal digestible (SID) CP concentrations on the plasma concentrations of metabolites. Data were fitted using linear regression, linear broken-line and quadratic broken-line models using nonlinear mixed-effects models. The model with the best fit is presented in the figure. Data points in the figures are least square means and SE of the six dietary groups, and the linear broken-line model or the linear regression model using data of individual sows are also presented. (a) Urea N was at a stable level until a linear increase from 139 g SID CP/kg (0.09 × (SID CP – 139) × (SID CP ≥ 139) + 3.8) in early lactation (day 3 + day 10) and at a stable level until a linear increase from 133 g SID CP/kg (0.09 × (SID CP – 133) × (SID CP ≥ 133) + 4.5) in late (day 17 + day 24) lactation, respectively (P<0.001); (b) albumin increased linearly (Y = 27.7 + 0.06 × SID CP (kg/day)) at late (day 17 + day 24) lactation (P<0.05); (c) alanine amino transferase increased linearly at day 17 (Y = 72.8 – 0.23 × SID CP (g/kg); P<0.01) and day 24 (Y = 80.1 – 0.26 × SID CP (g/kg); P<0.05) of lactation and (d) gamma-glutamyl transferase (days 3 + 10 + 17 + 24) increased linearly (Y = 31.2 + 0.05 × SID CP (g/kg)) with increasing dietary SID CP (P<0.05), respectively.
When the ANOVA revealed that there was no difference between sampling days (P>0.10), data for these days were pooled. For PUN, data from days 3 and 10, and days 17 and 24, respectively, were combined. Data for plasma albumin at days 17 and 24 were united. For GGT, data from days 3, 10, 17 and 24 were pooled.
Results
Results for the model with the best fit to data according to the criteria given above are presented. The nonlinear models were compared to the linear regression model, and for the change in body water and protein, concentrations of milk lactose and CP and PUN, the linear broken-line model was chosen over other models. For plasma concentrations of albumin, GGT and ALT, change in body ash and concentration of milk fat, the linear regression model was the best fit to data.
Diets and sow performance
The concentration of protein was slightly higher in the six diets than planned, because wheat and barley had a higher content of crude protein than expected in the feed formulation. The average daily feed intake of the sows was similar in all groups (Table 2; P=0.78), and as planned, the daily intake of protein increased with increasing SID CP concentrations of the six diets (P < 0.001).
Table 2 Effect of increased dietary standardized ileal digestible (SID) CP on sow and piglet performance (n = 92) 1

1 The interaction of Diet × Parity was not significant (P > 0.05).
Litter size (12.8 ± 1.2 piglets) at weaning was unaffected by dietary SID CP concentration, whereas ADG of the litters (0.01 × (SID CP − 135) × (SID
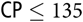 ${\rm{CP}} \le 135$
) + 2.53first parity or 3.07multiparity) and BF loss (−0.03 × (SID CP − 127) × (SID
${\rm{CP}} \le 135$
) + 2.53first parity or 3.07multiparity) and BF loss (−0.03 × (SID CP − 127) × (SID
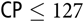 ${\rm{CP}} \le 127$
) − 2.97) of sows increased and BW loss (0.007 × (SID CP − 143) × (SID
${\rm{CP}} \le 127$
) − 2.97) of sows increased and BW loss (0.007 × (SID CP − 143) × (SID
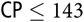 ${\rm{CP}} \le 143$
) − 0.58) decreased with increasing dietary SID CP (P < 0.001; data not shown). For a detailed description of these results, see Strathe et al. (Reference Strathe, Bruun, Geertsen, Zerrahn and Hansen2017a).
${\rm{CP}} \le 143$
) − 0.58) decreased with increasing dietary SID CP (P < 0.001; data not shown). For a detailed description of these results, see Strathe et al. (Reference Strathe, Bruun, Geertsen, Zerrahn and Hansen2017a).
