



Asthma is a heterogeneous disease that affects approximately 235 million people worldwide( 1 ). It is characterised by intermittent reversible bronchoconstriction, chronic airway inflammation and respiratory symptoms such as wheezing, dyspnoea, chest tightness and cough. The immunopathology of asthma is predominantly orchestrated by T-helper 2 (TH2) cells and their pro-inflammatory cytokines and chemokines, which recruit secondary effector cells including IgE-activated mast cells, macrophages, basophils and eosinophils( Reference Holgate 2 ). Exercise-induced bronchoconstriction (EIB) is a phenotype of asthma, which is characterised by transient airway narrowing during and/or after exercise( Reference Wenzel 3 , Reference Hallstrand, Altemeier and Aitken 4 ). EIB is ascribed to airway drying and changes in airway osmolality, which result in degranulation of inflammatory cells and release of inflammatory mediators( Reference Hallstrand, Altemeier and Aitken 4 ). Inhaled corticosteroids and short- and long-acting β 2-agonists provide effective therapy for asthma, but they are not curative nor do they modify disease progression( Reference Barnes 5 ). Furthermore, long-term inhaled corticosteroid use has undesirable side-effects and adherence is poor, whereas chronic β 2-agonist use results in tolerance( Reference Barnes 5 ). The development of therapies that modulate the immunopathology of asthma without adverse side-effects is therefore desirable.
Gut microbes have a substantial influence on systemic immune function and allergic sensitisation( Reference Roberfroid, Gibson and Hoyles 6 – Reference Hansel, Johnston and Openshaw 8 ); thus, it is possible that manipulation of the gut microbiome may provide a potential strategy to influence the immunopathology of asthma. The observation that allergic asthma patients display lower levels of Bifidobacterium adolescentis supports this concept( Reference Hevia, Milani and López 9 ). Moreover, in humans, 4 weeks of supplementation with Bifidobacterium breve M-16V combined with a prebiotic (non-digestible carbohydrate that enhances the growth and/or activity of beneficial indigenous gut bacteria) improved peak expiratory flow (PEF) and attenuated serum IL-5 after bronchial allergen challenge( Reference Van De Pol, Lutter and Smids 10 ).
Prebiotic galactooligosaccharides are derived from the action of the enzyme β-galactosidase. Bimuno-galactooligosaccharide (B-GOS) is especially potent in selectively increasing the growth and/or activity of bifidobacteria( Reference Depeint, Tzortzis and Vulevic 11 – Reference Vulevic, Juric and Walton 15 ), which has been shown to elicit beneficial immunomodulatory effects in both elderly and overweight adults( Reference Vulevic, Drakoularakou and Yaqoob 12 , Reference Vulevic, Juric and Tzortzis 14 ). In mice exposed to house dust mites, dietary galactooligosaccharides prevented the development of airway hyper-responsiveness and airway eosinophilia and reduced TH2-related cytokine IL-13 and chemokines (chemokine CC ligand 17 (CCL17), chemokine CC ligand 5 (CCL5)) in the lungs( Reference Verheijden, Willemsen and Braber 16 ). The effects of B-GOS on asthma in humans are currently unknown.
In humans, TH2-driven inflammation can be monitored using biomarkers such as the chemokine CC ligand 11 (CCL11) and CCL17, along with IgE, TNF-α, C-reactive protein (CRP) and fraction of exhaled nitric oxide (FENO)( Reference Wenzel 3 , Reference Szefler, Wenzel and Brown 17 ). Eucapnic voluntary hyperpnoea (EVH) causes a highly reproducible hyperpnoea-induced bronchoconstriction (HIB) (a surrogate for EIB) in adults( Reference Williams, Johnson and Hunter 18 ), which makes this an excellent challenge test to evaluate the effects of B-GOS supplementation on airway hyper-responsiveness.
Therefore, the aim of this study was to test the hypothesis that B-GOS supplementation in adults with asthma attenuates the severity of HIB and that this is associated with reduced systemic concentrations of TH2-driven inflammatory markers.