Milk composition and nutrient output
Milk composition was only affected by dietary SID CP at day 17 of lactation. Milk fat and DM contents decreased throughout lactation (P < 0.001), and at day 17, milk fat concentration increased linearly with increasing dietary SID CP (P < 0.05), as an increase by 1 g SID CP/kg increased the fat content with 0.02 percentage units. Milk lactose concentration on day 17 of lactation decreased with increasing dietary SID CP until a breakpoint at 124 g SID CP/kg (5.3%; P < 0.001), but the lactose concentration of the milk did not change throughout lactation (P=0.15). Milk protein concentration decreased from day 3 to day 10 of lactation (P < 0.001). The concentration of milk protein on day 17 increased until a breakpoint at 136 g SID CP/kg (5.0%; P < 0.001). Results of milk yield and composition are given in Table 3.
Table 3 Effect of increased dietary standardized ileal digestible (SID) CP on milk yield, composition and nutrient output of lactating sows 1

1 The interactions of Diet × Day and Diet × Parity and the effect of Parity were not significant (P>0.05).
2 Estimated from equation by Hansen et al. (Reference Hansen, Strathe, Kebreab, France and Theil2012).
Body composition of sows
Results of body composition measurements are given in Table 4 and Figure 1. The loss of body water from day 3 of lactation to weaning decreased with increasing dietary SID CP until a dietary SID CP of 130 g/kg (P < 0.001), and a similar pattern was seen for the loss of protein, which reached a breakpoint at 128 g SID CP/kg (P < 0.001). The body ash loss declined linearly with increasing dietary SID CP (P < 0.01), whereas the change in body fat was unaffected by dietary treatment (P=0.41).
Table 4 Body composition and changes in body composition in the sows measured by deuterium dilution technique 1

SID=standardized ileal digestible.
1 The interaction of Diet × Parity was not significant (P>0.05). For body composition on days 3 and 24, there was an effect of Parity (P<0.05), because of different BW of parity 2 and 3 sows. There was no effect of Parity on changes in body composition (P>0.05).
2 Body water was determined by the technique described by Theil et al. (Reference Theil, Nielsen, Kristensen, Labouriau, Danielsen, Lauridsen and Jakobsen2002).
3 Negative values indicate a gain.
Plasma metabolites
As expected, the concentrations of metabolites in plasma were similar for all groups at day −4 (P > 0.05). The concentrations of PUN increased with increasing dietary SID CP intake (Figure 2; P < 0.001). The concentrations of PUN were similar at days 3 and 10, and at days 17 and 24, respectively, and there was an interaction between diet and stage of lactation (P < 0.05). In early lactation (day 3 + day 10), PUN increased linearly after the breakpoint at 139 g SID CP/kg at a concentration at 3.8 mmol/l, but in late lactation (day 17 + day 24), PUN increased linearly after a breakpoint at 133 g SID CP/kg (P < 0.001, Figure 2) at a concentration of 4.5 mmol/l. Albumin concentration increased linearly in late lactation with increasing dietary SID CP (day 17 + day 24; P < 0.05), and GGT also increased linearly (P<0.05) at all stages of lactation, whereas the concentration of ALT decreased linearly at day 17 (P < 0.01) and day 24 (P < 0.05) with increasing dietary SID CP concentration (Figure 2).
The dietary SID CP did not affect plasma concentrations of glucose, lactate, NEFA, triglycerides, cholesterol, creatinine, total protein, AST and alkaline phosphatase (P>0.05; data not shown), whereas all of the abovementioned metabolites were affected by day (P < 0.001; data not shown) except for lactate (P>0.05).