Methods
Participants and study design
A total of ten participants (five males) formed a HIB group, and eight participants (five males) with no history of asthma formed a control group (Fig. 1; Table 1). All participants were non-smokers. Inclusion criteria for the HIB group were physician diagnosis of asthma, a baseline forced expiratory volume in 1 s (FEV1)>65 % of the predicted( Reference Williams, Johnson and Hunter 18 ) and a ≥10 % fall in FEV1 following an initial EVH screening test( Reference Parsons, Hallstrand and Mastronarde 19 ). Participants in the HIB group were on steps 1–3 of the global initiative for asthma stepwise approach to asthma control( Reference Boulet, FitzGerald and Reddel 20 ).

Fig. 1 Participant flow diagram. HIB, hyperpnoea-induced bronchoconstriction; B-GOS, Bimuno-galactooligosaccharides.
Table 1 Anthropometric data, baseline pulmonary function and medication (Numbers and percentages of predicted; mean values and standard deviations)

HIB, hyperpnoea-induced bronchoconstriction; FVC, forced vital capacity; FEV1, forced expiratory volume in 1 s; PEF, peak expiratory flow; M, male; S, salbutamol; BUD, budesonide; /, in combination with; FORM, formoterol; F, female; Sm, salmeterol; BEC, beclomethasone.
This study was conducted according to the guidelines laid down in the Declaration of Helsinki, and all procedures were approved by the Nottingham Trent University Human Ethics Committee (Approval no. 229; Clinical Trial no. ISRCTN15022880). All participants provided their written informed consent. The present study adopted a randomised (block randomisation), double-blind, placebo-controlled, cross-over design over 8 consecutive weeks. Participants were randomly assigned to receive 5·5 g/d of either B-GOS or placebo (maltodextrin) (Clasado Ltd) for 3 weeks. Thereafter, participants followed a 2-week washout period( Reference Depeint, Tzortzis and Vulevic 11 ) (normal diet) before commencing the alternative supplement for the remaining 3 weeks (Fig. 1). The supplements were identical in taste and colour, and double blinding was completed by Clasado Ltd. The B-GOS dose used in the present study has been shown to consistently increase the number of bifidobacteria within the gut( Reference Depeint, Tzortzis and Vulevic 11 – Reference Vulevic, Juric and Walton 15 ).
At day 0 and day 21 of each treatment, baseline FENO was measured and an EVH test was performed at Nottingham Trent University. Before and after EVH, pulmonary function was assessed, and venous blood samples were collected for analysis of inflammatory markers. HIB participants were permitted to use their medication as required but stopped taking it before each EVH test (see below).
Fraction of exhaled nitric oxide, pulmonary function and eucapnic voluntary hyperpnoea test
Baseline FENO was measured (NIOX MINO; Aerocrine) according to American Thoracic Society/European Respiratory Society (ATS/ERS) guidelines(
Reference Dweik, Boggs and Erzurum
21
) in the HIB group only, as it is elevated in asthma patients but not in healthy controls(
Reference Alving, Weitzberg and Lundberg
22
). In accordance with ATS/ERS guidelines(
Reference Miller, Hankinson and Brusasco
23
), pulmonary function (forced vital capacity (FVC), PEF and FEV1) was assessed in triplicate at baseline and in duplicate at 3, 6, 10, 20 and 30 min after EVH. The highest values recorded were used for analyses. Whole blood (20 ml) was collected at baseline and 15 min, 60 min and 24 h after EVH. The EVH test comprised 6 min of EVH using a dry gas mixture at a target minute ventilation
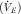 ${\rm (}\dot{V}_{E} )$
of 85 % of the predicted maximum voluntary ventilation (30×baseline FEV1)(
Reference Williams, Johnson and Hunter
18
,
Reference Anderson, Argyros and Magnussen
24
). Participants avoided exercise for 24 h before each EVH test, and participants in the HIB group stopped taking their medication (inhaled corticosteroids: 4 d; inhaled long-acting β
2 agonists: 2 d; inhaled short-acting β
2 agonists: the day of the test)(
Reference Anderson, Argyros and Magnussen
24
). On EVH test days, participants abstained from caffeine and alcohol and arrived at the laboratory >2 h postprandially(
Reference Williams, Johnson and Hunter
18
,
Reference Anderson, Argyros and Magnussen
24
).