Discussion
The current study estimated the SID CP requirement of hyper-prolific lactating sows using litter gain as the main determinant supported by measurements on several traits of sow metabolism. Based on the average feed intake and the SID CP concentration of 135 g/kg (Strathe et al., Reference Strathe, Bruun, Geertsen, Zerrahn and Hansen2017a), the daily requirement for optimal litter ADG was estimated to be 850 g SID CP/day. The 850 g SID CP/day found in the current study was considerably higher than the 700 g SID CP/day estimated in the NRC model (National Research Council, 2012) and slightly lower than the 880 g SID CP/day (±70 g/day) estimated in the requirement model by Strathe et al. (Reference Strathe, Strathe, Theil, Hansen and Kebreab2015). The largest difference between the two models is the equation used to determine milk production: the one used by NRC (2012) is based on weigh-suckle-weigh data, which underestimates milk yield and thereby estimates a lower requirement (Hansen et al., Reference Hansen, Strathe, Kebreab, France and Theil2012).
Milk production
The main purpose of the lactating sow is to produce milk with a nutrient composition and yield that matches the requirements of the piglets. Dietary SID CP concentration did only affect milk nutrient concentration and nutrient yield on day 17 of lactation. The improved ADG of the litter with increasing dietary SID CP most likely was a combined result of increased milk yield throughout lactation and an increased concentration of protein at peak lactation. Many studies have proven it difficult to increase the concentration of protein in sow milk above 5.0% to 5.5% (Hurley, Reference Hurley and Farmer2015). The true milk protein content at the breakpoint on day 17 reached only 5.0%, which was not unusually high compared to other studies (Guan et al., Reference Guan, Pettigrew, Ku, Ames, Bequette and Trottier2004; Laspiur et al., Reference Laspiur, Burton, Weber, Moore, Kirkwood and Trottier2009) although it was higher than shown in another recent trial with hyper-prolific sows where the true milk protein content was approximately 4.5% throughout lactation (Pedersen et al., Reference Pedersen, Bruun, Feyera, Larsen and Theil2016). The higher protein intake resulted in concomitant higher plasma concentrations of GGT, which supported the view that increased milk protein concentration was caused by increasing the uptake of amino acids, because this enzyme has been shown to be involved in the mammary gland uptake of amino acids in rats (Viña et al., Reference Viña, Puertes, Estrela, Viña and Galbis1981a and Reference Viña, Puertes and Viña1981b) The piglet has a high rate of muscle growth and therefore a high protein requirement (Theil and Jørgensen, Reference Theil and Jørgensen2016), and the litter gain reached maximum at 135 g SID CP/kg, which is identical to the dietary SID CP concentration which resulted in maximum milk protein content (136 g SID CP/kg).
Changes in body composition
Lactating sows often turn catabolic, and therefore milk nutrients are derived from both diet and body reserves. The decrease in BW loss in the current experiment was mainly a result of decreased body protein mobilization, because BF mobilization continued to increase with increased protein intake. A similar response to increased dietary protein was reported by Dourmad et al. (Reference Dourmad, Noblet and Etienne1998). This suggests that sows fed high levels of dietary SID CP use the dietary protein as substrate for milk protein and thereby save body protein. The lower protein mobilization in sows fed high SID CP levels were followed by decreased plasma ALT concentrations as a result of the downregulation of urea cycle enzymes (Das and Waterlow, Reference Das and Waterlow1974). Feeding protein-deficient diets results in a reduction of the body protein turnover to ensure a higher utilization of dietary protein. A lower plasma albumin concentration observed in sows fed low SID CP concentrations has also been shown in pigs suffering protein deficiency (Wykes et al., Reference Wykes, Fiorotto, Burrin, Del Rosario, Frazer, Pond and Jahoor1996), and this indicated that these sows had a reduced albumin synthesis. In this study, minimum body protein loss (128 SID CP g/kg) occurred at a dietary SID CP concentration close to that where milk protein was maximized (136 g SID CP/kg). In addition, the N balance was optimized at SID CP levels close to these concentrations as indicated by an increase in PUN in both early (139 g SID CP/kg) and late (133 g SID CP/kg) lactation above these concentrations. The increase in PUN concentrations after the breakpoint indicated that protein intake above this dietary protein concentration was in excess of the requirement (Coma et al., Reference Coma, Carrion and Zimmerman1995 and Reference Coma, Zimmerman and Carrion1996). The breakpoint for PUN was reached at higher SID CP concentrations in early compared to late lactation, because of the lower daily CP intake in early lactation. Minimizing body protein mobilization during lactation is important, because it can be difficult for the sow to regain the lost body protein during the following gestation. This is in accordance with Dourmad et al. (Reference Dourmad, Etienne and Noblet1996), who reported that sows losing 35 v. 12 kg during lactation were unable to restore the body reserves, especially body protein, during subsequent gestation.