${\rm (}\dot{V}_{E} )$
of 85 % of the predicted maximum voluntary ventilation (30×baseline FEV1)(
Reference Williams, Johnson and Hunter
18
,
Reference Anderson, Argyros and Magnussen
24
). Participants avoided exercise for 24 h before each EVH test, and participants in the HIB group stopped taking their medication (inhaled corticosteroids: 4 d; inhaled long-acting β
2 agonists: 2 d; inhaled short-acting β
2 agonists: the day of the test)(
Reference Anderson, Argyros and Magnussen
24
). On EVH test days, participants abstained from caffeine and alcohol and arrived at the laboratory >2 h postprandially(
Reference Williams, Johnson and Hunter
18
,
Reference Anderson, Argyros and Magnussen
24
).
Analysis of inflammatory markers
Concentrations of serum chemokines CCL11 and CCL17 were determined using multiplex analysis (Bio-Plex 200; Bio-Rad Laboratories Limited) and Luminex screening assay plates (R&D Systems) as previously described( Reference Sutavani, Bradley and Ramage 25 ). ELISA was used to determine concentrations of TNF-α (R&D Systems), CRP and IgE (Universal Biologicals). The TNF-α ELISA does not cross-react with human IL-1β, IL-1α, IL-2-13 or TNF-β and is specific for the measurement of natural and recombinant human TNF-α. For TNF-α, the intra- and inter-assay variation was <10 % and the minimum detectable level of the assay was 0·60 pg/ml, which all samples exceeded. For CRP and IgE, the minimum detectable levels of the assays were 0·25 ng/ml and 0·29 KU/l, respectively, which all samples exceeded.
Statistics
The average minimum perceptible improvement in FEV1 in adults with asthma is 230 ml( Reference Santanello, Zhang and Seidenberg 26 ), whereas the within-participant standard deviation for the fall in FEV1 after EVH is 100 ml( Reference Williams, Johnson and Hunter 18 ). An a priori sample size calculation revealed that with power=0·90 and α=0·05, a sample size of seven would be required to detect a 230-ml change in the fall in FEV1 after EVH.
Pulmonary function data and serum TNF-α concentrations were analysed using repeated-measures ANOVA and Bonferroni-adjusted paired t tests. CCL11, CCL17, CRP and IgE were analysed using non-parametric Friedman’s repeated measures and Wilcoxon’s signed-rank tests. Between-group differences in IgE were analysed using the Kruskal–Wallis test. The AUC of the percentage fall in FEV1 during the 30-min period after EVH (%ΔFEV1AUC0–30) was calculated using trapezoidal integration. Statistical significance was set at P<0·05. Data are presented as mean values and standard deviations unless otherwise indicated.
Results
Pulmonary function
In the control and HIB groups, there were no between-day differences in baseline FEV1. Pooled baseline FEV1 tended to be lower in the HIB group compared with the control group (P=0·066) (Table 1).
For the peak fall in FEV1 after EVH, three-way, repeated-measures ANOVA revealed a treatment×day×group interaction (P<0·001). In the control group, the peak fall in FEV1 after EVH was unchanged after placebo and B-GOS (pooled data: −3 (sd 3) %). In the HIB group, the peak fall in FEV1 after EVH was unchanged from day 0 (−880 (sd 480) ml) to day 21 (−840 (sd 430) ml) of placebo. Conversely, following B-GOS, the peak fall in FEV1 after EVH was attenuated by 40 % from day 0 (−940 (sd 460) ml) to day 21 (−570 (sd 310) ml) (mean difference=370 (sd 290) ml; 95 % CI 166, 575 ml, P=0·004) (Fig. 2 and 3). In the control group, the overall severity of HIB, as determined by %ΔFEV1AUC0–30, was unchanged after placebo and B-GOS (pooled data: −48 (sd 53) ml). In the HIB group, %ΔFEV1AUC0–30 was unchanged from day 0 (−530 (sd 384)) to day 21 (−523 (sd 366)) of placebo, whereas a 41 % reduction was observed from day 0 (−583 (sd 404)) to day 21 (−345 (sd 267)) of B-GOS (mean difference=−237 (sd 263); 95 % CI −425, −48, P=0·019).

Fig. 2 Peak fall in forced expiratory volume in 1 s (FEV1) after eucapnic voluntary hyperpnoea. Values are means and standard deviations represented by vertical bars. * Bimuno-galactooligosaccharides (B-GOS) day 0 v. B-GOS day 21 (P=0·004). ![]() , Placebo day 0;
, Placebo day 0; ![]() , placebo day 21;
, placebo day 21; ![]() , B-GOS day 0;
, B-GOS day 0; ![]() , B-GOS day 21.
, B-GOS day 21.

Fig. 3 The individual data and mean values and standard deviations for the peak fall in forced expiratory volume in 1 s (FEV1) after eucapnic voluntary hyperpnoea in hyperpnoea-induced bronchoconstriction participants only. Individual participants are represented by the same symbols in both the placebo and B-GOS figures. B-GOS, Bimuno-galactooligosaccharides.
In the control and HIB groups, there were no between-day differences in baseline FVC and PEF. Baseline FVC and PEF were not different between control and HIB groups (Table 1). The peak fall in FVC after EVH was greater in the HIB group (−18 (sd 16) %; −78 (sd 54) ml) than in the control group (−3 (sd 2) %; −14 (sd 12) ml) (P=0·005), but was unchanged in both groups after placebo and B-GOS. The peak fall in PEF after EVH was greater in the HIB group (−27 (sd 12) %) than in the control group (−9 (sd 6) %) (P=0·007) and was unchanged after placebo in both groups and after B-GOS in the control group. Conversely, after B-GOS in the HIB group, the peak fall in PEF after EVH was reduced from day 0 (−28 (sd 14) %) to day 21 (−17 (sd 10) %) (mean difference=11 (sd 13) %; 95 % CI 1·8, 20·9 %, P=0·024).
Serum TNF-α
At day 0, baseline TNF-α was higher in the HIB group (2·64 (sd 0·81) pg/ml) than in the control group (1·37 (sd 0·37) pg/ml) (P=0·001). A four-way, repeated-measures ANOVA revealed an intervention×day×time×group interaction (P=0·036). Subsequent within-group, three-way, repeated-measures ANOVA revealed an intervention×day×time interaction in the HIB group only (P=0·042). In the HIB group, TNF-α increased by 29 % after EVH at day 0, and this response was unchanged after placebo. Conversely, after B-GOS, baseline TNF-α was reduced (mean difference=0·50 (sd 0·61) pg/ml; 95 % CI 0·02, 0·96 pg/ml, P=0·04) and the 29 % increase in TNF-α after EVH was completely abolished (P=0·002) (Fig. 4).

Fig. 4 TNF-α concentration at baseline and after eucapnic voluntary hyperpnoea (EVH) in hyperpnoea-induced bronchoconstriction (a) and control (b) groups. Values are means and standard deviations. Post-EVH values were averaged for statistical analysis. Significant difference: B-GOS day 0 v. B-GOS day 21 (* P=0·04; ** P=0·002). ●, Placebo day 0; ○, placebo day 21; ▲, Bimuno-galactooligosaccharides (B-GOS) day 0; Δ, B-GOS day 21.
Serum chemokines, C-reactive protein and IgE
In control and HIB groups, serum CCL11, CCL17, CRP and IgE were unchanged after every EVH test; therefore, subsequent analyses were performed on baseline data only. In the control group, baseline CCL11 and CCL17 were unchanged after both interventions. In the HIB group, baseline CCL11 was unchanged after placebo and B-GOS, and CCL17 was unchanged after placebo. Conversely, a priori Wilcoxon’s signed-rank tests revealed a reduction in baseline CCL17 after B-GOS (P=0·005; effect size=−0·88) (Fig. 5).