Body ash is a measure of bone mass, and therefore the lower mobilization of ash with a higher dietary protein intake suggests that bone mobilization was reduced. Low protein intakes reduce intestinal absorption of calcium and increase the concentration of parathyroid hormone, resulting in calcium being released from the bones (Heaney and Layman, Reference Heaney and Layman2008). This can negatively affect the longevity of the sows, because leg disorders are a cause of culling in sow herds (Engblom et al., Reference Engblom, Lundeheim, Dalin and Andersson2007).
In the current study, milk fat content and BF mobilization increased with increasing dietary protein intake, which was in accordance with the results by King et al. (Reference King, Toner, Dove, Atwood and Brown1993). This indicated that the mobilized body fat was redirected towards the mammary glands for milk fat production. Back fat loss increased with increased dietary protein, but changes in body fat estimated by D2O dilution technique was unaffected by the tested dietary protein levels, which is in accordance with Mejia-Guadarrama et al. (Reference Mejia-Guadarrama, Pasquier, Dourmad, Prunier and Quesnel2002). The lacking response of body fat might be ascribed to the used equations (Rozeboom et al., Reference Rozeboom, Pettigrew, Moser, Cornelius and el Kandelgy1994), which were developed on data from gilts. The increased BF mobilization suggests that the sows fed high levels of dietary SID CP most likely were in an energy deficit. Increased energy demand will often result in increased body fat mobilization, when energy intake is not increased at the same rate as the requirement. The six diets were isoenergetic, and this resulted in diets with different ratios between energy and protein. It has been observed in other studies that sows fed high CP levels increase BF mobilization (Tritton et al., Reference Tritton, King, Campbell, Edwards and Hughes1996; McNamara and Pettigrew, Reference McNamara and Pettigrew2002; Heo et al., Reference Heo, Yang, Jin, Park, Ynag and Chae2008). However, it was also shown that both increasing dietary CP and energy level could minimize BF loss (Tritton et al., Reference Tritton, King, Campbell, Edwards and Hughes1996; McNamara and Pettigrew, Reference McNamara and Pettigrew2002), which emphasizes the importance of also determining the energy requirement of sows when increasing protein intake.
Conclusions
The SID CP requirement of hyper-prolific lactating sows was estimated at 135 g/kg based on the ADG of the litter. At this level of SID CP, BW loss, body protein loss and body ash loss were considerably reduced, and N balance of the sows measured by PUN was optimized. The higher litter gain was a result of increased milk yield and higher contents of protein and fat in milk at peak lactation.
Acknowledgements
The project received financial support from the Pig Levy Foundation and from the EU and Rural Development Program under the Danish Ministry of Food, Agriculture and Fisheries, journal number 32101-U-13-00239, and the Faculty of Health and Medical Sciences, University of Copenhagen. Feed analysis was sponsored by Evonik Nutrition & Care GmbH Tyskland Filial Denmark. Results in this article were presented in the PhD thesis of the first author (Strathe, Reference Strathe2017).
Declaration of interest
None.
Ethics statement
The Danish Animal Experimentation Inspectorate (Authorization No. 2013-15-2934-00961) approved the trial.
Software and data repository resources
None of the data were deposited in an official repository.