Fig. 5 Baseline concentrations of the chemokine CC ligand 17 (CCL17) and chemokine CC ligand 11 (CCL11) and C-reactive protein (CRP). Values are means and standard deviations represented by vertical bars. Significant difference: Bimuno-galactooligosaccharides (B-GOS) day 0 v. B-GOS day 21 (* P=0·05; ** P=0·015; *** P=0·005). ![]() , Placebo day 0;
, Placebo day 0; ![]() , placebo day 21;
, placebo day 21; ![]() , B-GOS day 0;
, B-GOS day 0; ![]() , B-GOS day 21.
, B-GOS day 21.
In control and HIB groups, baseline CRP was unchanged after placebo, whereas a priori Wilcoxon’s signed-rank tests revealed a reduction in CRP in control (P=0·050; effect size=−0·49) and HIB (P=0·015; effect size=−0·57) groups after B-GOS (Fig. 5). Baseline IgE was higher in the HIB group (pooled data: 37 (sd 17) KU/l) than in the control group (pooled data: 14 (sd 8) KU/l) (P=0·006) and no changes were observed after placebo or B-GOS.
Fraction of exhaled nitric oxide
In the HIB group, baseline FENO (pooled data: 45 (sd 30) ppb) was unchanged after placebo and B-GOS.
Discussion
The main finding of this study was that supplementation with B-GOS in adults with asthma attenuated the fall in pulmonary function after EVH (reduced peak falls in FEV1 and PEF) and the overall severity of bronchoconstriction (%ΔFEV1AUC0–30). Furthermore, B-GOS reduced baseline concentrations of CCL17, CRP and TNF-α and abolished the EVH-induced increase in TNF-α. These findings suggest that B-GOS can potentially mediate the underlying immunopathology of asthma, and thereby attenuate the airway hyper-responsiveness associated with HIB/EIB.
The 40 % (370 ml) improvement in the post-EVH fall in FEV1 after B-GOS supplementation exceeds the minimum perceptible change of 230 ml( Reference Santanello, Zhang and Seidenberg 26 ), and is therefore clinically relevant. Prophylactic use of β 2-agonists for EIB prevention/protection is common because of their efficacy. For example, salbutamol (200 µg) and salmeterol (50 µg) delivered before exercise reduced the fall in FEV1 by approximately 78 and 61 %, respectively( Reference Anderson, Lambert and Brannan 27 , Reference Anderson, Rodwell and Du Toit 28 ). Comparatively β 2-agonists therefore offer greater protection than B-GOS; however, they are not curative, and some individuals with EIB either do not respond to β 2-agonists or experience a reduction in severity but not symptoms( Reference Anderson, Caillaud and Brannan 29 ). Furthermore, chronic use of β 2-agonists causes tolerance or β 2-receptor desensitisation with associated symptom exacerbation( Reference Anderson, Caillaud and Brannan 29 ). B-GOS is well tolerated( Reference Vulevic, Drakoularakou and Yaqoob 12 , Reference Vulevic, Juric and Tzortzis 14 ), and the mechanisms of action may modify the underlying immunopathological features of asthma( Reference Verheijden, Willemsen and Braber 16 , Reference Sagar, Vos and Morgan 30 ).
The mechanisms by which prebiotics ameliorate airway hyper-responsiveness remain unclear. Previous studies report that B-GOS robustly supports the growth of bifidobacteria in the human gut( Reference Depeint, Tzortzis and Vulevic 11 – Reference Vulevic, Juric and Walton 15 ) and these microbes may interact with the intestinal mucosal immune system to enhance immunomodulatory effects. Dendritic cell sampling of bifidobacteria and lactobacilli may alter naïve T-cell differentiation by promoting an increase in regulatory T-cells expressing Forkhead box protein P3 (Foxp3) and, subsequently, increased production of anti-inflammatory cytokines IL-10 and TGF-β ( Reference McLoughlin and Mills 31 ). Suppression of effector T-cell formation may partially explain the observed changes in TNF-α, which is released by mast cells, neutrophils, eosinophils and airway epithelial cells and causes airway hyper-responsiveness. The HIB group had a higher baseline TNF-α concentration than the control group, which concurs with previous reports of raised TNF-α in athletes with EIB( Reference Mickleborough, Murray and Ionescu 32 ). In the HIB group, B-GOS supplementation reduced baseline TNF-α and, remarkably, abolished the EVH-induced increase in TNF-α. Interestingly, reduced baseline TNF-α concentration after B-GOS supplementation was previously observed in the elderly( Reference Vulevic, Drakoularakou and Yaqoob 12 ). Consequently, the reduced severity of HIB observed after B-GOS supplementation may be explained by modulation of the immune system by the gut microbiota, which results in an attenuated inflammatory response to increased osmolarity of the airway surface liquid during/following EVH.
Allergic asthma and EIB are characterised by increased TH2 cell infiltration, which is partly controlled by the release of CCL11, CCL17 and CCL5 from airway epithelium and bronchial smooth muscle cells( Reference Ying, O’Connor and Ratoff 33 , Reference Holgate 34 ). Baseline serum CCL17 concentration was similar to that reported previously in individuals with allergic asthma( Reference Machura, Rusek‐Zychma and Jachimowicz 35 ) and tended to be higher in the HIB group than in the control group. The reduction in CCL17 following B-GOS supplementation in the HIB group further indicates reduced systemic chemokine expression, which may lower infiltration of TH2 lymphocytes into the airways.
Serum CRP is associated with airway inflammation, obstruction and bronchial hyper-responsiveness in individuals with asthma( Reference Takemura, Matsumoto and Niimi 36 ). Consistent with previous reports in the elderly( Reference Vulevic, Drakoularakou and Yaqoob 12 ), we observed a reduction in CRP in both HIB and control groups after B-GOS. Conversely, FENO and serum IgE were unchanged after B-GOS. FENO is a marker of eosinophilic airway inflammation that is raised in individuals with asthma( Reference Szefler, Wenzel and Brown 17 ). However, because of the heterogeneity of the FENO and IgE measures in the current cohort, a detectable reduction may require a longer duration of B-GOS supplementation.
Faecal samples were not collected in the present study to verify that B-GOS increased the numbers of bifidobacteria. However, we are confident that this was the case, as the prebiotic index of B-GOS, which measures the increase in the number of beneficial bacteria (Bifidobacterium and Lactobacillus–Enterococcus) compared with the reductions in less-favourable bacteria (Bacteroides–Prevotella and Clostridium perfringens–histolyticum), was 0·40 (sd 0·13)( Reference Depeint, Tzortzis and Vulevic 11 ), and previous studies have consistently reported an increase in the number of bifidobacteria within the gut after B-GOS supplementation in humans( Reference Depeint, Tzortzis and Vulevic 11 – Reference Vulevic, Juric and Walton 15 ).
In conclusion, B-GOS supplementation reduced the severity of HIB, and this was associated with reduced systemic concentrations of TH2-driven inflammatory markers. These findings suggest that B-GOS, through its impact on the gut microbiota, has the potential to modulate the underlying immunopathology of asthma, and thereby attenuate the airway hyper-responsiveness associated with HIB/EIB. The precise mechanisms by which B-GOS modulates immune function and reduces airway inflammation remain unclear and warrant further exploration.
Acknowledgements
In-kind support of the B-GOS supplement was provided by Clasado Research Services, but the funder was not involved in the design of the study.
N. C. W., M. A. J., D. E. S., G. R. S. and K. A. H. designed the study; N. C. W., M. A. J., G. R. S. and K. A. H. conducted the study; N. C. W., J. V. and I. S. provided essential reagents and conducted analysis of blood samples; N. C. W., M. A. J. and G. R. S. analysed the data; N. C. W., M. A. J., G. R. S. and K. A. H. wrote the paper; N. C. W., M. A. J., D. E. S., I. S., J. V., G. R. S. and K. A. H. contributed to reviewing and approval of the final manuscript.
J. V. is employed by Clasado Research Services who supplied the supplement but were not involved in the design of the study. None of the other authors had personal or financial conflicts of interest.






